Inflammation in Dry Eye Syndrome: Identification and Targeting of Oxylipin-Mediated Mechanisms

 , , ,
, , ,
Abstract
:1. Introduction
2. Experimental Section
2.1. Materials
2.2. Experimental Animals and Ethics Statement
2.3. Experimental Model
2.4. Histological Analysis
2.5. Schirmer’s Test
2.6. TF Collection
2.7. Total Protein Concentration Measurement
2.8. Total Antioxidant Activity Analysis
2.9. Antioxidant Enzymes Activity and Cytokine Concentration Analysis
2.10. Lipid Extraction
2.11. UPLC-MS/MS Analysis
2.12. Statistical Analysis
3. Results
3.1. Dynamics of Inflammation in DES: Tear Production and Morphological Changes in the Cornea
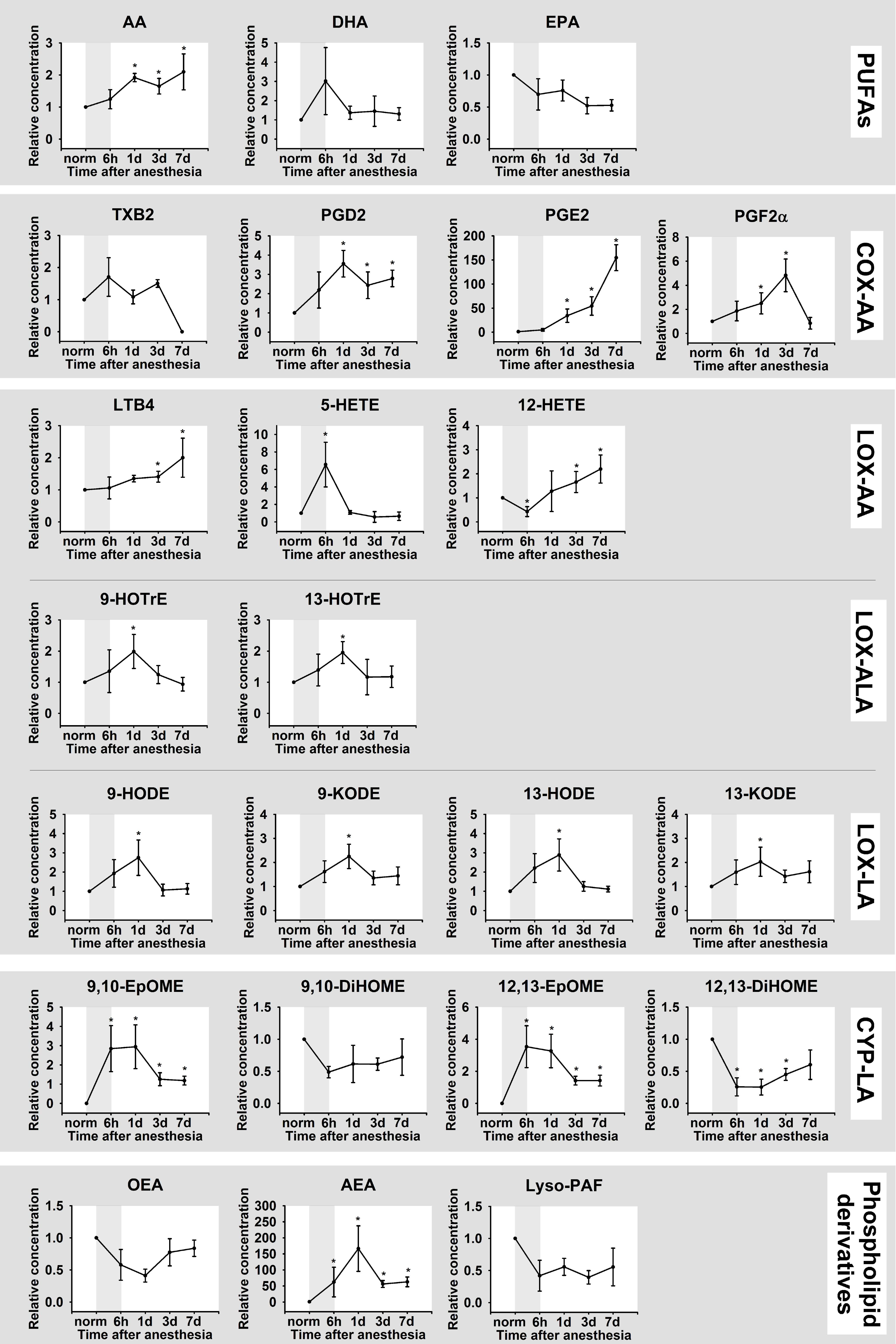
3.2. Dynamics of Inflammation in DES: Biochemistry and Lipidomics of the Tear Fluid
3.3. Selective Targeting of Inflammation in DES: Rationale for the Drug Formulation
3.4. Selective Targeting of Inflammation in DES: A Morphological Study
3.5. Selective Targeting of Inflammation in DES: Biochemistry and Lipidomics of the Tear Fluid
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Arachidonic acid |
| AEA | N-arachidonoylethanolamine |
| ALA | Alpha-linolenic acid |
| BCA | Bicinchoninic acid |
| COX | Cyclooxygenases |
| CYP | Cytochrome P450 monooxygenases |
| DES | Dry eye syndrome |
| DHA | Docosahexaenoic acids |
| DMSO | Dimethyl sulfoxide |
| 9,10-DiHOME | 9,10-dihydroxyoctadecamonoenoic acid |
| 12,13-DiHOME | 12,13-dihydroxyoctadecamonoenoic acid |
| EPA | Eicosapentaenoic acid |
| 9,10-EpOME | 9,10-epoxyoctadecamonoenoic acid |
| 12,13-EpOME | 12,13-epoxyoctadecamonoenoic acid |
| 9-HODE | 9-hydroxyoctadecadienoic acid |
| 13-HODE | 13-hydroxyoctadecadienoic acid |
| IL-1 | Interleukin-1 |
| IL-10 | Interleukin-10 |
| IL-1β | Interleukin-1 beta |
| 9-KODE | 9-oxo-octadecadienoic acid |
| 13-KODE | 13-oxo-octadecadienoic acid |
| LTB4 | Leukotriene B4 |
| LA | Linoleic acid |
| LOX | Lipoxygenase |
| lyso-PAF | Lyso-Platelet-Activating Factor (1-O-hexadecyl-sn-glyceryl-3-phosphorylcholine) |
| MGD | Meibomian gland dysfunction |
| NSAID | Nonsteroidal anti-inflammatory drug |
| OEA | Oleoylethanolamine ((Z)-N-(2-Hydroxyethyl)octadec-9-enamide) |
| PBS | Phosphate-buffered saline |
| PUFA | Polyunsaturated fatty acid |
| PGD2 | Prostaglandin D2 |
| PGE2 | Prostaglandin E2 |
| PGF2α | Prostaglandin F2 alpha |
| ROS | Reactive oxygen species |
| TF | Tear fluid |
| TNF-α | Tumor necrosis factor alpha |
| UPLC-MS/MS | Ultra performance liquid chromatography-tandem mass spectrometry |
References
- Bron, A.J.; De Paiva, C.S.; Chauhan, S.K.; Bonini, S.; Gabison, E.E.; Jain, S.; Knop, E.; Markoulli, M.; Ogawa, Y.; Perez, V.; et al. TFOS DEWS II pathophysiology report. Ocul. Surf. 2017, 15, 438–510. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, F.; Alves, M.; Bunya, V.Y.; Jalbert, I.; Lekhanont, K.; Malet, F.; Na, K.-S.; Schaumberg, D.; Uchino, M.; Vehof, J.; et al. TFOS DEWS II Epidemiology Report. Ocul. Surf. 2017, 15, 334–365. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T. Inflammatory Response in Dry Eye. Investig. Opthalmol. Vis. Sci. 2018, 59, DES192–DES199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malafa, M.M.; Coleman, J.E.; Bowman, R.W.; Rohrich, R.J. Perioperative Corneal Abrasion: Updated Guidelines for Prevention and Management. Plast. Reconstr. Surg. 2016, 137, 790e–798e. [Google Scholar] [CrossRef] [PubMed]
- Kara-Junior, N.; De Espíndola, R.F.; Filho, J.V.; Rosa, C.P.; Ottoboni, A.; Silva, E.D. Ocular risk management in patients undergoing general anesthesia: An analysis of 39,431 surgeries. Clinics 2015, 70, 541–543. [Google Scholar] [CrossRef]
- Zernii, E.Y.; Gancharova, O.S.; Baksheeva, V.E.; Golovastova, M.O.; Kabanova, E.I.; Savchenko, M.S.; Tiulina, V.V.; Sotnikova, L.F.; Zamyatnin, J.A.A.; Philippov, P.P.; et al. Mitochondria-Targeted Antioxidant SkQ1 Prevents Anesthesia-Induced Dry Eye Syndrome. Oxid. Med. Cell. Longev. 2017, 2017, 9281519. [Google Scholar] [CrossRef]
- Baksheeva, V.E.; Gancharova, O.S.; Tiulina, V.V.; Iomdina, E.N.; Zamyatnin, J.A.A.; Philippov, P.P.; Zernii, E.Y.; Senin, I.I. Iatrogenic Damage of Eye Tissues: Current Problems and Possible Solutions. Biochemistry 2018, 83, 1563–1574. [Google Scholar] [CrossRef]
- The Definition and Classification of Dry Eye Disease: Report of the Definition and Classification Subcommittee of the International Dry Eye Workshop. Ocul. Surf. 2007, 5, 75–92. [CrossRef]
- Skalicky, S.E.; Petsoglou, C.; Gurbaxani, A.; Fraser, C.L.; McCluskey, P. New Agents for Treating Dry Eye Syndrome. Curr. Allergy Asthma Rep. 2012, 13, 322–328. [Google Scholar] [CrossRef]
- Zernii, E.Y.; Baksheeva, V.E.; Yani, E.V.; Philippov, P.P.; Senin, I.I. Therapeutic Proteins for Treatment of Corneal Epithelial Defects. Curr. Med. Chem. 2019, 26, 517–545. [Google Scholar] [CrossRef]
- Messmer, E.M. The Pathophysiology, Diagnosis, and Treatment of Dry Eye Disease. Dtsch. Aerzteblatt Online 2015, 112, 71–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.Q.; Luo, L.; Chen, Z.; Kim, H.S.; Song, X.J.; Pflugfelder, S.C. JNK and ERK MAP kinases mediate induction of IL-1beta, TNF-alpha and IL-8 following hyperosmolar stress in human limbal epithelial cells. Exp. Eye Res. 2006, 82, 588–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, V.L.; Pflugfelder, S.C.; Zhang, S.; Shojaei, A.; Haque, R. Lifitegrast, a Novel Integrin Antagonist for Treatment of Dry Eye Disease. Ocul. Surf. 2016, 14, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Li, W.; Dong, N.; Chen, W.; Liu, J.; Chen, L.; Yuan, H.; Geng, Z.; Liu, Z. Changes in Corneal Epithelial Layer Inflammatory Cells in Aqueous Tear–Deficient Dry Eye. Investig. Opthalmol. Vis. Sci. 2010, 51, 122–128. [Google Scholar] [CrossRef]
- Matsuda, S.; Koyasu, S. Mechanisms of action of cyclosporine. Immunopharmacology 2000, 47, 119–125. [Google Scholar] [CrossRef]
- Colligris, B.; Alkozi, H.A.; Pintor, J. Recent developments on dry eye disease treatment compounds. Saudi J. Ophthalmol. 2013, 28, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Saijyothi, A.V.; Fowjana, J.; Madhumathi, S.; Rajeshwari, M.; Thennarasu, M.; Prema, P.; Angayarkanni, N. Tear fluid small molecular antioxidants profiling shows lowered glutathione in keratoconus. Exp. Eye Res. 2012, 103, 41–46. [Google Scholar] [CrossRef]
- Crouch, R.K.; Goletz, P.; Snyder, A.; Coles, W.H. Antioxidant Enzymes in Human Tears. J. Ocul. Pharmacol. Ther. 1991, 7, 253–258. [Google Scholar] [CrossRef]
- Dogru, M.; Kojima, T.; Simsek, C.; Tsubota, K. Potential Role of Oxidative Stress in Ocular Surface Inflammation and Dry Eye Disease. Investig. Opthalmol. Vis. Sci. 2018, 59, DES163–DES168. [Google Scholar] [CrossRef] [Green Version]
- Gabbs, M.; Leng, S.; Devassy, J.G.; Monirujjaman, M.; Aukema, H.M. Advances in Our Understanding of Oxylipins Derived from Dietary PUFAs. Adv. Nutr. 2015, 6, 513–540. [Google Scholar] [CrossRef] [Green Version]
- Walter, S.D.; Gronert, K.; McClellan, A.L.; Levitt, R.C.; Sarantopoulos, K.D.; Galor, A. omega-3 Tear Film Lipids Correlate with Clinical Measures of Dry Eye. Investig. Ophtamol. Vis. Sci. 2016, 57, 2472–2478. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.; Park, C.; Lee, H.S.; Park, M.S.; Lim, H.T.; Chauhan, S.; Dana, R.; Lee, H.; Lee, H.K. Change in Prostaglandin Expression Levels and Synthesizing Activities in Dry Eye Disease. Ophthalmology 2012, 119, 2211–2219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lekhanont, K.; Sathianvichitr, K.; Pisitpayat, P.; Anothaisintawee, T.; Soontrapa, K.; Udomsubpayakul, U. Association between the levels of prostaglandin E2 in tears and severity of dry eye. Int. J. Ophthalmol. 2019, 12, 1127–1133. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.W.; Seo, Y.; Choi, W.; Yeo, A.; Noh, H.; Kim, E.K.; Lee, H.K. Dry Eye-Induced CCR7+CD11b+ Cell Lymph Node Homing Is Induced by COX-2 Activities. Investig. Opthalmol. Vis. Sci. 2014, 55, 6829–6838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigas, B.; Huang, W.; Honkanen, R. NSAID-induced corneal melt: Clinical importance, pathogenesis, and risk mitigation. Surv. Ophthalmol. 2020, 65, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guidera, A.C.; I Luchs, J.; Udell, I.J. Keratitis, ulceration, and perforation associated with topical nonsteroidal anti-inflammatory drugs. Ophthalmology 2001, 108, 936–944. [Google Scholar] [CrossRef]
- Zernii, E.Y.; Golovastova, M.O.; Baksheeva, V.E.; Kabanova, E.; Ishutina, I.E.; Gancharova, O.S.; Gusev, A.E.; Savchenko, M.S.; Loboda, A.P.; Sotnikova, L.F.; et al. Alterations in tear biochemistry associated with postanesthetic chronic dry eye syndrome. Biochemistry 2016, 81, 1549–1557. [Google Scholar] [CrossRef]
- White, E.; Crosse, M.M. The aetiology and prevention of peri-operative corneal abrasions. Anaesthesia 1998, 53, 157–161. [Google Scholar] [CrossRef]
- Chistyakov, D.V.; Azbukina, N.V.; Astakhova, A.A.; Goriainov, S.V.; Chistyakov, V.V.; Tiulina, V.V.; Baksheeva, V.E.; Kotelin, V.I.; Fedoseeva, E.V.; Zamyatnin, A.A., Jr.; et al. Comparative lipidomic analysis of inflammatory mediators in the aqueous humor and tear fluid of humans and rabbits. Metabolomics 2020, 16, 27. [Google Scholar] [CrossRef]
- Zernii, E.Y.; Baksheev, V.E.; Kabanova, E.I.; Tiulina, V.V.; Golovastova, M.O.; Gancharova, O.S.; Savchenko, M.S.; Sotikova, L.F.; Zamyatnin, J.A.A.; Filippov, P.P.; et al. Effect of General Anesthesia Duration on Recovery of Secretion and Biochemical Properties of Tear Fluid in the Post-Anesthetic Period. Bull. Exp. Biol. Med. 2018, 165, 269–271. [Google Scholar] [CrossRef]
- Zernii, E.Y.; Gancharova, O.S.; Tiulina, V.V.; Zamyatnin, J.A.A.; Philippov, P.P.; Baksheeva, V.E.; Senin, I.I. Mitochondria-targeted antioxidant SKQ1 protects cornea from oxidative damage induced by ultraviolet irradiation and mechanical injury. BMC Ophthalmol. 2018, 18, 336. [Google Scholar] [CrossRef] [PubMed]
- Chistyakov, D.; Azbukina, N.; Goriainov, S.; Gancharova, O.; Tiulina, V.; Baksheeva, V.; Iomdina, E.; Philippov, P.; Sergeeva, M.; Senin, I.; et al. Inflammatory metabolites of arahidonic acid in tear fluid in UV-induced corneal damage. Biomed. Khidm. 2019, 65, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Baksheeva, V.E.; Tiulina, V.V.; Tikhomirova, N.K.; Gancharova, O.S.; Komarov, S.V.; Philippov, P.P.; Zamyatnin, J.A.A.; Senin, I.I.; Zernii, E.Y. Suppression of Light-Induced Oxidative Stress in the Retina by Mitochondria-Targeted Antioxidant. Antioxidants 2018, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Chistyakov, D.V.; Baksheeva, V.E.; Tiulina, V.V.; Goriainov, S.V.; Azbukina, N.V.; Gancharova, O.S.; Arifulin, E.A.; Komarov, S.V.; Chistyakov, V.V.; Tikhomirova, N.K.; et al. Mechanisms and Treatment of Light-Induced Retinal Degeneration-Associated Inflammation: Insights from Biochemical Profiling of the Aqueous Humor. Int. J. Mol. Sci. 2020, 21, 704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chistyakov, D.V.; Grabeklis, S.; Goriainov, S.V.; Chistyakov, V.V.; Sergeeva, M.G.; Reiser, G. Astrocytes synthesize primary and cyclopentenone prostaglandins that are negative regulators of their proliferation. Biochem. Biophys. Res. Commun. 2018, 500, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Bouchette, D.; Preuss, C.V. Zileuton; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Rossi, A.; Pergola, C.; Koeberle, A.; Hoffmann, M.; Dehm, F.; Bramanti, P.; Cuzzocrea, S.; Werz, O.; Sautebin, L. The 5-lipoxygenase inhibitor, zileuton, suppresses prostaglandin biosynthesis by inhibition of arachidonic acid release in macrophages. Br. J. Pharmacol. 2010, 161, 555–570. [Google Scholar] [CrossRef] [Green Version]
- Pelletier, J.S.; Devine, J.; Capriotti, K.; Barone, S.B.; Capriotti, J.A. Topical application of povidone-iodine/dimethylsulfoxide ophthalmic gel preparation in Dutch-Belted rabbits. Cutan. Ocul. Toxicol. 2019, 38, 221–226. [Google Scholar] [CrossRef]
- Gordon, D.M. Dimethyl sulfoxide in ophthalmology, with special reference to possible toxic effects. Ann. N. Y. Acad. Sci. 1967, 141, 392–402. [Google Scholar] [CrossRef]
- Maibach, H.I.; Feldmann, R.J. The effect of DMSO on percutaneous penetration of hydrocortisone and testosterone in. Ann. N. Y. Acad. Sci. 1967, 141, 423–427. [Google Scholar] [CrossRef]
- Capriotti, K.; Capriotti, J.A. Dimethyl sulfoxide: History, chemistry, and clinical utility in dermatology. J. Clin. Aesthet. Dermatol. 2012, 5, 24–26. [Google Scholar]
- Hanna, C.; Fraunfelder, F.T.; Meyer, S.M. Effects of dimethyl sulfoxide on ocular inflammation. Ann. Ophthalmol. 1977, 9, 61. [Google Scholar] [PubMed]
- Zileuton: Cayman Chemical Product Information. Available online: https://www.caymanchem.com/pdfs/10006967.pdf (accessed on 1 June 2020).
- Del Castillo, F.B.; Cantu-Dibildox, J.; Sanz-González, S.M.; Zanon-Moreno, V.C.; Pinazo-Durán, M.D. Cytokine expression in tears of patients with glaucoma or dry eye disease: A prospective, observational cohort study. Eur. J. Ophthalmol. 2018, 29, 437–443. [Google Scholar] [CrossRef]
- Ambaw, Y.A.; Chao, C.; Ji, S.; Raida, M.; Torta, F.; Wenk, M.R.; Tong, L. Tear eicosanoids in healthy people and ocular surface disease. Sci. Rep. 2018, 8, 11296. [Google Scholar] [CrossRef] [PubMed]
- Altinsoy, A.; Dileköz, E.; Kul, O.; Ilhan, S.Ö.; Tunccan, O.G.; Seven, I.; Bagriacik, E.U.; Sarioglu, Y.; Or, M.; Ercan, Z.S. A Cannabinoid Ligand, Anandamide, Exacerbates Endotoxin-Induced Uveitis in Rabbits. J. Ocul. Pharmacol. Ther. 2011, 27, 545–552. [Google Scholar] [CrossRef]
- Vangaveti, V.; Baune, B.T.; Kennedy, R.L. Hydroxyoctadecadienoic acids: Novel regulators of macrophage differentiation and atherogenesis. Ther. Adv. Endocrinol. Metab. 2010, 1, 51–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, Y.; Umeno, A.; Ogawa, Y.; Shichiri, M.; Murotomi, K.; Horie, M. Chemistry of Lipid Peroxidation Products and Their Use as Biomarkers in Early Detection of Diseases. J. Oleo Sci. 2015, 64, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Sevanian, A.; Mead, J.F.; Stein, R.A. Epoxides as products of lipid autoxidation in rat lungs. Lipids 1979, 14, 634–643. [Google Scholar] [CrossRef]
- Thompson, D.; Hammock, B.D. Dihydroxyoctadecamonoenoate esters inhibit the neutrophil respiratory burst. J. Biosci. 2007, 32, 279–291. [Google Scholar] [CrossRef]
- Seen, S.; Tong, L. Dry eye disease and oxidative stress. Acta Ophthalmol. 2017, 96, e412–e420. [Google Scholar] [CrossRef] [Green Version]
- Perez-Garmendia, R.; Rodriguez, A.L.D.E.; Ramos-Martinez, I.; Zuñiga, N.M.; Gonzalez-Salinas, R.; Quiroz-Mercado, H.; Zenteno, E.; Hernández, E.R.; Hernández-Zimbrón, L.F.; Martínez-Ramos, I. Interplay between Oxidative Stress, Inflammation, and Amyloidosis in the Anterior Segment of the Eye; Its Pathological Implications. Oxid. Med. Cell. Longev. 2020, 2020, 6286105. [Google Scholar] [CrossRef]
- Kojima, T.; Wakamatsu, T.H.; Dogru, M.; Ogawa, Y.; Igarashi, A.; Ibrahim, O.M.A.; Inaba, T.; Shimizu, T.; Noda, S.; Obata, H.; et al. Age-Related Dysfunction of the Lacrimal Gland and Oxidative Stress: Evidence from the Cu,Zn-superoxide dismutase-1 (Sod1) knockout mice. Am. J. Pathol. 2012, 180, 1879–1896. [Google Scholar] [CrossRef] [PubMed]
- Caligiuri, S.P.; Love, K.; Winter, T.; Gauthier, J.; Taylor, C.G.; Blydt-Hansen, T.; Zahradka, P.; Aukema, H.M. Dietary Linoleic Acid and α-Linolenic Acid Differentially Affect Renal Oxylipins and Phospholipid Fatty Acids in Diet-Induced Obese Rats. J. Nutr. 2013, 143, 1421–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, N.L.; Dunford, P.J.; Xue, X.; Jiang, X.; Lundeen, K.A.; Coles, F.; Riley, J.P.; Williams, K.N.; Grice, C.A.; Edwards, J.P.; et al. Anti-Inflammatory Activity of a Potent, Selective Leukotriene A4 Hydrolase Inhibitor in Comparison with the 5-Lipoxygenase Inhibitor Zileuton. J. Pharmacol. Exp. Ther. 2007, 321, 1154–1160. [Google Scholar] [CrossRef] [PubMed]
- Suh, J.H.; Yum, E.K.; Cho, Y.S. Synthesis and Biological Evaluation of N-Aryl-5-aryloxazol-2-amine Derivatives as 5-Lipoxygenase Inhibitors. Chem. Pharm. Bull. 2015, 63, 573–578. [Google Scholar] [CrossRef] [Green Version]
- Altan, S.; Oğurtan, Z. Dimethyl sulfoxide but not indomethacin is efficient for healing in hydrofluoric acid eye burns. J. Int. Soc. Burn. Inj. 2017, 43, 232–244. [Google Scholar] [CrossRef]
- Altan, S.; Sağsöz, H.; Oğurtan, Z. Topical dimethyl sulfoxide inhibits corneal neovascularization and stimulates corneal repair in rabbits following acid burn. Biotech. Histochem. 2017, 92, 619–636. [Google Scholar] [CrossRef]
- Toczołowski, J.; Wolski, T.; Klamut-Sory, K. Effect of drugs inhibiting the formation of hydroxide radicals on healing of experimental corneal ulcer. Klin. Ocz. 1992, 94, 83. [Google Scholar]
- Skrypuch, O.W.; Tokarewicz, A.C.; Willis, N.R. Effects of dimethyl sulfoxide on a model of corneal alkali injury. Can. J. Ophthalmol. 1987, 22, 17–20. [Google Scholar]
- Elisia, I.; Nakamura, H.; Lam, V.; Hofs, E.; Cederberg, R.; Cait, J.; Hughes, M.R.; Lee, L.; Jia, W.; Adomat, H.H.; et al. DMSO Represses Inflammatory Cytokine Production from Human Blood Cells and Reduces Autoimmune Arthritis. PLoS ONE 2016, 11, e0152538. [Google Scholar] [CrossRef] [Green Version]
- Nagy, L.; Tontonoz, P.; Alvarez, J.G.; Chen, H.; Evans, R.M. Oxidized LDL regulates macrophage gene expression through ligand activation of PPARgamma. Cell 1998, 93, 229–240. [Google Scholar] [CrossRef] [Green Version]
- AlSalem, M.; Wong, A.; Millns, P.; Arya, P.H.; Chan, M.S.L.; Bennett, A.J.; Barrett, D.A.; Chapman, V.; Kendall, D.A. The contribution of the endogenous TRPV1 ligands 9-HODE and 13-HODE to nociceptive processing and their role in peripheral inflammatory pain mechanisms. Br. J. Pharmacol. 2013, 168, 1961–1974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balicki, I. Clinical study on the application of tacrolimus and DMSO in the treatment of chronic superficial keratitis in dogs. Pol. J. Vet. Sci. 2012, 15, 667–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelletier, J.S.; Stewart, K.P.; Capriotti, K.; Capriotti, J.A. Rosacea Blepharoconjunctivitis Treated with a Novel Preparation of Dilute Povidone Iodine and Dimethylsulfoxide: A Case Report and Review of the Literature. Ophthalmol. Ther. 2015, 4, 143–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelletier, J.S.; Capriotti, K.; Stewart, K.S.; Capriotti, J.A. Demodex Blepharitis Treated with a Novel Dilute Povidone-Iodine and DMSO System: A Case Report. Ophthalmol. Ther. 2017, 6, 361–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhalerao, H.; Koteshwara, K.; Chandran, S. Brinzolamide Dimethyl Sulfoxide in Situ Gelling Ophthalmic Solution: Formulation Optimisation and In Vitro and In Vivo Evaluation. AAPS PharmSciTech 2020, 21, 69. [Google Scholar] [CrossRef]
- Roda, M.; Corazza, I.; Bacchi-Reggiani, M.L.; Pellegrini, M.; Taroni, L.; Giannaccare, G.; Versura, P. Dry Eye Disease and Tear Cytokine Levels—A Meta-Analysis. Int. J. Mol. Sci. 2020, 21, 3111. [Google Scholar] [CrossRef]
- Silva, B.; De Miranda, A.; Rodrigues, F.; Silveira, A.M.; Resende, G.D.S.; Moraes, M.F.D.; De Oliveira, A.P.; Parreiras, P.; Barcelos, L.S.; Teixeira, M.; et al. The 5-lipoxygenase (5-LOX) Inhibitor Zileuton Reduces Inflammation and Infarct Size with Improvement in Neurological Outcome Following Cerebral Ischemia. Curr. Neurovasc. Res. 2015, 12, 398–403. [Google Scholar] [CrossRef]
- Sahin, A.; Kam, W.R.; Darabad, R.R.; Topilow, K.; Sullivan, D.A. Regulation of Leukotriene B4Secretion by Human Corneal, Conjunctival, and Meibomian Gland Epithelial Cells. Arch. Ophthalmol. 2012, 130, 1013–1018. [Google Scholar] [CrossRef] [Green Version]
- Masoudi, S.; Zhao, Z.; Stapleton, F.; Willcox, M.D. Contact Lens–Induced Discomfort and Inflammatory Mediator Changes in Tears. Eye Contact Lens Sci. Clin. Pract. 2017, 43, 40–45. [Google Scholar] [CrossRef]
- Tager, A.M.; Bromley, S.K.; Medoff, B.D.; Islam, S.A.; Bercury, S.D.; Friedrich, E.B.; Carafone, A.D.; Gerszten, R.E.; Luster, A.D. Leukotriene B4 receptor BLT1 mediates early effector T cell recruitment. Nat. Immunol. 2003, 4, 982–990. [Google Scholar] [CrossRef]
- Goodarzi, K.; Goodarzi, M.; Tager, A.M.; Luster, A.D.; Von Andrian, U.H. Leukotriene B4 and BLT1 control cytotoxic effector T cell recruitment to inflamed tissues. Nat. Immunol. 2003, 4, 965–973. [Google Scholar] [CrossRef]
- Harauma, A.; Saito, J.; Watanabe, Y.; Moriguchi, T. Potential for daily supplementation of n-3 fatty acids to reverse symptoms of dry eye in mice. Prostaglandins Leukot. Essent. Fat. Acids 2014, 90, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Downie, L.E.; Ng, S.M.; Lindsley, K.B.; Akpek, E.K. Omega-3 and omega-6 polyunsaturated fatty acids for dry eye disease. Cochrane Database Syst. Rev. 2019, 12, CD011016. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.A.; Nai, G.; Giuffrida, R.; Sgrignoli, M.R.; Dos Santos, D.R.; Donadão, I.V.; Andrade, S.F.; DiNallo, H.R.; Andrade, S.F. Oral omega 3 in different proportions of EPA, DHA, and antioxidants as adjuvant in treatment of keratoconjunctivitis sicca in dogs. Arq. Bras. Oftalmol. 2018, 81, 421–428. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Treatment | Analysis | Time of Analysis |
|---|---|---|---|
| 1 | - | Histology | 0 h (control) |
| 2 | - | Histology | 6 h |
| 3 | - | Histology | 1 d 1 |
| 4 | - | Histology | 3 d |
| 5 | - | Histology | 7 d |
| 6 | - | Schirmer’s test | 0 h, 6 h, 1 d, 3 d, 7 d |
| 7 | - | Biochemistry 1 | 0 h, 6 h, 1 d, 3 d, 7 d |
| 8 | - | Lipidomics 2 | 0 h, 6 h, 1 d, 3 d, 7 d |
| 6 | DMSO | Histology | 1 d |
| 7 | DMSO | Histology | 3 d |
| 8 | DMSO | Histology | 7 d |
| 9 | DMSO/zileuton | Histology | 1 d |
| 10 | DMSO/zileuton | Histology | 3 d |
| 11 | DMSO/zileuton | Histology | 7 d |
| 12 | DMSO | Schirmer’s test | 1 d, 3 d, 7 d |
| 13 | DMSO | Biochemistry 1 | 1 d, 3 d, 7 d |
| 14 | DMSO | Lipidomics 2 | 1 d, 3 d, 7 d |
| 15 | DMSO/zileuton | Schirmer’s test | 1 d, 3 d, 7 d |
| 16 | DMSO/zileuton | Biochemistry 1 | 1 d, 3 d, 7 d |
| 17 | DMSO/zileuton | Lipidomics 2 | 1 d, 3 d, 7 d |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chistyakov, D.V.; Gancharova, O.S.; Baksheeva, V.E.; Tiulina, V.V.; Goriainov, S.V.; Azbukina, N.V.; Tsarkova, M.S.; Zamyatnin, A.A., Jr.; Philippov, P.P.; Sergeeva, M.G.; et al. Inflammation in Dry Eye Syndrome: Identification and Targeting of Oxylipin-Mediated Mechanisms. Biomedicines 2020, 8, 344. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8090344
Chistyakov DV, Gancharova OS, Baksheeva VE, Tiulina VV, Goriainov SV, Azbukina NV, Tsarkova MS, Zamyatnin AA Jr., Philippov PP, Sergeeva MG, et al. Inflammation in Dry Eye Syndrome: Identification and Targeting of Oxylipin-Mediated Mechanisms. Biomedicines. 2020; 8(9):344. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8090344
Chicago/Turabian StyleChistyakov, Dmitry V., Olga S. Gancharova, Viktoriia E. Baksheeva, Veronika V. Tiulina, Sergei V. Goriainov, Nadezhda V. Azbukina, Marina S. Tsarkova, Andrey A. Zamyatnin, Jr., Pavel P. Philippov, Marina G. Sergeeva, and et al. 2020. "Inflammation in Dry Eye Syndrome: Identification and Targeting of Oxylipin-Mediated Mechanisms" Biomedicines 8, no. 9: 344. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8090344