
Profiles of Cytokines Secreted by ARPE-19 Cells Exposed to Light and Incubated with Anti-VEGF Antibody


Abstract
:1. Introduction
2. Methods
2.1. Cell Culture and Medium for Illuminated Culture
2.2. Cell Culture under Continuous Light Irradiation from Fluorescent Lamp
2.3. Cell Culture Incubated with Anti-VEGF Antibody
2.4. Cytokine Measurement
2.5. Statistical Analysis
3. Results
3.1. Inflammatory Cytokine Levels in ARPE-19 Cell Culture Supernatant
3.2. Correlation between Inflammatory Cytokine Levels
3.3. Hierarchical Cluster Analysis of Inflammatory Cytokine Expression Patterns
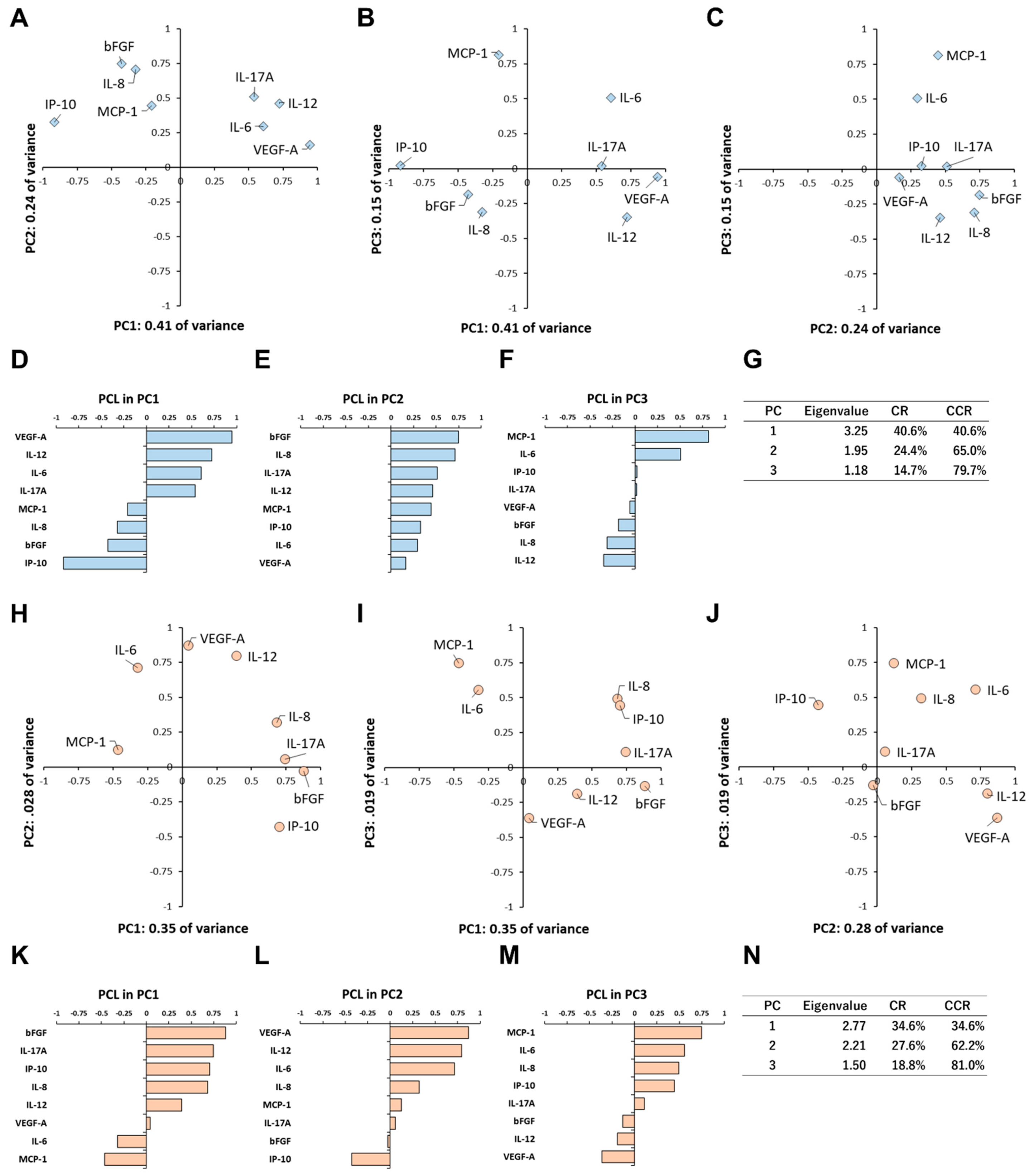
3.4. Principal Component Analysis of Inflammatory Cytokine Expression Patterns
3.5. Principal Component Analysis of Inflammatory Cytokine Expression Patterns under Light Irradiation
3.6. Cytokine Levels under Various Light Intensities
3.7. Cytokine Levels under Blue, Green or Red Light Irradiation
3.8. Levels of Inflammatory Cytokines with Anti-VEGF Antibody
3.9. Levels of Inflammatory Cytokines with Anti-VEGF Antibody under Light Irradiation
3.10. Cytokine Levels in Aflibercept- or Ranibizumab-exposed ARPE-19 Cells Cultured in Dark and under Light Irradiation
3.11. Cytokine Levels in Dark Culture Incubated with Aflibercept at Various Concentrations
3.12. Cytokine Levels in Light-Irradiated Culture Incubated with Aflibercept at Various Concentrations
3.13. Cytokine Levels in Dark Culture Incubated with Ranibizumab at Various Concentrations
3.14. Cytokine Levels under Light Irradiation Culture Incubated with Ranibizumab at Various Concentrations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Simo, R.; Villarroel, M.; Corraliza, L.; Hernandez, C.; Garcia-Ramirez, M. The retinal pigment epithelium: Something more than a constituent of the blood-retinal barrier—Implications for the pathogenesis of diabetic retinopathy. J. Biomed. Biotechnol. 2010, 2010, 190724. [Google Scholar] [CrossRef]
- Fields, M.A.; Del Priore, L.V.; Adelman, R.A.; Rizzolo, L.J. Interactions of the choroid, Bruch’s membrane, retinal pigment epithelium, and neurosensory retina collaborate to form the outer blood-retinal-barrier. Prog. Retin. Eye Res. 2020, 76, 100803. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, R.; Foss, A.; Leach, L. Establishment of a human in vitro model of the outer blood–retinal barrier. J. Anat. 2007, 211, 707–716. [Google Scholar] [CrossRef]
- Holtkamp, G.M.; Kijlstra, A.; Peek, R.; de Vos, A.F. Retinal pigment epithelium-immune system interactions: Cytokine production and cytokine-induced changes. Prog. Retin. Eye Res. 2001, 20, 29–48. [Google Scholar] [CrossRef]
- Mittal, R.; Bencie, N.; Parrish, J.M.; Liu, G.; Mittal, J.; Yan, D.; Liu, X.Z. An Update on Phosphodiesterase Mutations Underlying Genetic Etiology of Hearing Loss and Retinitis Pigmentosa. Front. Genet. 2018, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Keeling, E.; Lotery, A.J.; Tumbarello, D.A.; Ratnayaka, J.A. Impaired Cargo Clearance in the Retinal Pigment Epithelium (RPE) Underlies Irreversible Blinding Diseases. Cells 2018, 7, 16. [Google Scholar] [CrossRef] [Green Version]
- AnandBabu, K.; Sen, P.; Angayarkanni, N. Oxidized LDL, homocysteine, homocysteine thiolactone and advanced glycation end products act as pro-oxidant metabolites inducing cytokine release, macrophage infiltration and pro-angiogenic effect in ARPE-19 cells. PLoS ONE 2019, 14, e0216899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yam, M.; Engel, A.L.; Wang, Y.; Zhu, S.; Hauer, A.; Zhang, R.; Lohner, D.; Huang, J.; Dinterman, M.; Zhao, C.; et al. Proline mediates metabolic communication between retinal pigment epithelial cells and the retina. J. Biol. Chem. 2019, 294, 10278–10289. [Google Scholar] [CrossRef] [PubMed]
- Japanese Study Group of Polypoidal Choroidal Vasculopathy. Criteria for diagnosis of polypoidal choroidal vasculopathy. Nippon Ganka Gakkai Zasshi 2005, 109, 417–427. [Google Scholar]
- Grossniklaus, H.E.; Ling, J.X.; Wallace, T.M.; Dithmar, S.; Lawson, D.H.; Cohen, C.; Elner, V.M.; Elner, S.G.; Sternberg, P.J. Macrophage and retinal pigment epithelium expression of angiogenic cytokines in choroidal neovascularization. Mol. Vis. 2002, 8, 119–126. [Google Scholar]
- West, S.K.; Rosenthal, F.S.; Bressler, N.M.; Bressler, S.B.; Munoz, B.; Fine, S.L.; Taylor, H.R. Exposure to sunlight and other risk factors for age-related macular degeneration. Arch. Ophthalmol. 1989, 107, 875–879. [Google Scholar] [CrossRef] [PubMed]
- Blasiak, J.; Petrovski, G.; Veréb, Z.; Facskó, A.; Kaarniranta, K. Oxidative stress, hypoxia, and autophagy in the neovascular processes of age-related macular degeneration. BioMed Res. Int. 2014, 2014, 768026. [Google Scholar] [CrossRef]
- Wecker, T.; Ehlken, C.; Bühler, A.; Lange, C.; Agostini, H.; Böhringer, D.; Stahl, A. Five-year visual acuity outcomes and injection patterns in patients with pro-re-nata treatments for AMD, DME, RVO and myopic CNV. Br. J. Ophthalmol. 2017, 101, 353–359. [Google Scholar] [CrossRef] [Green Version]
- Goto, S.; Onishi, A.; Misaki, K.; Yonemura, S.; Sugita, S.; Ito, H.; Ohigashi, Y.; Ema, M.; Sakaguchi, H.; Nishida, K.; et al. Neural retina-specific Aldh1a1 controls dorsal choroidal vascular development via Sox9 expression in retinal pigment epithelial cells. eLife 2018, 7, e32358. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Nelson, K.C.; Wu, M.; Sternberg, P., Jr.; Jones, D.P. Oxidative damage and protection of the RPE. Prog. Retin. Eye Res. 2000, 19, 205–221. [Google Scholar] [CrossRef]
- Beatty, S.; Koh, H.H.; Phil, M.; Henson, D.; Boulton, M. The role of oxidative stress in the pathogenesis of age-related macular degeneration. Surv. Ophthalmol. 2000, 45, 115–134. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.; Tsujikawa, M.; Itabe, H.; Du, Z.-J.; Xie, P.; Matsumura, N.; Fu, X.; Zhang, R.; Sonoda, K.; Egashira, K.; et al. Chronic photo-oxidative stress and subsequent MCP-1 activation as causative factors for age-related macular degeneration. J. Cell Sci. 2012, 125, 2407–2415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruickshanks, K.J.; Klein, R.; Klein, B.E. Sunlight and age-related macular degeneration: The Beaver Dam Eye Study. Arch. Ophthalmol. 1993, 111, 514–518. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Perez, J.M.; Morillas-Ruiz, J.M. A review: Inflammatory process in Alzheimer’s disease, role of cytokines. Sci. World J. 2012, 2012, 756357. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Cohen, M.C.; Cohen, S. Cytokine function: A study in biologic diversity. Am. J. Clin. Pathol. 1996, 105, 589–598. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Ito, M.; Ishida, M.; Karasawa, Y. Phototoxicity of indocyanine green under continuous fluorescent lamp illumination and its prevention by blocking red light on cultured Muller cells. Investig. Ophthalmol. Vis. Sci. 2010, 51, 4337–4345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takayama, K.; Sato, T.; Karasawa, Y.; Sato, S.; Ito, M.; Takeuchi, M. Phototoxicity of indocyanine green and Brilliant Blue G under continuous fluorescent illumination on cultured human retinal pigment epithelial cells. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7389–7394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.; Karasawa, Y.; Ishikawa, S.; Taguchi, M.; Muraoka, T.; Ito, M.; Takeuchi, M. Potential Phototoxicity of Indocyanine Green in Retinal Pigment Epithelial Cells after Angiography under Ambient Illumination. Oxid. Med. Cell Longev. 2018, 2018, 6065285. [Google Scholar] [CrossRef]
- Sato, T.; Takeuchi, M.; Karasawa, Y.; Enoki, T.; Ito, M. Intraocular inflammatory cytokines in patients with neovascular age-related macular degeneration before and after initiation of intravitreal injection of anti-VEGF inhibitor. Sci. Rep. 2018, 8, 1098. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Takeuchi, M.; Karasawa, Y.; Takayama, K.; Enoki, T. Comprehensive expression patterns of inflammatory cytokines in aqueous humor of patients with neovascular age-related macular degeneration. Sci. Rep. 2019, 9, 19447. [Google Scholar] [CrossRef] [PubMed]
- Wilks, D.S. Cluster Analysis in International Geophysics; Elsevier: Amsterdam, The Netherlands, 2011; Volume 100, pp. 603–616. [Google Scholar]
- Jolliffe, I. Principal Component Analysis in International Encyclopedia of Statistical Science; Springer: Heidelberg, Germany, 2011; pp. 1094–1096. [Google Scholar]
- Dunn, K.C.; Aotaki-Keen, A.E.; Putkey, F.R.; Hjelmeland, L.M. ARPE-19, a human retinal pigment epithelial cell line with differentiated properties. Exp. Eye Res. 1996, 62, 155–169. [Google Scholar] [CrossRef]
- Samuel, W.; Jaworski, C.; Postnikova, O.A.; Kutty, R.K.; Duncan, T.; Tan, L.X.; Poliakov, E.; Lakkaraju, A.; Redmond, T.M. Appropriately differentiated ARPE-19 cells regain phenotype and gene expression profiles similar to those of native RPE cells. Mol. Vis. 2017, 23, 60–89. [Google Scholar]
- Bhisitkul, R. Vascular endothelial growth factor biology: Clinical implications for ocular treatments. Br. J. Ophthalmol. 2006, 90, 1542–1547. [Google Scholar] [CrossRef]
- Gospodarowicz, D.; Neufeld, G.; Schweigerer, L. Fibroblast growth factor: Structural and biological properties. J. Cell Physiol. Suppl. 1987, 133, 15–26. [Google Scholar] [CrossRef]
- Gospodarowicz, D.; Neufeld, G.; Schweigerer, L. Molecular and biological characterization of fibroblast growth factor, an angiogenic factor which also controls the proliferation and differentiation of mesoderm and neuroectoderm derived cells. Cell Differ. 1986, 19, 1–17. [Google Scholar] [CrossRef]
- Wenkel, H.; Streilein, J.W. Evidence that retinal pigment epithelium functions as an immune-privileged tissue. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3467–3473. [Google Scholar]
- Streilein, J.W.; Ma, N.; Wenkel, H.; Ng, T.F.; Zamiri, P. Immunobiology and privilege of neuronal retina and pigment epithelium transplants. Vis. Res. 2002, 42, 487–495. [Google Scholar] [CrossRef]
- Taylor, A.W. Ocular immune privilege. Eye 2009, 23, 1885–1889. [Google Scholar] [CrossRef] [Green Version]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef]
- Arras, M.; Ito, W.D.; Scholz, D.; Winkler, B.; Schaper, J.; Schaper, W. Monocyte activation in angiogenesis and collateral growth in the rabbit hindlimb. J. Clin. Investig. 1998, 101, 40–50. [Google Scholar] [CrossRef] [Green Version]
- Ito, W.D.; Arras, M.; Winkler, B.; Scholz, D.; Schaper, J.; Schaper, W. Monocyte chemotactic protein-1 increases collateral and peripheral conductance after femoral artery occlusion. Circ. Res. 1997, 80, 829–837. [Google Scholar] [CrossRef]
- Naldini, A.; Carraro, F. Role of inflammatory mediators in angiogenesis. Curr. Drug Targets Inflamm. Allergy 2005, 4, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Watford, W.T.; Moriguchi, M.; Morinobu, A.; O’Shea, J.J. The biology of IL-12: Coordinating innate and adaptive immune responses. Cytokine Growth Factor Rev. 2003, 14, 361–368. [Google Scholar] [CrossRef]
- Voest, E.E.; Kenyon, B.M.; O’Reilly, M.S.; Truitt, G.; D’Amato, R.J.; Folkman, J. Inhibition of angiogenesis in vivo by interleukin 12. J. Natl. Cancer Inst. 1995, 87, 581–586. [Google Scholar] [CrossRef]
- Shen, Y.; Xie, C.; Gu, Y.; Li, X.; Tong, J. Illumination from light-emitting diodes (LEDs) disrupts pathological cytokines expression and activates relevant signal pathways in primary human retinal pigment epithelial cells. Exp. Eye Res. 2016, 145, 456–467. [Google Scholar] [CrossRef] [PubMed]
- King, A.; Gottlieb, E.; Brooks, D.G.; Murphy, M.P.; Dunaief, J.L. Mitochondria-derived reactive oxygen species mediate blue light-induced death of retinal pigment epithelial cells. Photochem. Photobiol. 2004, 79, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Godley, B.F.; Shamsi, F.A.; Liang, F.Q.; Jarrett, S.G.; Davies, S.; Boulton, M. Blue light induces mitochondrial DNA damage and free radical production in epithelial cells. J. Biol. Chem. 2005, 280, 21061–21066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, J.P.; Lascaratos, G.; Bron, A.J.; Osborne, N.N. The influence of visible light exposure on cultured RGC-5 cells. Mol. Vis. 2007, 14, 334–344. [Google Scholar] [PubMed]
- Osborne, N.N.; Li, G.Y.; Ji, D.; Mortiboys, H.J.; Jackson, S. Light affects mitochondria to cause apoptosis to cultured cells: Possible relevance to ganglion cell death in certain optic neuropathies. J. Neurochem. 2008, 105, 2013–2028. [Google Scholar] [CrossRef] [PubMed]
- Knels, L.; Valtink, M.; Roehlecke, C.; Lupp, A.; de la Vega, J.; Mehner, M.; Funk, R.H.W. Blue light stress in retinal neuronal (R28) cells is dependent on wavelength range and irradiance. Eur. J. Neurosci. 2011, 34, 548–558. [Google Scholar] [CrossRef]
- Rutar, M.; Provis, J.M.; Valter, K. Brief exposure to damaging light causes focal recruitment of macrophages, and long-term destabilization of photoreceptors in the albino rat retina. Curr. Eye Res. 2010, 35, 631–643. [Google Scholar] [CrossRef]
- Karu, T.I.; Pyatibrat, L.V.; Afanasyeva, N.I. A novel mitochondrial signaling pathway activated by visible-to-near infrared radiation. Photochem. Photobiol. 2004, 80, 366–372. [Google Scholar] [CrossRef]
- Wong-Riley, M.T.; Liang, H.L.; Eells, J.T.; Chance, B.; Henry, M.M.; Buchmann, E.; Kane, M.; Whelan, H.T. Photobiomodulation directly benefits primary neurons functionally inactivated by toxins: Role of cytochrome c oxidase. J. Biol. Chem. 2005, 280, 4761–4771. [Google Scholar] [CrossRef] [Green Version]
- Begum, R.; Powner, M.; Hudson, N.; Hogg, C.; Jeffery, G. Treatment with 670 nm light up regulates cytochrome C oxidase expression and reduces inflammation in an age-related macular degeneration model. PLoS ONE 2013, 8, e57828. [Google Scholar] [CrossRef] [Green Version]
- Ying, R.; Liang, H.L.; Whelan, H.T.; Eells, J.T.; Wong-Riley, M.T. Pretreatment with near-infrared light via light-emitting diode provides added benefit against rotenone- and MPP+-induced neurotoxicity. Brain Res. 2008, 1243, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Núñez-Álvarez, C.; del Olmo-Aguado, S.; Merayo-Lloves, J.; Osborne, N.N. Near infra-red light attenuates corneal endothelial cell dysfunction in situ and in vitro. Exp. Eye Res. 2017, 161, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Saenz-de-Viteri, M.; Fernández-Robredo, P.; Hernández, M.; Bezunartea, J.; Reiter, N.; Recalde, S.; García-Layana, A. Single- and repeated-dose toxicity study of bevacizumab, ranibizumab, and aflibercept in ARPE-19 cells under normal and oxidative stress conditions. Biochem. Pharmacol. 2016, 103, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Ciombor, K.K.; Berlin, J.; Chan, E. Aflibercept. Clin. Cancer Res. 2013, 19, 1920–1925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blick, S.K.; Keating, G.M.; Wagstaff, A.J. Ranibizumab. Drugs 2007, 67, 1199–1206. [Google Scholar] [CrossRef]
- Brenner, M.; Hearing, V.J. The protective role of melanin against UV damage in human skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.N.; Simon, J.D.; Sarna, T. Role of ocular melanin in ophthalmic physiology and pathology. Photochem. Photobiol. 2008, 84, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Pilat, A.; Gerwat, W.; Skumatz, C.M.; Ito, M.; Kiyono, A.; Zadlo, A.; Nakanishi, Y.; Kolbe, L.; Burke, J.M.; et al. Photoaging of human retinal pigment epithelium is accompanied by oxidative modifications of its eumelanin. Pigment Cell Melanoma Res. 2013, 26, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Sarna, T. Properties and function of the ocular melanin--a photobiophysical view. J. Photochem. Photobiol. B 1992, 12, 215–258. [Google Scholar] [CrossRef]
- Rózanowski, B.; Burke, J.M.; Boulton, M.E.; Sarna, T.; Rózanowska, M. Human RPE melanosomes protect from photosensitized and iron-mediated oxidation but become pro-oxidant in the presence of iron upon photodegradation. Investig. Ophthalmol. Vis. Sci. 2008, 49, 2838–2847. [Google Scholar] [CrossRef] [Green Version]
- Burke, J.M.; Kaczara, P.; Skumatz, C.M.; Zareba, M.; Raciti, M.W.; Sarna, T. Dynamic analyses reveal cytoprotection by RPE melanosomes against non-photic stress. Mol. Vis. 2011, 17, 2864–2877. [Google Scholar]
- Simon, J.D.; Peles, D.; Wakamatsu, K.; Ito, S. Current challenges in understanding melanogenesis: Bridging chemistry, biological control, morphology, and function. Pigment Cell Melanoma Res. 2009, 22, 563–579. [Google Scholar] [CrossRef]
- Simon, J.D.; Peles, D.N. The red and the black. Acc. Chem. Res. 2010, 43, 1452–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, S.; Wakamatsu, K. Human hair melanins: What we have learned and have not learned from mouse coat color pigmentation. Pigment Cell Melanoma Res. 2011, 24, 63–74. [Google Scholar] [CrossRef]
- Marmor, M.F.; Wolfensberger, T. The retinal pigment epithelium. Funct. Dis. 1998, 103–134. [Google Scholar]
- Boulton, M.; Rozanowska, M.; Rozanowski, B. Retinal photodamage. J. Photochem. Photobiol. B 2001, 64, 144–161. [Google Scholar] [CrossRef]
- Seagle, B.L.; Rezai, K.A.; Kobori, Y.; Gasyna, E.M.; Rezaei, K.A.; Norris, J.R. Melanin photoprotection in the human retinal pigment epithelium and its correlation with light-induced cell apoptosis. Proc. Natl. Acad. Sci. USA 2005, 102, 8978–8983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blasiak, J.; Piechota, M.; Pawlowska, E.; Szatkowska, M.; Sikora, E.; Kaarniranta, K. Cellular senescence in age-related macular degeneration: Can autophagy and DNA damage response play a role? Oxid. Med. Cell Longev. 2017, 2017, 5293258. [Google Scholar] [CrossRef] [PubMed]
- Sikora, E. Rejuvenation of senescent cells-the road to postponing human aging and age-related disease? Exp. Gerontol. 2013, 48, 661–666. [Google Scholar] [CrossRef]
- Kozlowski, M.R. The ARPE-19 cell line: Mortality status and utility in macular degeneration research. Curr. Eye Res. 2015, 40, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Alster, O.; Bielak-Zmijewska, A.; Mosieniak, G.; Moreno-Villanueva, M.; Dudka-Ruszkowska, W.; Wojtala, A.; Kusio-Kobiałka, M.; Korwek, Z.; Burkle, A.; Piwocka, K.; et al. The role of nibrin in doxorubicin-induced apoptosis and cell senescence in Nijmegen Breakage Syndrome patients lymphocytes. PLoS ONE 2014, 13, e104964. [Google Scholar] [CrossRef] [Green Version]
- Pospelova, T.V.; Demidenko, Z.N.; Bukreeva, E.I.; Pospelov, V.A.; Gudkov, A.V.; Blagosklonny, M.V. Pseudo-DNA damage response in senescent cells. Cell Cycle 2009, 8, 4112–4118. [Google Scholar] [CrossRef] [Green Version]
- Arend, N.; Wertheimer, C.; Laubichler, P.; Wolf, A.; Kampik, A.; Kernt, M. Idebenone prevents oxidative stress, cell death and senescence of retinal pigment epithelium cells by stabilizing BAX/Bcl-2 ratio. Ophthalmologica 2015, 234, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Aryan, N.; Betts-Obregon, B.S.; Perry, G.; Tsin, A.T. Oxidative stress induces senescence in cultured RPE cells. Open Neurol. J. 2016, 10, 83–87. [Google Scholar] [CrossRef] [Green Version]
- Shimomachi, M.; Hasan, M.Z.; Kawaichi, M.; Oka, C. HtrA1 is induced by oxidative stress and enhances cell senescence through p38 MAPK pathway. Exp. Eye Res. 2013, 112, 79–92. [Google Scholar] [CrossRef]
- Zhuge, C.-C.; Xu, J.-Y.; Zhang, J.; Li, W.; Li, P.; Li, Z.; Chen, L.; Liu, X.; Shang, P.; Xu, H.; et al. Fullerenol protects retinal pigment epithelial cells from oxidative stress–induced premature senescence via activating SIRT1. Investig. Ophthalmol. Vis. Sci. 2014, 55, 4628–4638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spitzer, M.H.; Nolan, G.P. Mass cytometry: Single cells, many features. Cell 2016, 165, 780–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Dark | Light | p Value | Detection Range | ||||
|---|---|---|---|---|---|---|---|---|
| n | 20 | 18 | ||||||
| Detectable | Level | Detectable | Level | |||||
| Samples (%) | Mean ± SD (Median) | Samples (%) | Mean ± SD (Median) | |||||
| PDGF-BB | 0 (0) | 0 | 5 (27.8) | 0.69 ± 1.16 (0) | 5.83 × 10−3 | 0 | to | 26,155 |
| IL-1β | 0 (0) | 0 | 0 (0) | 0 | — | 0.33 | to | 6228 |
| IL-1ra | 16 (80.0) | 8.06 ± 4.69 (9.07) | 14 (77.8) | 10.5 ± 6.63 (11.5) | 0.100 | 4.97 | to | 132,198 |
| IL-2 | 2 (10.0) | 0.12 ± 0.36 (0) | 4 (22.2) | 0.30 ± 0.57 (0) | 0.128 | 0.54 | to | 18,791 |
| IL-4 | 1 (5.0) | 0.08 ± 0.34 (0) | 2 (11.1) | 0.15 ± 0.44 (0) | 0.279 | 0.80 | to | 4272 |
| IL-5 | 0 (0) | 0 | 0 (0) | 0 | — | 1.61 | to | 5381 |
| IL-6 | 20 (100) | 120.2 ± 63.3 (98.9) | 18 (100) | 70.0 ± 38.4 (56.6) | 0.003 | 1.39 | to | 21,699 |
| IL-7 | 20 (100) | 3.89 ± 0.42 (3.89) | 18 (100) | 3.39 ± 0.71 (3.32) | 0.005 | 0.49 | to | 11,208 |
| IL-8 | 20 (100) | 151.3 ± 39.9 (140.5) | 18 (100) | 112.1 ± 23.1 (110.3) | 4.18 × 10−4 | 1.38 | to | 25,457 |
| IL-9 | 19 (95.0) | 4.18 ± 1.64 (4.12) | 18 (100) | 8.17 ± 2.35 (8.89) | 2.31 × 10−75 | 1.35 | to | 9686 |
| IL-10 | 16 (80.0) | 2.47 ± 1.32 (2.88) | 17 (94.4) | 2.78 ± 0.82 (2.93) | 0.196 | 2.17 | to | 34,225 |
| IL-12 | 20 (100) | 47.2 ± 5.57 (47.0) | 18 (100) | 46.6 ± 3.59 (46.3) | 0.351 | 2.33 | to | 35,172 |
| IL-13 | 7 (35.0) | 0.84 ± 1.19 (0) | 2 (11.1) | 0.25 ± 0.72 (0) | 0.038 | 0.32 | to | 8051 |
| IL-15 | 20 (100) | 3.68 ± 1.27 (3.78) | 17 (94.4) | 3.95 ± 1.18 (4.14) | 0.250 | 1.50 | to | 6322 |
| IL-17A | 16 (80.0) | 11.3 ± 6.51 (13.8) | 17 (94.4) | 19.6 ± 10.0 (23.0) | 0.002 | 1.82 | to | 32,080 |
| Eotaxin | 3 (15.0) | 1.54 ± 3.86 (0) | 0 (0) | 0 | 0.050 | 0 | to | 25,602 |
| bFGF | 20 (100) | 30.7 ± 10.4 (29.6) | 18 (100) | 116.4 ± 31.5 (130.6) | 6.50 × 10−145 | 2.90 | to | 3719 |
| G-CSF | 19 (95.0) | 3.17 ± 1.05 (3.49) | 15 (83.3) | 3.49 ± 1.52 (3.92) | 0.227 | 1.67 | to | 8005 |
| GM-CSF | 0 (0) | 0 | 0 (0) | 0 | — | 2.62 | to | 13,998 |
| IFN-γ | 7 (35.0) | 3.56 ± 5.31 (0) | 4 (22.2) | 1.47 ± 2.85 (0) | 0.073 | 4.57 | to | 23,703 |
| IP-10 | 19 (95.0) | 24.4 ± 17.9 (18.3) | 15 (83.3) | 25.8 ± 17.9 (20.8) | 0.404 | 11.0 | to | 37,606 |
| MCP-1 | 20 (100) | 1652.5 ± 392.0 (1688.1) | 18 (100) | 1229.9 ± 396.3 (1208.7) | 0.001 | 1.52 | to | 19,447 |
| MIP-1α | 6 (30.0) | 0.39 ± 0.62 (0) | 2 (11.1) | 0.14 ± 0.40 (0) | 0.077 | 0.05 | to | 1083 |
| MIP-1β | 1 (5.0) | 0.05 ± 0.23 (0) | 1 (5.6) | 0.07 ± 0.30 (0) | 0.418 | 0.29 | to | 3679 |
| RANTES | 0 (0) | 0 | 1 (5.6) | 0.14 ± 0.59 (0) | 0.149 | 1.83 | to | 5365 |
| TNFα | 0 (0) | 0 | 0 (0) | 0 | — | 3.26 | to | 20,533 |
| VEGF-A | 20 (100) | 3963.8 ± 726.1 (4102.0) | 18 (100) | 3896.7 ± 553.1 (4043.5) | 0.376 | 2.42 | to | 38,721 |
| Category | Dark | p Value | ||||
|---|---|---|---|---|---|---|
| Anti-VEGF Antibody | None | Aflibercept | Ranibizumab | None | Aflibercept | |
| Level | Level | Level | vs. | vs. | ||
| Mean ± SD | Mean ± SD | Mean ± SD | Aflibercept | Ranibizumab | Ranibizumab | |
| PDGF-BB | 1.90 ± 0.54 | 1.51 ± 0 | 2.64 ± 2.31 | 0.211 | 0.351 | 0.281 |
| IL-1β | 0 | 0.66 ± 0.11 | 0.70 ± 0.06 | 0.007 | 0.002 | 0.349 |
| IL-1ra | 10.5 ± 0.62 | 12.2 ± 0.60 | 11.7 ± 0 | 0.053 | 0.049 | 0.211 |
| IL-2 | 0.51 ± 0.72 | 0.39 ± 0.55 | 0 | 0.434 | 0.211 | 0.211 |
| IL-4 | 0 | 0 | 0 | — | — | — |
| IL-5 | 0 | 0 | 0 | — | — | — |
| IL-6 | 4.08 ± 0.59 | 17.9 ± 1.20 | 17.0 ± 1.31 | 0.002 | 0.003 | 0.263 |
| IL-7 | 2.91 ± 0.56 | 2.20 ± 0.15 | 3.50 ± 0.28 | 0.112 | 0.156 | 0.014 |
| IL-8 | 28.7 ± 5.37 | 47.4 ± 6.80 | 52.8 ± 1.99 | 0.046 | 0.014 | 0.200 |
| IL-9 | 2.29 ± 0.09 | 3.25 ± 0 | 3.88 ± 0.70 | 0.002 | 0.043 | 0.167 |
| IL-10 | 0 | 0 | 0 | — | — | — |
| IL-12 | 11.9 ± 1.32 | 0 | 3.68 ± 0.30 | 0.003 | 0.007 | 0.002 |
| IL-13 | 0.94 ± 0.03 | 0.41 ± 0 | 0.54 ± 0.06 | 0.001 | 0.008 | 0.054 |
| IL-15 | 2.53 ± 0.02 | 5.51 ± 0.46 | 5.22 ± 0.06 | 0.006 | 1.26 × 10−4 | 0.238 |
| IL-17A | 7.46 ± 0.62 | 10.3 ± 1.83 | 10.9 ± 1.22 | 0.088 | 0.035 | 0.359 |
| Eotaxin | 0 | 0 | 0 | — | — | — |
| bFGF | 139.0 ± 2.69 | 301.3 ± 29.8 | 319.3 ± 8.35 | 0.008 | 0.001 | 0.249 |
| G-CSF | 0 | 0 | 0 | — | — | — |
| GM-CSF | 0 | 0 | 0 | — | — | — |
| IFN-γ | 0 | 0 | 0 | — | — | — |
| IP-10 | 0 | 0 | 0 | — | — | — |
| MCP-1 | 97.7 ± 1.00 | 231.1 ± 11.8 | 211.7 ± 11.0 | 0.002 | 0.002 | 0.116 |
| MIP-1α | 0 | 0.13 ± 0.18 | 0 | 0.211 | — | 0.211 |
| MIP-1β | 0 | 0 | 0 | — | — | — |
| RANTES | 0 | 0 | 0 | — | — | — |
| TNFα | 0 | 0 | 0 | — | — | — |
| VEGF-A | 243.0 ± 2.51 | 3.24 ± 0.28 | 115.2 ± 3.04 | 2.77 × 10−55 | 2.38 × 10−4 | 1.86 × 10−4 |
| Category | Light | p value | ||||
|---|---|---|---|---|---|---|
| Anti-VEGF Antibody | None | Aflibercept | Ranibizumab | None | Aflibercept | |
| Level | Level | Level | vs. | vs. | ||
| Mean ± SD | Mean ± SD | Mean ± SD | Aflibercept | Ranibizumab | Ranibizumab | |
| PDGF-BB | 2.11 ± 0.73 | 2.02 ± 0 | 1.01 ± 1.43 | 0.442 | 0.162 | 0.211 |
| IL-1β | 0 | 0 | 0 | — | — | — |
| IL-1ra | 11.2 ± 1.96 | 15.9 ± 1.15 | 14.0 ± 2.04 | 0.029 | 0.106 | 0.190 |
| IL-2 | 0.75 ± 0.13 | 1.34 ± 0.17 | 0.90 ± 0.38 | 0.011 | 0.280 | 0.137 |
| IL-4 | 0 | 0 | 0 | — | — | — |
| IL-5 | 0 | 0 | 0 | — | — | — |
| IL-6 | 2.88 ± 0.09 | 4.16 ± 0.47 | 3.53 ± 0.12 | 0.008 | 0.003 | 0.102 |
| IL-7 | 3.61 ± 1.13 | 0.74 ± 0 | 1.66 ± 0.62 | 0.021 | 0.060 | 0.085 |
| IL-8 | 19.2 ± 4.30 | 24.9 ± 0.88 | 26.1 ± 5.90 | 0.087 | 0.109 | 0.401 |
| IL-9 | 2.00 ± 0.08 | 1.66 ± 0.43 | 0.71 ± 1.00 | 0.117 | 0.047 | 0.173 |
| IL-10 | 0 | 0 | 0 | — | — | — |
| IL-12 | 12.4 ± 0.55 | 0 | 0 | 4.02 × 10−55 | 4.02 × 10−55 | — |
| IL-13 | 1.18 ± 0.11 | 0.21 ± 0.29 | 0.21 ± 0.29 | 0.006 | 0.006 | 0.550 |
| IL-15 | 3.41 ± 0.23 | 4.06 ± 0.45 | 3.74 ± 0.18 | 0.058 | 0.098 | 0.226 |
| IL-17A | 6.23 ± 0.87 | 7.67 ± 0.31 | 5.73 ± 1.53 | 0.061 | 0.330 | 0.110 |
| Eotaxin | 0 | 0 | 0 | — | — | — |
| bFGF | 99.2 ± 16.3 | 127.0 ± 22.5 | 120.4 ± 29.3 | 0.100 | 0.180 | 0.412 |
| G-CSF | 0 | 0 | 0 | — | — | — |
| GM-CSF | 0 | 0 | 0 | — | — | — |
| IFN-γ | 0 | 0 | 0 | — | — | — |
| IP-10 | 0 | 0 | 0 | — | — | — |
| MCP-1 | 48.8 ± 0.99 | 55.6 ± 2.19 | 57.7 ± 0.26 | 0.008 | 0.001 | 0.154 |
| MIP-1α | 0.26 ± 0 | 0.13 ± 0.18 | 0.34 ± 0.11 | 0.136 | 0.136 | 0.151 |
| MIP-1β | 0.19 ± 0.17 | 0 | 0 | 0.110 | 0.110 | — |
| RANTES | 0 | 0 | 0 | — | — | — |
| TNFα | 0 | 0 | 0 | — | — | — |
| VEGF-A | 242.2 ± 20.1 | 3.15 ± 0.42 | 47.4 ± 12.4 | 2.67 × 10−4 | 0.001 | 0.019 |
| Anti-VEGF Antibody | Aflibercept | Ranibizumab | ||||
|---|---|---|---|---|---|---|
| Category | Dark | Light | p Value | Dark | Light | p Value |
| Level | Level | Dark vs. Light | Level | Level | Dark vs. Light | |
| Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD | |||
| PDGF-BB | 1.51 ± 0 | 2.02 ± 0 | 1.000 | 2.64 ± 2.31 | 1.01 ± 1.43 | 0.243 |
| IL-1β | 0.66 ± 0.11 | 0 | 0.007 | 0.70 ± 0.06 | 0 | 0.002 |
| IL-1ra | 12.2 ± 0.60 | 15.9 ± 1.15 | 0.028 | 11.7 ± 0 | 14.0 ± 2.04 | 0.126 |
| IL-2 | 0.39 ± 0.55 | 1.34 ± 0.17 | 0.073 | 0 | 0.90 ± 0.38 | 0.040 |
| IL-4 | 0 | 0 | — | 0 | 0 | — |
| IL-5 | 0 | 0 | — | 0 | 0 | — |
| IL-6 | 17.9 ± 1.20 | 4.16 ± 0.47 | 0.002 | 17.0 ± 1.31 | 3.53 ± 0.12 | 0.002 |
| IL-7 | 2.20 ± 0.15 | 0.74 ± 0 | 0.003 | 3.50 ± 0.28 | 1.66 ± 0.62 | 0.031 |
| IL-8 | 47.4 ± 6.80 | 24.9 ± 0.88 | 0.022 | 52.8 ± 1.99 | 26.1 ± 5.90 | 0.013 |
| IL-9 | 3.25 ± 0 | 1.66 ± 0.43 | 0.017 | 3.88 ± 0.70 | 0.71 ± 1.00 | 0.034 |
| IL-10 | 0 | 0 | — | 0 | 0 | — |
| IL-12 | 0 | 0 | — | 3.68 ± 0.30 | 0 | 0.002 |
| IL-13 | 0.41 ± 0 | 0.21 ± 0.29 | 0.211 | 0.54 ± 0.06 | 0.21 ± 0.29 | 0.128 |
| IL-15 | 5.51 ± 0.46 | 4.06 ± 0.45 | 0.043 | 5.22 ± 0.06 | 3.74 ± 0.18 | 0.004 |
| IL-17A | 10.3 ± 1.83 | 7.67 ± 0.31 | 0.094 | 10.9 ± 1.22 | 5.73 ± 1.53 | 0.032 |
| Eotaxin | 0 | 0 | — | 0 | 0 | — |
| bFGF | 301.3 ± 29.8 | 127.0 ± 22.5 | 0.011 | 319.3 ± 8.35 | 120.4 ± 29.3 | 0.006 |
| G-CSF | 0 | 0 | — | 0 | 0 | — |
| GM-CSF | 0 | 0 | — | 0 | 0 | — |
| IFN-γ | 0 | 0 | — | 0 | 0 | — |
| IP-10 | 0 | 0 | — | 0 | 0 | — |
| MCP-1 | 231.1 ± 11.8 | 55.6 ± 2.19 | 0.001 | 211.7 ± 11.0 | 57.7 ± 0.26 | 0.001 |
| MIP-1α | 0.13 ± 0.18 | 0.13 ± 0.18 | 0.550 | 0 | 0.34 ± 0.11 | 0.026 |
| MIP-1β | 0 | 0 | — | 0 | 0 | — |
| RANTES | 0 | 0 | — | 0 | 0 | — |
| TNFα | 0 | 0 | — | 0 | 0 | — |
| VEGF-A | 3.24 ± 0.28 | 3.15 ± 0.42 | 0.407 | 115.2 ± 3.04 | 47.4 ± 12.4 | 0.009 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato, T.; Takeuchi, M.; Karasawa, Y.; Ito, M. Profiles of Cytokines Secreted by ARPE-19 Cells Exposed to Light and Incubated with Anti-VEGF Antibody. Biomedicines 2021, 9, 1333. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9101333
Sato T, Takeuchi M, Karasawa Y, Ito M. Profiles of Cytokines Secreted by ARPE-19 Cells Exposed to Light and Incubated with Anti-VEGF Antibody. Biomedicines. 2021; 9(10):1333. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9101333
Chicago/Turabian StyleSato, Tomohito, Masaru Takeuchi, Yoko Karasawa, and Masataka Ito. 2021. "Profiles of Cytokines Secreted by ARPE-19 Cells Exposed to Light and Incubated with Anti-VEGF Antibody" Biomedicines 9, no. 10: 1333. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9101333