miR-142-3p Reduces the Size, Migration, and Contractility of Endometrial and Endometriotic Stromal Cells by Targeting Integrin- and Rho GTPase-Related Pathways That Regulate Cytoskeletal Function




Abstract
:1. Introduction
2. Methods
2.1. Cell Culture
2.2. microRNA Transfection
2.3. qRT-PCR
2.4. DUAL-Luciferase Reporter Assay
2.5. Western Blot
2.6. Cell Spread Area Analysis
2.7. Migration Assay
2.8. Collagen Gel Contraction Assay
2.9. Immunofluorescence Staining
2.10. Imaging
2.11. Bioinformatics
2.12. Statistical Analysis
3. Results
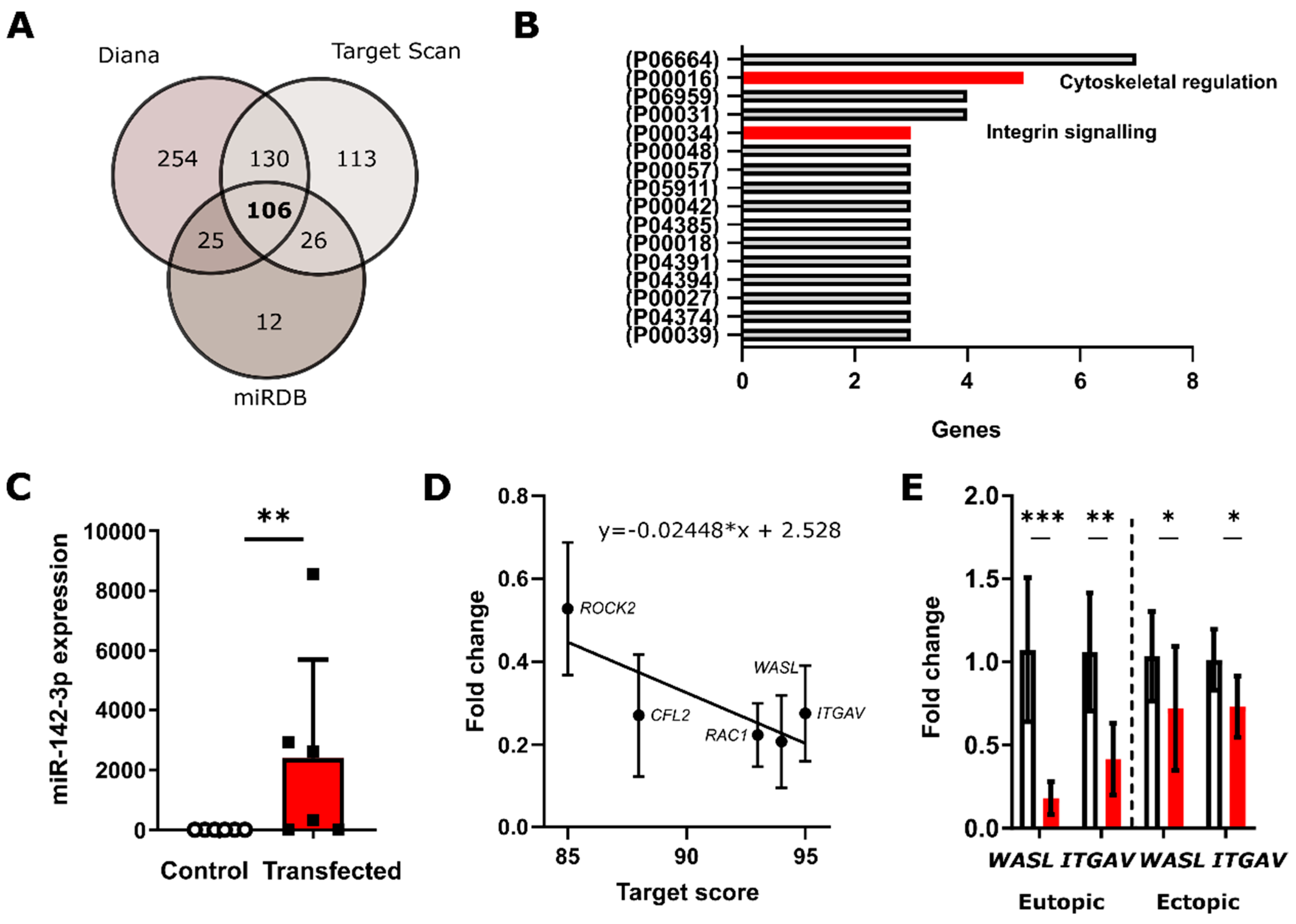
3.1. In Silico Analysis Predicts miRNA-142-3p Targets Several Rho GTPases and Integrin Signaling Pathway Constituents
3.2. qRT-PCR Validates miR-142-3p Regulates ROCK2, CFL2, RAC1 and WASL Gene Expression Levels
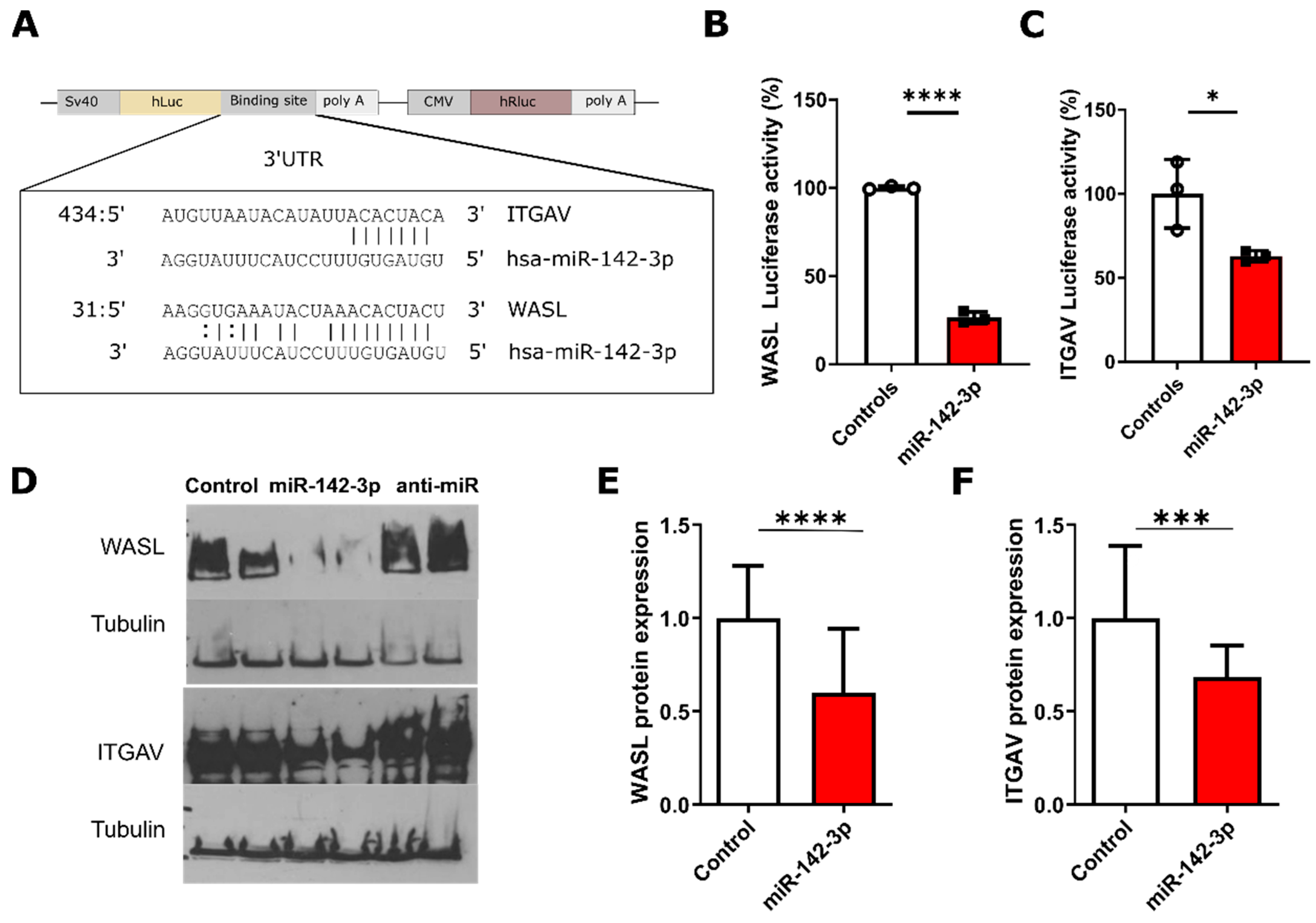
3.3. Luciferase Validation of The Predicted Targets WASL and ITGAV
3.4. Protein-Level Validation of WASL and ITGAV
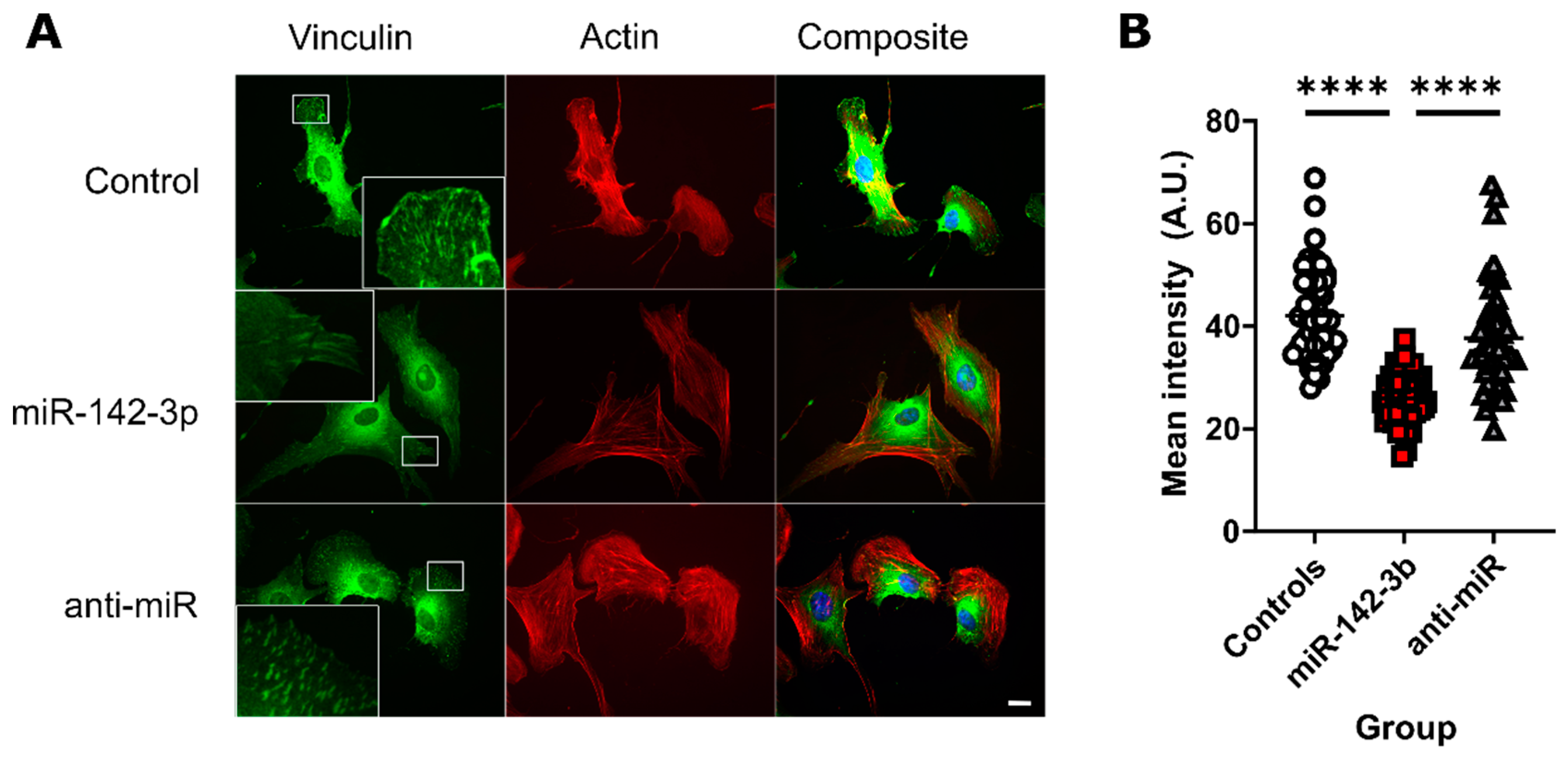
3.5. miR-142-3p Lowers the Concentration of Vinculin in Focal Adhesions
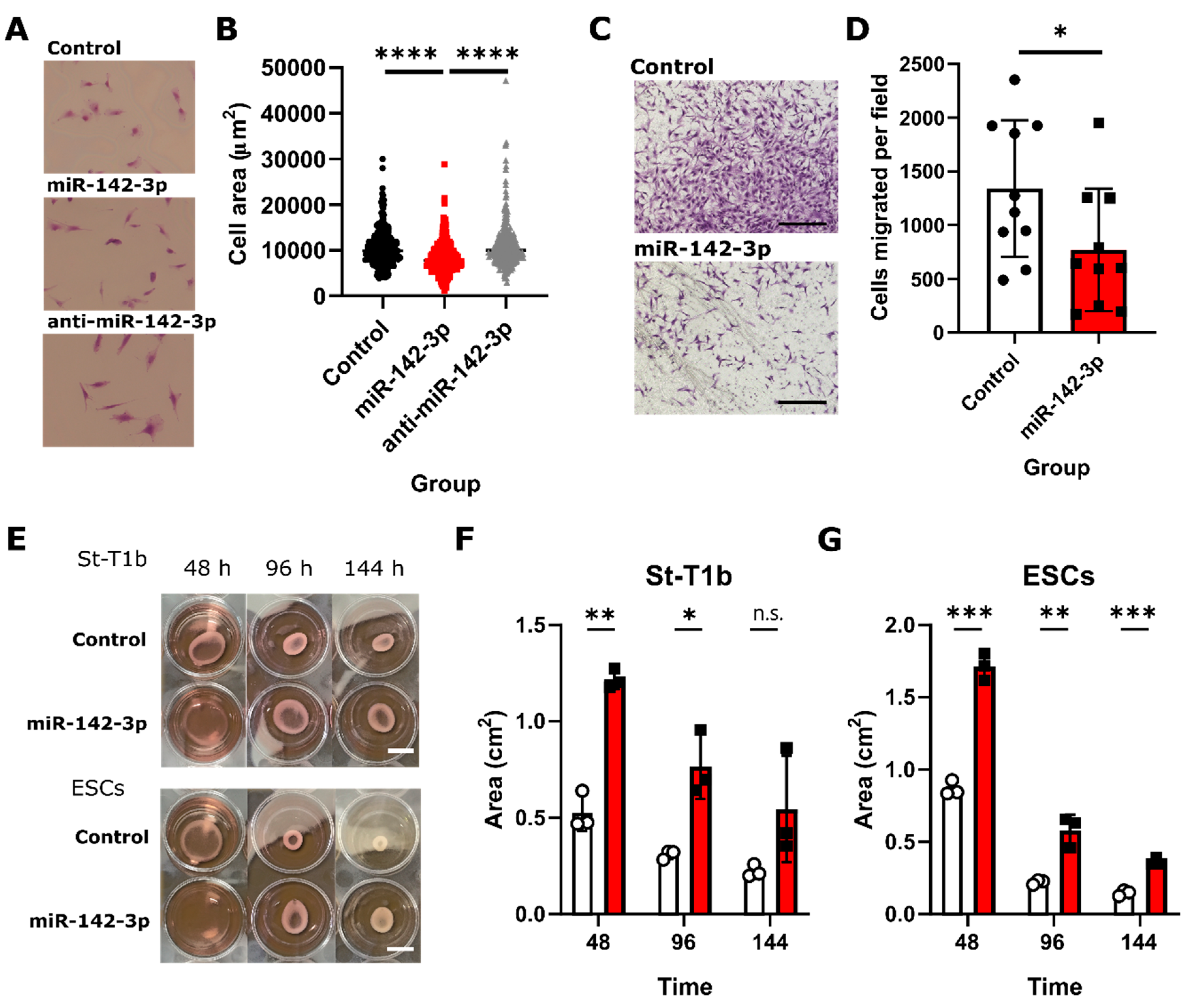
3.6. Functional Analysis of the Role of miRNA-142-3p in Endometriosis Suggests Its Effects on Endometrial Stromal Cell Size, Migration, and Collagen I Contractility
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability Statement
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Patient Age at Biopsy | rASRM Score | Endometriosis Manifestations | Location of the Biopsy |
|---|---|---|---|---|
| #1 | 33 | III | Plica vesicouterina, Lig. Sacrouterinum right. Pelvic wall left, Ovary left | Plica vesicouterina |
| #2 | 35 | IV | Septum rectovaginale. Pelvic wall both sites. Douglas space | Septum rectovaginale |
| #3 | 39 | III | Uterus, Ovary both sites, Pelvic wall both sites, Bladder peritoneum, Douglas space | Uterus |
| #4 | 33 | II | Plica vesicouterina, Pelvic wall left, Lig. Sacrouterinum both sites, Douglas space, Rectum | Plica vesicouterina |
| #5 | 19 | III | Pelvic peritoneum both sites, Douglas space, Septum rectovaginale, Vagina | Pelvic peritoneum |
| #6 | 22 | III | Pelvic wall left, Ligamentum sacrouterinum left, Rectum | Pelvic wall, left |
| #7 | 39 | III | Peritoneum, bladder, pelvic wall close to the urethra | Peritoneum |
| #A | 30 | III | Pelvic wall left, Lig. Sacrouterinum left, Plica vesicouterina, Ovary both sides, Recessus ovarian both sites, Douglas space. Rectal front wall | Eutopic uterine endometrium |
| #B | 37 | III | Pelvic wall both sides, Abdominal front wall left. Ovary right. Douglas space, Rectum | Eutopic uterine endometrium |
References
- Bartel, D.P.; Chen, C.Z. Micromanagers of gene expression: The potentially widespread influence of metazoan microRNAs. Nat. Rev. Genet. 2004, 5, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Hammond, S.M. An overview of microRNAs. Adv. Drug Deliv. Rev. 2015, 87, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croce, C.M. Causes and consequences of microRNA dysregulation in cancer. Nat. Rev. Genet. 2009, 10, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, F.; Moreno-Moya, J.M.; Lozoya, T.; Romero, A.; Martínez, S.; Monterde, M.; Gurrea, M.; Ferri, B.; Núñez, M.J.; Simón, C.; et al. Embryonic miRNA profiles of normal and ectopic pregnancies. PLoS ONE 2014, 9, e102185. [Google Scholar] [CrossRef] [Green Version]
- Ohlsson Teague, E.M.C.; Van der Hoek, K.H.; Van der Hoek, M.B.; Perry, N.; Wagaarachchi, P.; Robertson, S.A.; Print, C.G.; Hull, L.M. MicroRNA-Regulated Pathways Associated with Endometriosis. Mol. Endocrinol. 2009, 23, 265–275. [Google Scholar] [CrossRef]
- Saare, M.; Rekker, K.; Laisk-Podar, T.; Rahmioglu, N.; Zondervan, K.; Salumets, A.; Götte, M.; Peters, M. Challenges in endometriosis miRNA studies—From tissue heterogeneity to disease specific miRNAs. Biochim. Biophys. Acta-Mol. Basis Dis. 2017, 1863, 2282–2292. [Google Scholar] [CrossRef]
- Panir, K.; Schjenken, J.E.; Robertson, S.A.; Hull, M.L. Non-coding RNAs in endometriosis: A narrative review. Hum. Reprod. Update 2018, 24, 497–515. [Google Scholar] [CrossRef] [Green Version]
- Vercellini, P.; Viganò, P.; Somigliana, E.; Fedele, L. Endometriosis: Pathogenesis and treatment. Nat. Rev. Endocrinol. 2014, 10, 261–275. [Google Scholar] [CrossRef]
- Klemmt, P.A.B.; Starzinski-Powitz, A. Biology of Eutopic and Ectopic Endometrium in Women with Endometriosis. In Endometriosis; Wiley-Blackwell: Oxford, UK, 2012; pp. 115–129. [Google Scholar]
- Kao, A.-P.; Wang, K.-H.; Chang, C.-C.; Lee, J.-N.; Long, C.-Y.; Chen, H.-S.; Tsai, C.-F.; Hsieh, T.-H.; Tsai, E.-M. Comparative study of human eutopic and ectopic endometrial mesenchymal stem cells and the development of an in vivo endometriotic invasion model. Fertil. Steril. 2011, 95, 1308–1315. [Google Scholar] [CrossRef]
- Laudanski, P.; Charkiewicz, R.; Kuzmicki, M.; Szamatowicz, J.; Charkiewicz, A.; Niklinski, J. MicroRNAs expression profiling of eutopic proliferative endometrium in women with ovarian endometriosis. Reprod. Biol. Endocrinol. 2013, 11, 78. [Google Scholar] [CrossRef] [Green Version]
- Patel, B.G.; Rudnicki, M.; Yu, J.; Shu, Y.; Taylor, R.N. Progesterone resistance in endometriosis: Origins, consequences and interventions. Acta Obstet. Gynecol. Scand. 2017, 96, 623–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulun, S.E.; Zeitoun, K.; Takayama, K.; Noble, L.; Michael, D.; Simpson, E.; Johns, A.; Putman, M.; Sasano, H. Estrogen production in endometriosis and use of aromatase inhibitors to treat endometriosis. Endocr. Relat. Cancer 1999, 6, 293–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuge, A.; Nasu, K.; Matsumoto, H.; Nishida, M.; Narahara, H. Collagen gel contractility is enhanced in human endometriotic stromal cells: A possible mechanism underlying the pathogenesis of endometriosis-associated fibrosis. Hum. Reprod. 2007, 22, 938–944. [Google Scholar] [CrossRef] [PubMed]
- Orellana, R.; García-Solares, J.; Donnez, J.; van Kerk, O.; Dolmans, M.-M.; Donnez, O. Important role of collective cell migration and nerve fiber density in the development of deep nodular endometriosis. Fertil. Steril. 2017, 107, 987–995.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohlsson Teague, E.M.C.; Print, C.G.; Hull, M.L. The role of microRNAs in endometriosis and associated reproductive conditions. Hum. Reprod. Update 2009, 16, 142–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chelariu-Raicu, A.; Wilke, C.; Brand, M.; Starzinski-Powitz, A.; Kiesel, L.; Schüring, A.N.; Götte, M. Syndecan-4 expression is upregulated in endometriosis and contributes to an invasive phenotype. Fertil. Steril. 2016, 106, 378–385. [Google Scholar] [CrossRef] [Green Version]
- Klemmt, P.A.B.; Starzinski-Powitz, A. Molecular and Cellular Pathogenesis of Endometriosis. Curr. Womens Health Rev. 2018, 14, 106–116. [Google Scholar] [CrossRef]
- Ma, L.; Li, Z.; Li, W.; Ai, J.; Chen, X. MicroRNA-142-3p suppresses endometriosis by regulating KLF9-mediated autophagy in vitro and in vivo. RNA Biol. 2019, 1–16. [Google Scholar] [CrossRef]
- Kästingschäfer, C.S.; Schäfer, S.D.; Kiesel, L.; Götte, M. MiR-142-3p is a novel regulator of cell viability and proinflammatory signalling in endometrial stroma cells. Reprod. Biomed. Online 2015, 30, 553–556. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Li, X.; He, Q.; Gao, J.; Gao, Y.; Liu, B.; Liu, F. MiR-142-3p regulates the formation and differentiation of hematopoietic stem cells in vertebrates. Cell Res. 2013, 23, 1356–1368. [Google Scholar] [CrossRef] [Green Version]
- Godfrey, J.D.; Morton, J.P.; Wilczynska, A.; Sansom, O.J.; Bushell, M.D. MiR-142-3p is downregulated in aggressive p53 mutant mouse models of pancreatic ductal adenocarcinoma by hypermethylation of its locus article. Cell Death Dis. 2018, 9, 644. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.F.; Chen, Z.Y.; Wang, L.; Wang, M.; Liu, X.H. MiR-142-3p functions as an oncogene in prostate cancer by targeting FOXO1. J. Cancer 2020, 11, 1614–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwickert, A.; Weghake, E.; Brüggemann, K.; Engbers, A.; Brinkmann, B.F.; Kemper, B.; Seggewiß, J.; Stock, C.; Ebnet, K.; Kiesel, L.; et al. microRNA miR-142-3p Inhibits Breast Cancer Cell Invasiveness by Synchronous Targeting of WASL, Integrin Alpha V, and Additional Cytoskeletal Elements. PLoS ONE 2015, 10, e0143993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troschel, F.M.; Böhly, N.; Borrmann, K.; Braun, T.; Schwickert, A.; Kiesel, L.; Eich, H.T.; Götte, M.; Greve, B. miR-142-3p attenuates breast cancer stem cell characteristics and decreases radioresistance in vitro. Tumour Biol. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.; Kim, S.E.; Yano, H.; Matsumoto, G.; Ohuchida, R.; Ishikura, Y.; Araki, M.; Araki, K.; Park, S.; Komatsu, T.; et al. MiR-142 Is Required for Staphylococcus aureus Clearance at Skin Wound Sites via Small GTPase-Mediated Regulation of the Neutrophil Actin Cytoskeleton. J. Investig. Dermatol. 2017, 137, 931–940. [Google Scholar] [CrossRef]
- Samalecos, A.; Reimann, K.; Wittmann, S.; Schulte, H.M.; Brosens, J.J.; Bamberger, A.M.; Gellersen, B. Characterization of a novel telomerase-immortalized human endometrial stromal cell line, St-T1 b. Reprod. Biol. Endocrinol. 2009. [Google Scholar] [CrossRef] [Green Version]
- Schüring, A.N.; Schulte, N.; Kelsch, R.; Röpke, A.; Kiesel, L.; Götte, M. Characterization of endometrial mesenchymal stem-like cells obtained by endometrial biopsy during routine diagnostics. Fertil. Steril. 2011, 95, 423–426. [Google Scholar] [CrossRef]
- Ramirez Williams, L.; Brüggemann, K.; Hubert, M.; Achmad, N.; Kiesel, L.; Schäfer, S.D.; Greve, B.; Götte, M. γ-Secretase inhibition affects viability, apoptosis, and the stem cell phenotype of endometriotic cells. Acta Obstet. Gynecol. Scand. 2019. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Liu, W.; Wang, X. Prediction of functional microRNA targets by integrative modeling of microRNA binding and target expression data. Genome Biol. 2019. [Google Scholar] [CrossRef]
- Mi, H.; Muruganujan, A.; Ebert, D.; Huang, X.; Thomas, P.D. PANTHER version 14: More genomes, a new PANTHER GO-slim and improvements in enrichment analysis tools. Nucleic Acids Res. 2019, 47, D419–D426. [Google Scholar] [CrossRef] [PubMed]
- Humphries, J.D.; Wang, P.; Streuli, C.; Geiger, B.; Humphries, M.J.; Ballestrem, C. Vinculin controls focal adhesion formation by direct interactions with talin and actin. J. Cell Biol. 2007, 179, 1043–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morandi, E.M.; Verstappen, R.; Zwierzina, M.E.; Geley, S.; Pierer, G.; Ploner, C. ITGAV and ITGA5 diversely regulate proliferation and adipogenic differentiation of human adipose derived stem cells. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laganà, A.S.; Garzon, S.; Götte, M.; Viganò, P.; Franchi, M.; Ghezzi, F.; Martin, D.C. The pathogenesis of endometriosis: Molecular and cell biology insights. Int. J. Mol. Sci. 2019, 20, 5615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, S.M.; Thompson, J.A.; Ufkin, M.L.; Sathyanarayana, P.; Liaw, L.; Congdon, C.B. Common features of microRNA target prediction tools. Front. Genet. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Reimand, J.; Isserlin, R.; Voisin, V.; Kucera, M.; Tannus-Lopes, C.; Rostamianfar, A.; Wadi, L.; Meyer, M.; Wong, J.; Xu, C.; et al. Pathway enrichment analysis and visualization of omics data using g:Profiler, GSEA, Cytoscape and EnrichmentMap. Nat. Protoc. 2019, 14, 482–517. [Google Scholar] [CrossRef] [PubMed]
- Zigmond, S.H. How WASP regulates actin polymerization. J. Cell Biol. 2000, 150, F117–F120. [Google Scholar] [CrossRef] [Green Version]
- Alekhina, O.; Burstein, E.; Billadeau, D.D. Cellular functions of WASP family proteins at a glance. J. Cell. Sci. 2017, 130, 2235–2241. [Google Scholar] [CrossRef] [Green Version]
- Kyama, C.M.; Overbergh, L.; Mihalyi, A.; Meuleman, C.; Mwenda, J.M.; Mathieu, C.; D’Hooghe, T.M. Endometrial and peritoneal expression of aromatase, cytokines, and adhesion factors in women with endometriosis. Fertil. Steril. 2008, 89, 301–310. [Google Scholar] [CrossRef]
- Mackay, D.J.G.; Esch, F.; Furthmayr, H.; Hall, A. Rho- and Rac-dependent assembly of focal adhesion complexes and actin filaments in permeabilized fibroblasts: An essential role for ezrin/radixin/moesin proteins. J. Cell Biol. 1997, 138, 927–938. [Google Scholar] [CrossRef] [Green Version]
- Wozniak, M.A.; Modzelewska, K.; Kwong, L.; Keely, P.J. Focal adhesion regulation of cell behavior. Biochim. Biophys. Acta Mol. Cell Res. 2004, 1692, 103–119. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, D.E.; Martin, M.M.; Feldman, D.S.; Terry, A.V.; Nuovo, G.J.; Elton, T.S. Experimental validation of miRNA targets. Methods 2008, 44, 47–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi-wen, X.; Liu, S.; Eastwood, M.; Sonnylal, S.; Denton, C.P.; Abraham, D.J.; Leask, A. Rac Inhibition Reverses the Phenotype of Fibrotic Fibroblasts. PLoS ONE 2009, 4, e7438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyhani, V.; Tsioumpekou, M.; Van Wieringen, T.; Rask, L.; Lennartsson, J.; Rubin, K. PDGF-BB enhances collagen gel contraction through a PI3 K-PLCγ-PKC-cofilin pathway. Sci. Rep. 2017, 7, 8924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsang, F.H.C.; Au, S.L.K.; Wei, L.; Fan, D.N.Y.; Lee, J.M.F.; Wong, C.C.L.; Ng, I.O.L.; Wong, C.M. MicroRNA-142-3p and microRNA-142-5 p are downregulated in hepatocellular carcinoma and exhibit synergistic effects on cell motility. Front. Med. 2015, 9, 331–343. [Google Scholar] [CrossRef]
- Liu, J.; Li, W.; Wang, S.; Wu, Y.; Li, Z.; Wang, W.; Liu, R.; Ou, J.; Zhang, C.; Wang, S. MiR-142-3p Attenuates the Migration of CD4+ T Cells through Regulating Actin Cytoskeleton via RAC1 and ROCK2 in Arteriosclerosis Obliterans. PLoS ONE 2014, 9, e95514. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Börschel, C.S.; Stejskalova, A.; Schäfer, S.D.; Kiesel, L.; Götte, M. miR-142-3p Reduces the Size, Migration, and Contractility of Endometrial and Endometriotic Stromal Cells by Targeting Integrin- and Rho GTPase-Related Pathways That Regulate Cytoskeletal Function. Biomedicines 2020, 8, 291. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8080291
Börschel CS, Stejskalova A, Schäfer SD, Kiesel L, Götte M. miR-142-3p Reduces the Size, Migration, and Contractility of Endometrial and Endometriotic Stromal Cells by Targeting Integrin- and Rho GTPase-Related Pathways That Regulate Cytoskeletal Function. Biomedicines. 2020; 8(8):291. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8080291
Chicago/Turabian StyleBörschel, Christin S., Anna Stejskalova, Sebastian D. Schäfer, Ludwig Kiesel, and Martin Götte. 2020. "miR-142-3p Reduces the Size, Migration, and Contractility of Endometrial and Endometriotic Stromal Cells by Targeting Integrin- and Rho GTPase-Related Pathways That Regulate Cytoskeletal Function" Biomedicines 8, no. 8: 291. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8080291