The Ubiquitin Ligase SIAH2 Negatively Regulates Glucocorticoid Receptor Activity and Abundance

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Experimental Animals
2.2. Serum ELISA and Liver TG Measurements
2.3. Gene Expression Analysis
2.4. Immunohistochemistry and Immunostaining
2.5. Preparation of Whole-Cell Extracts and Immunoblotting
2.6. Transient Transfections
2.7. Statistical Analysis
3. Results
3.1. Corticosterone-Mediated Changes in Body Mass and Body Composition in SIAH2+/+ and SIAH2−/− Mice
3.2. Genetic Deletion of SIAH2 Alters Endocrine Responses to Cort and Enhances Cort-Dependent Accumulation of Liver Triglycerides (TGs)
3.3. Genetic Deletion of SIAH2 Leads to Dysregulation of Genes Supporting Inflammation in eWAT within One Week
3.4. Inflammation-Associated Gene Expression Is Modulated by the Presence of SIAH2 during Three Weeks of Drug Regimen
3.5. Cort-Dependent Activation of Fibrotic Genes Occurs in the Absence of SIAH2
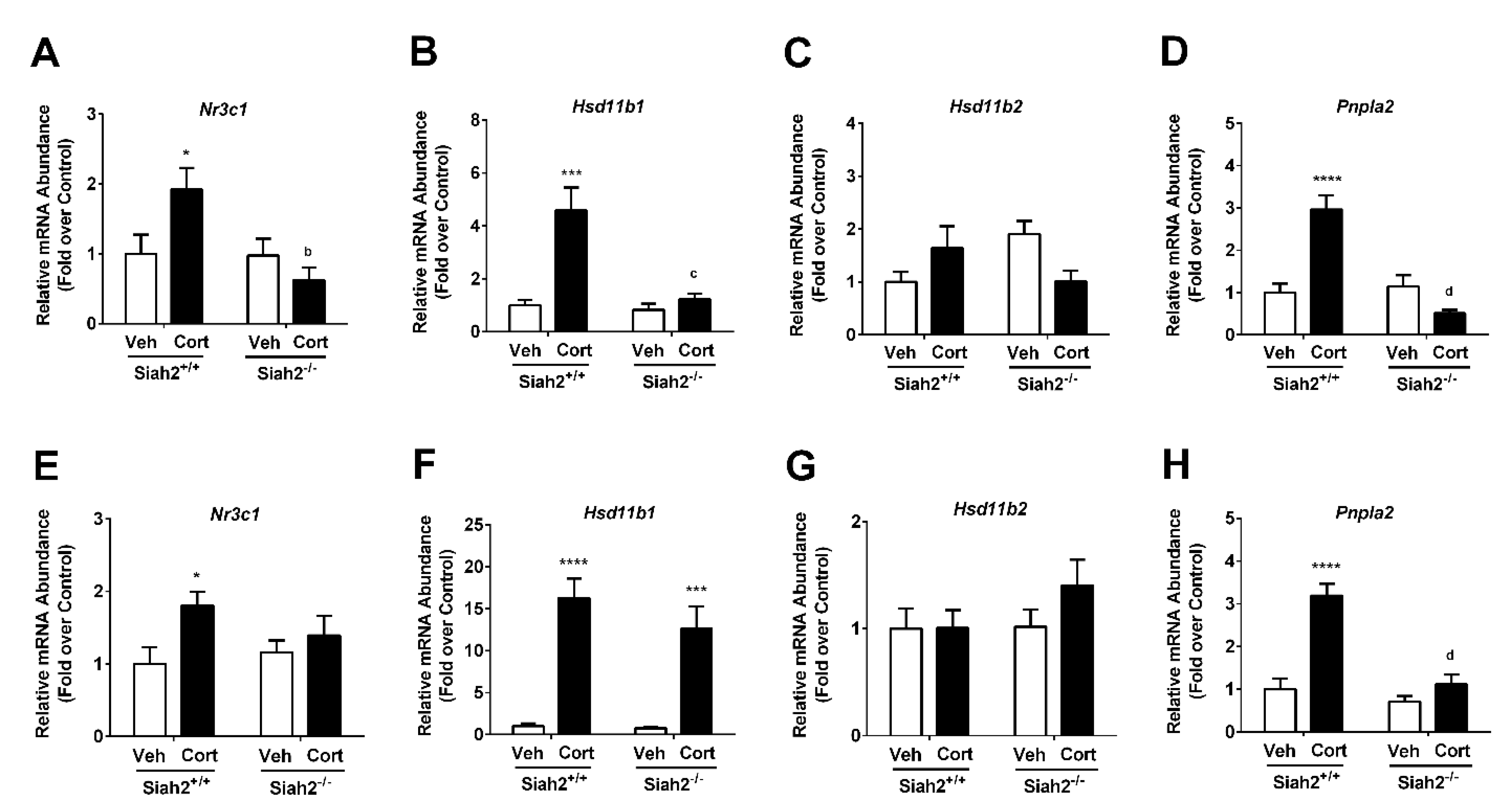
3.6. Glucocorticoid-Regulated Genes Are Influenced by Both Cort Exposure and the Presence of SIAH2
3.7. Glucocorticoid Receptor Abundance and Activity Are Modulated by SIAH2
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cain, D.W.; Cidlowski, J.A. Immune regulation by glucocorticoids. Nat. Rev. Immunol. 2017, 4, 233–247. [Google Scholar] [CrossRef] [PubMed]
- Kadmiel, M.; Cidlowski, J.A. Glucocorticoid receptor signaling in health and disease. Trends Pharmacol. Sci. 2013, 34, 518–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, I.R.; Wattie, D.J.; Evans, M.C.; Stapleton, J.P. Testosterone Therapy in Glucocorticoid-Treated Men. Arch. Intern. Med. 1996, 156, 1173–1177. [Google Scholar] [CrossRef] [PubMed]
- Burke, S.J.; Batdorf, H.M.; Huang, T.-Y.; Jackson, J.W.; Jones, K.A.; Martin, T.M.; Rohli, K.E.; Karlstad, M.D.; Sparer, T.E.; Burk, D.H.; et al. One week of continuous corticosterone exposure impairs hepatic metabolic flexibility, promotes islet β-cell proliferation, and reduces physical activity in male C57BL/6 J mice. J. Steroid Biochem. Mol. Biol. 2019, 195, 105468. [Google Scholar] [CrossRef] [PubMed]
- Burke, S.J.; Batdorf, H.M.; Eder, A.E.; Karlstad, M.D.; Burk, D.H.; Noland, R.C.; Floyd, Z.E.; Collier, J.J. Oral Corticosterone Administration Reduces Insulitis but Promotes Insulin Resistance and Hyperglycemia in Male Nonobese Diabetic Mice. Am. J. Pathol. 2017, 187, 614–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freedman, M.R.; Horwitz, B.A.; Stern, J.S. Effect of adrenalectomy and glucocorticoid replacement on development of obesity. Am. J. Physiol. Integr. Comp. Physiol. 1986, 250, R595–R607. [Google Scholar] [CrossRef]
- Peeke, P.M.; Chrousos, G.P. Hypercortisolism and Obesity. Ann. N. Y. Acad. Sci. 1995, 771, 665–676. [Google Scholar] [CrossRef]
- Schäcke, H.; Döcke, W.-D.; Asadullah, K. Mechanisms involved in the side effects of glucocorticoids. Pharmacol. Ther. 2002, 96, 23–43. [Google Scholar] [CrossRef]
- Do, T.T.H.; Marie, G.; Héloïse, D.; Guillaume, D.; Marthe, M.; Fève, B.; Marion, B. Glucocorticoid-induced insulin resistance is related to macrophage visceral adipose tissue infiltration. J. Steroid Biochem. Mol. Biol. 2019, 185, 150–162. [Google Scholar] [CrossRef]
- Kilroy, G.; Burk, D.H.; Floyd, Z.E. Siah2 Protein Mediates Early Events in Commitment to an Adipogenic Pathway. J. Biol. Chem. 2016, 291, 27289–27297. [Google Scholar] [CrossRef] [Green Version]
- Kilroy, G.; Carter, L.E.; Newman, S.; Burk, D.H.; Manuel, J.; Möller, A.; Bowtell, D.D.; Mynatt, R.L.; Ghosh, S.; Floyd, Z.E. The ubiquitin ligase Siah2 regulates obesity-induced adipose tissue inflammation. Obesity 2015, 23, 2223–2232. [Google Scholar] [CrossRef] [Green Version]
- Maneix, L.; Catic, A. Touch and go: Nuclear proteolysis in the regulation of metabolic genes and cancer. FEBS Lett. 2016, 590, 908–923. [Google Scholar] [CrossRef] [PubMed]
- Catic, A.; Suh, C.Y.; Hill, C.T.; Daheron, L.; Henkel, T.; Orford, K.W.; Dombkowski, D.M.; Liu, T.; Liu, X.S.; Scadden, D.T. Genome-wide Map of Nuclear Protein Degradation Shows NCoR1 Turnover as a Key to Mitochondrial Gene Regulation. Cell 2013, 155, 1380–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Guenther, M.G.; Carthew, R.W.; Lazar, M.A. Proteasomal regulation of nuclear receptor corepressor-mediated repression. Genes Dev. 1998, 12, 1775–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perissi, V.; Aggarwal, A.; Glass, C.K.; Rose, D.W.; Rosenfeld, M.G. A Corepressor/Coactivator Exchange Complex Required for Transcriptional Activation by Nuclear Receptors and Other Regulated Transcription Factors. Cell 2004, 116, 511–526. [Google Scholar] [CrossRef] [Green Version]
- Schulz, M.; Eggert, M.; Baniahmad, A.; Dostert, A.; Heinzel, T.; Renkawitz, R. RU486-induced Glucocorticoid Receptor Agonism Is Controlled by the Receptor N Terminus and by Corepressor Binding. J. Biol. Chem. 2002, 277, 26238–26243. [Google Scholar] [CrossRef] [Green Version]
- Frew, I.J.; Dickins, R.A.; Cuddihy, A.R.; Del Rosario, M.; Reinhard, C.; O’Connell, M.J.; Bowtell, D.D. Normal p53 Function in Primary Cells Deficient for Siah Genes. Mol. Cell. Biol. 2002, 22, 8155–8164. [Google Scholar] [CrossRef] [Green Version]
- Burke, S.J.; Batdorf, H.M.; Burk, D.H.; Noland, R.C.; Eder, A.E.; Boulos, M.S.; Karlstad, M.D.; Collier, J.J. db/db Mice Exhibit Features of Human Type 2 Diabetes That Are Not Present in Weight-Matched C57BL/6J Mice Fed a Western Diet. J. Diabetes Res. 2017, 2017, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, T.; Cepko, C. Electroporation and RNA interference in the rodent retina in vivo and in vitro. Proc. Natl. Acad. Sci. USA 2004, 101, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Spalding, K.L.; Arner, E.; Westermark, P.O.; Bernard, S.; Buchholz, B.A.; Bergmann, O.; Blomqvist, L.; Hoffstedt, J.; Näslund, E.; Britton, T.; et al. Dynamics of fat cell turnover in humans. Nat. Cell Biol. 2008, 453, 783–787. [Google Scholar] [CrossRef]
- Knittle, J.L.; Timmers, K.; Ginsberg-Fellner, F.; E Brown, R.; Katz, D.P. The growth of adipose tissue in children and adolescents. Cross-sectional and longitudinal studies of adipose cell number and size. J. Clin. Investig. 1979, 63, 239–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salans, L.B.; Knittle, J.L.; Hirsch, J. The role of adipose cell size and adipose tissue insulin sensitivity in the carbohydrate intolerance of human obesity. J. Clin. Investig. 1968, 47, 153–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sai, S.; Esteves, C.L.; Kelly, V.; Michailidou, Z.; Anderson, K.; Coll, A.P.; Nakagawa, Y.; Ohzeki, T.; Seckl, J.R.; Chapman, K.E. Glucocorticoid regulation of the promoter of 11beta-hydroxysteroid dehydrogenase type 1 is indirect and requires CCAAT/enhancer-binding protein-beta. Mol. Endocrinol. 2008, 9, 2049–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellingham, D.L.; Sar, M.; A Cidlowski, J. Ligand-dependent down-regulation of stably transfected human glucocorticoid receptors is associated with the loss of functional glucocorticoid responsiveness. Mol. Endocrinol. 1992, 6, 2090–2102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, P.-Z.; Zhu, Y.-M.; Zou, G.-H.; Sun, Y.-X.; Xiu, X.-L.; Huang, X.; Zhang, Q.-H. Relationship between Glucocorticoids and Insulin Resistance in Healthy Individuals. Med Sci. Monit. 2016, 22, 1887–1894. [Google Scholar] [CrossRef] [Green Version]
- Dalle, H.; Garcia, M.; Antoine, B.; Boehm, V.; Do, T.T.H.; Buyse, M.; Ledent, T.; Lamazière, A.; Magnan, C.; Postic, C.; et al. Adipocyte Glucocorticoid Receptor Deficiency Promotes Adipose Tissue Expandability and Improves the Metabolic Profile Under Corticosterone Exposure. Diabetes 2019, 68, 305–317. [Google Scholar] [CrossRef] [Green Version]
- Asterholm, I.W.; Tao, C.; Morley, T.S.; Wang, Q.A.; Delgado-Lopez, F.; Wang, Z.V.; Scherer, P.E. Adipocyte Inflammation Is Essential for Healthy Adipose Tissue Expansion and Remodeling. Cell Metab. 2014, 20, 103–118. [Google Scholar] [CrossRef] [Green Version]
- De Bosscher, K.; Vanden Berghe, W.; Haegeman, G. The interplay between the glucocorticoid receptor and nuclear factor-kappaB or activator protein-1: Molecular mechanisms for gene repression. Endocr. Rev. 2003, 4, 488–522. [Google Scholar] [CrossRef] [Green Version]
- Burke, S.J.; Goff, M.R.; Updegraff, B.L.; Lu, D.; Brown, P.L.; Minkin, S.C., Jr.; Biggerstaff, J.P.; Zhao, L.; Karlstad, M.D.; Collier, J.J. Regulation of the CCL2 Gene in Pancreatic beta-Cells by IL-1beta and Glucocorticoids: Role of MKP-1. PLoS ONE 2012, 10, e46986. [Google Scholar]
- Wallace, A.D.; Cidlowski, J.A. Proteasome-mediated Glucocorticoid Receptor Degradation Restricts Transcriptional Signaling by Glucocorticoids. J. Biol. Chem. 2001, 276, 42714–42721. [Google Scholar] [CrossRef] [Green Version]
- Freeman, B.C.; Yamamoto, K.R. Continuous recycling: A mechanism for modulatory signal transduction. Trends Biochem. Sci. 2001, 26, 285–290. [Google Scholar] [CrossRef]
- Unger, R.H. Lipid overload and overflow: Metabolic trauma and the metabolic syndrome. Trends Endocrinol. Metab. 2003, 14, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Unger, R.H. Lipotoxic Diseases. Annu. Rev. Med. 2002, 53, 319–336. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Thompson, C.F.; Balkovec, J.M.; Graham, D.W.; Hammond, M.L.; Quraishi, N.; Tata, J.R.; Einstein, M.; Ge, L.; Harris, G.; et al. NovelN-Arylpyrazolo[3,2-c]-Based Ligands for the Glucocorticoid Receptor: Receptor Binding and in Vivo Activity. J. Med. Chem. 2004, 47, 2441–2452. [Google Scholar] [CrossRef] [PubMed]
- Rosen, J.; Miner, J.N. The Search for Safer Glucocorticoid Receptor Ligands. Endocr. Rev. 2005, 26, 452–464. [Google Scholar] [CrossRef] [Green Version]
- Schäcke, H.; Berger, M.; Rehwinkel, H.; Asadullah, K. Selective glucocorticoid receptor agonists (SEGRAs): Novel ligands with an improved therapeutic index. Mol. Cell. Endocrinol. 2007, 275, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Weinstein, D.S.; Gong, H.; Doweyko, A.M.; Cunningham, M.; Habte, S.; Wang, J.H.; Holloway, D.A.; Burke, C.; Gao, L.; Guarino, V.; et al. Azaxanthene Based Selective Glucocorticoid Receptor Modulators: Design, Synthesis, and Pharmacological Evaluation of (S)-4-(5-(1-((1,3,4-Thiadiazol-2-yl)amino)-2-methyl-1-oxopropan-2-yl)-5H-chromeno[2,3-b]pyridin-2-yl)-2-fluoro-N,N-dimethylbenzamide (BMS-776532) and Its Methylene Homologue (BMS-791826). J. Med. Chem. 2011, 54, 7318–7333. [Google Scholar] [CrossRef]
- Burke, S.J.; May, A.L.; Noland, R.C.; Lu, D.; Brissova, M.; Powers, A.C.; Sherrill, E.M.; Karlstad, M.D.; Campagna, S.R.; Stephens, J.M. Thiobenzothiazole-modified Hydrocortisones Display Anti-inflammatory Activity with Reduced Impact on Islet beta-Cell Function. J. Biol. Chem. 2015, 21, 13401–13416. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burke, S.J.; Taylor, J.L.; Batdorf, H.M.; Noland, R.C.; Burk, D.H.; Yu, Y.; Floyd, Z.E.; Collier, J.J. The Ubiquitin Ligase SIAH2 Negatively Regulates Glucocorticoid Receptor Activity and Abundance. Biomedicines 2021, 9, 22. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9010022
Burke SJ, Taylor JL, Batdorf HM, Noland RC, Burk DH, Yu Y, Floyd ZE, Collier JJ. The Ubiquitin Ligase SIAH2 Negatively Regulates Glucocorticoid Receptor Activity and Abundance. Biomedicines. 2021; 9(1):22. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9010022
Chicago/Turabian StyleBurke, Susan J., Jessica L. Taylor, Heidi M. Batdorf, Robert C. Noland, David H. Burk, Yongmei Yu, Z. Elizabeth Floyd, and J. Jason Collier. 2021. "The Ubiquitin Ligase SIAH2 Negatively Regulates Glucocorticoid Receptor Activity and Abundance" Biomedicines 9, no. 1: 22. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9010022