
OM-85 Broncho-Vaxom®, a Bacterial Lysate, Reduces SARS-CoV-2 Binding Proteins on Human Bronchial Epithelial Cells


Abstract
:1. Introduction
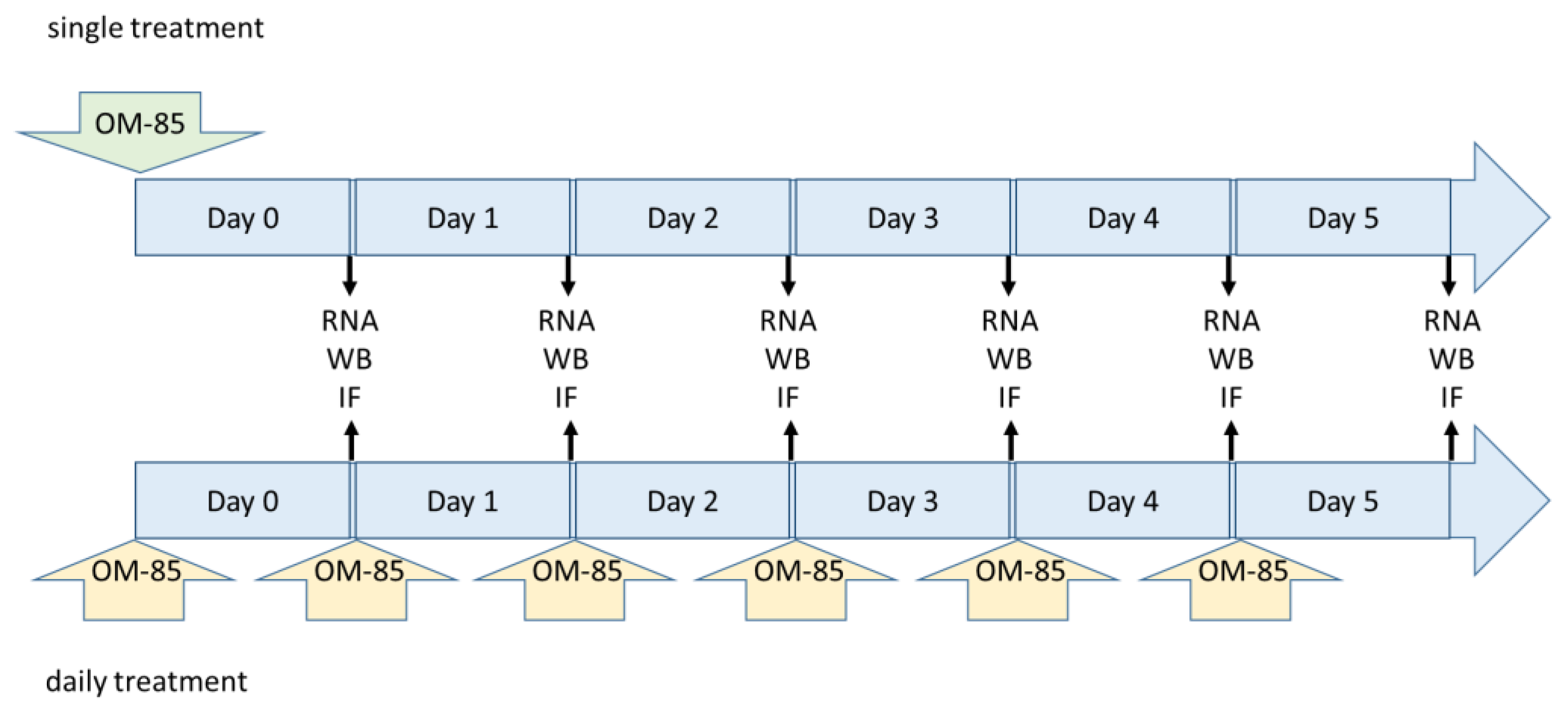
2. Materials and Methods
2.1. Cell Lines
2.2. Protein Analysis
2.3. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.4. Soluble ACE2 (sACE2) and Glycosaminoglycans
2.5. Hyaluronic Acid, HA-Sythanse, Hyaluronidases, Heparan Sulfate, and Heparanase
2.6. Epithelial Cells Infection with S-Protein Pseudo-Type Lentivirus
2.7. Statistics
3. Results
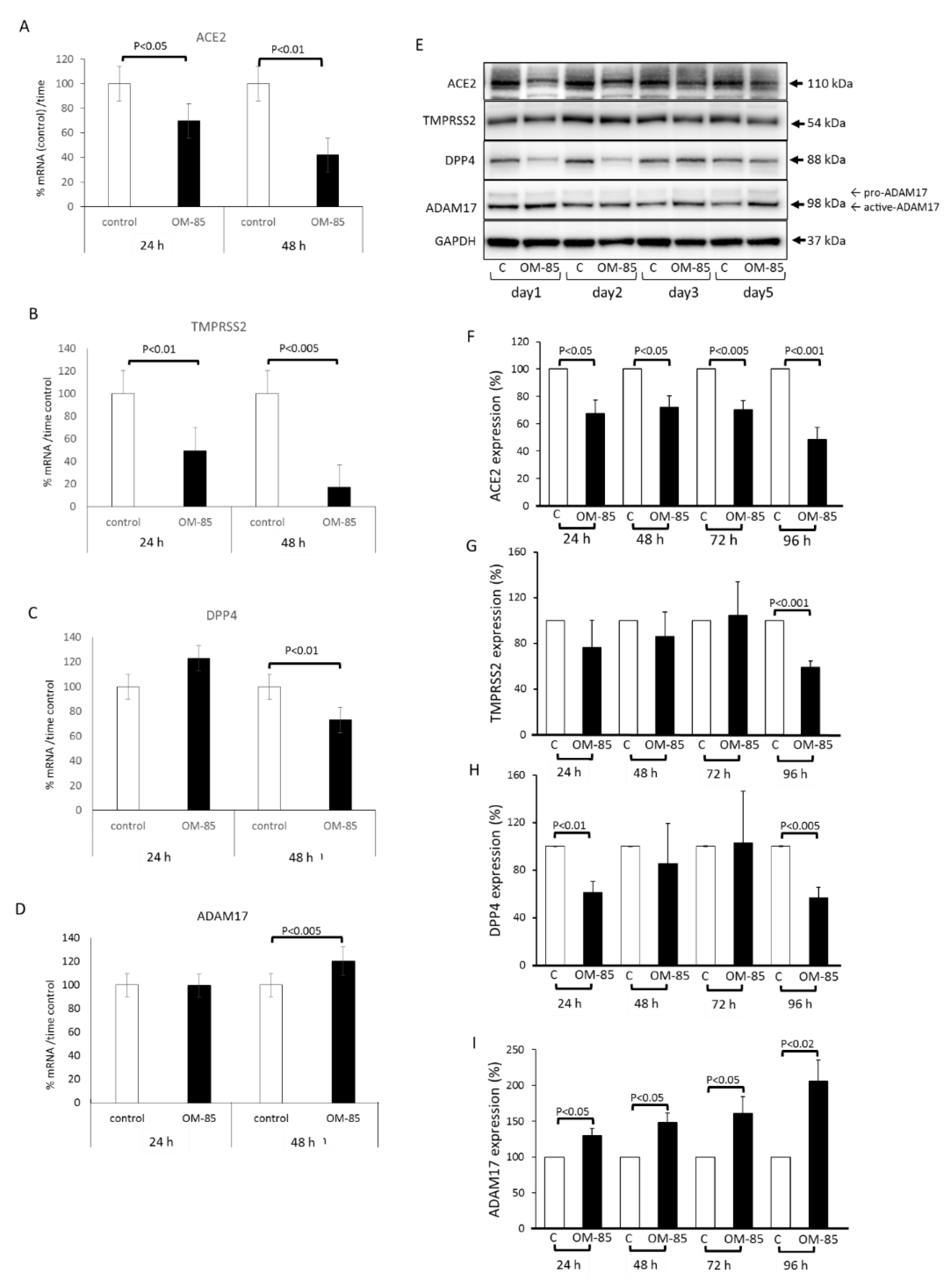
3.1. OM-85 Regulates SARS-CoV-2 Interacting Proteins on the Protein Level
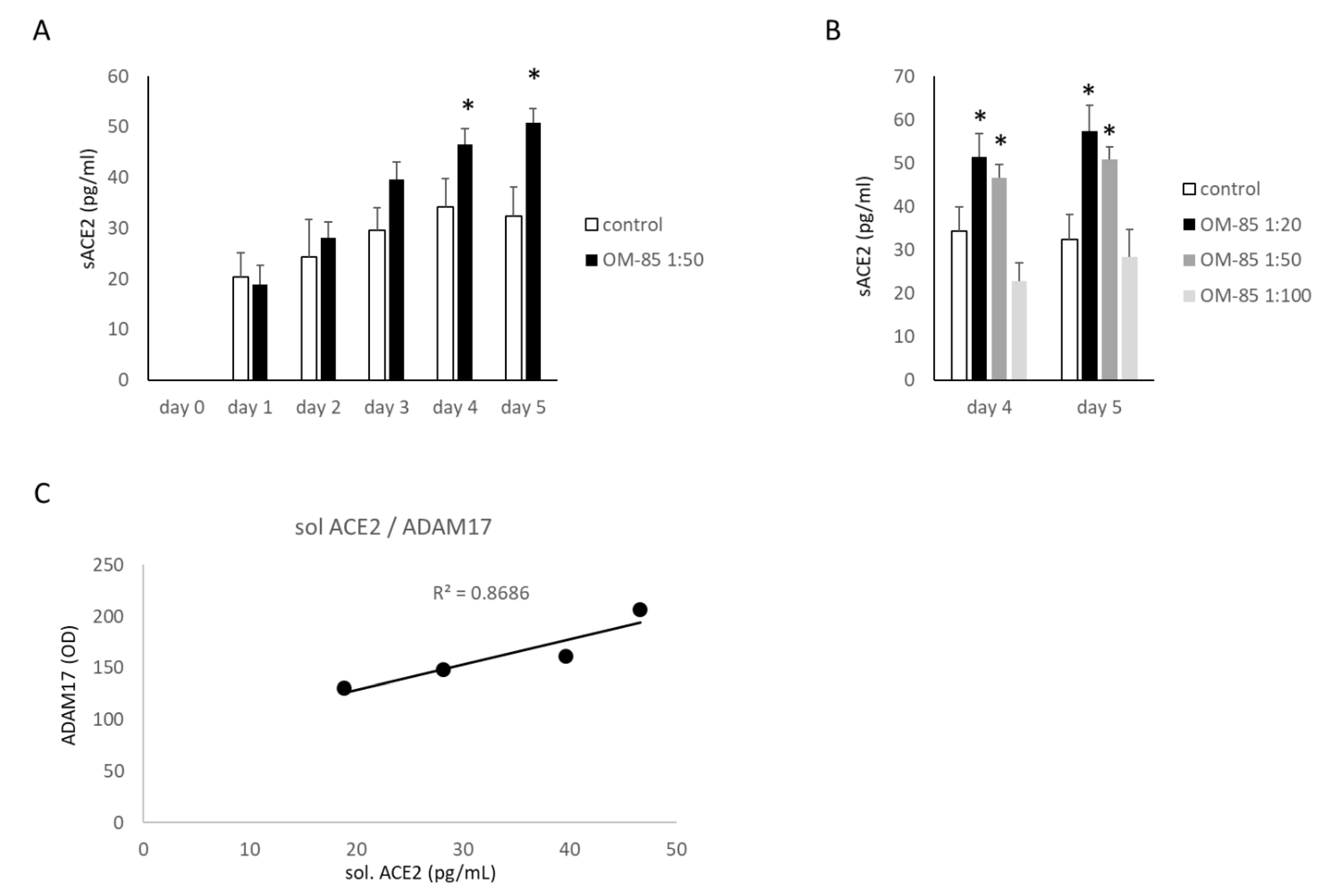
3.2. The Generation of Soluble ACE2 (sACE2)
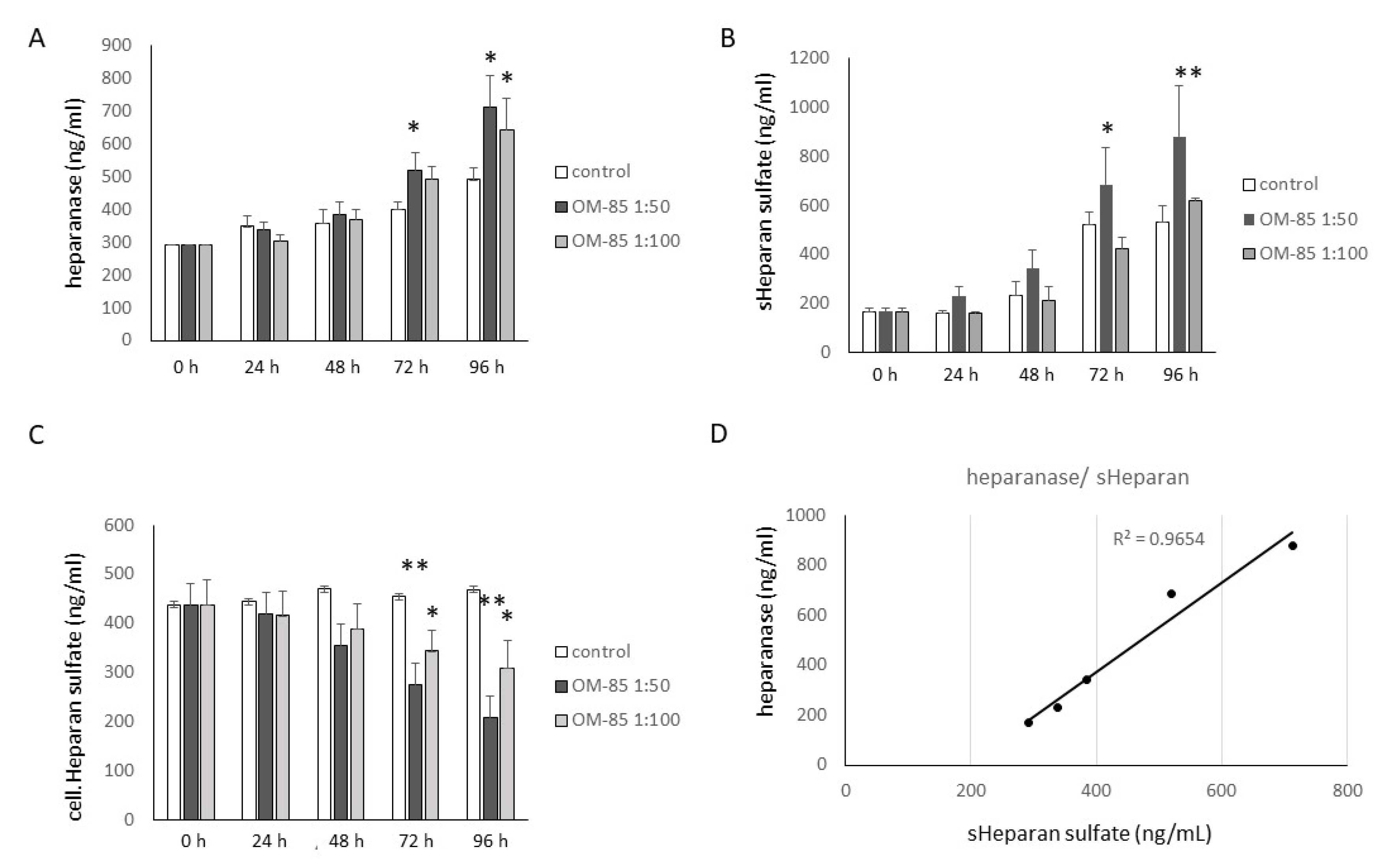
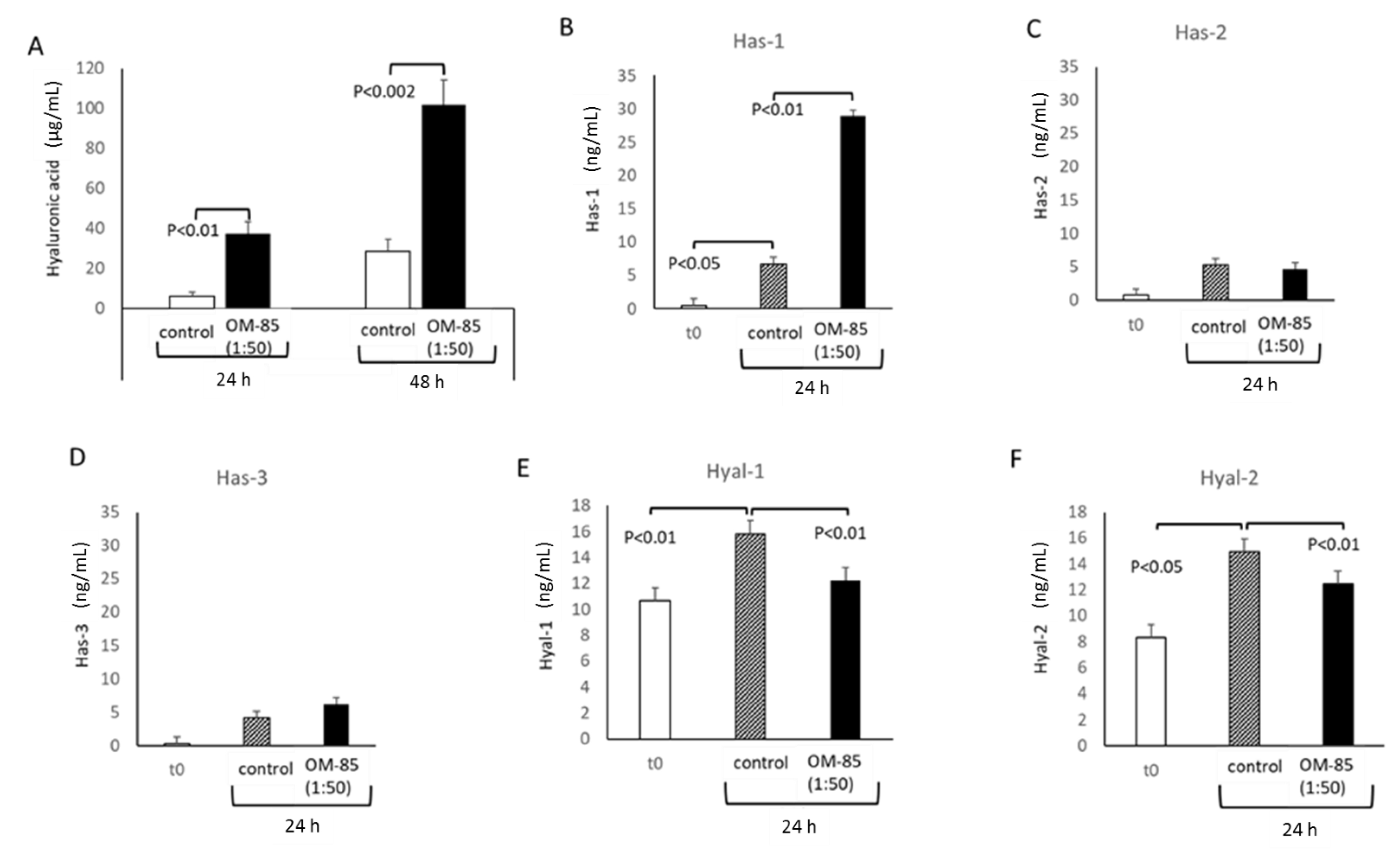
3.3. OM-85 Reduced HS by Increasing Heparanase Expression, but Increased HA Secretion
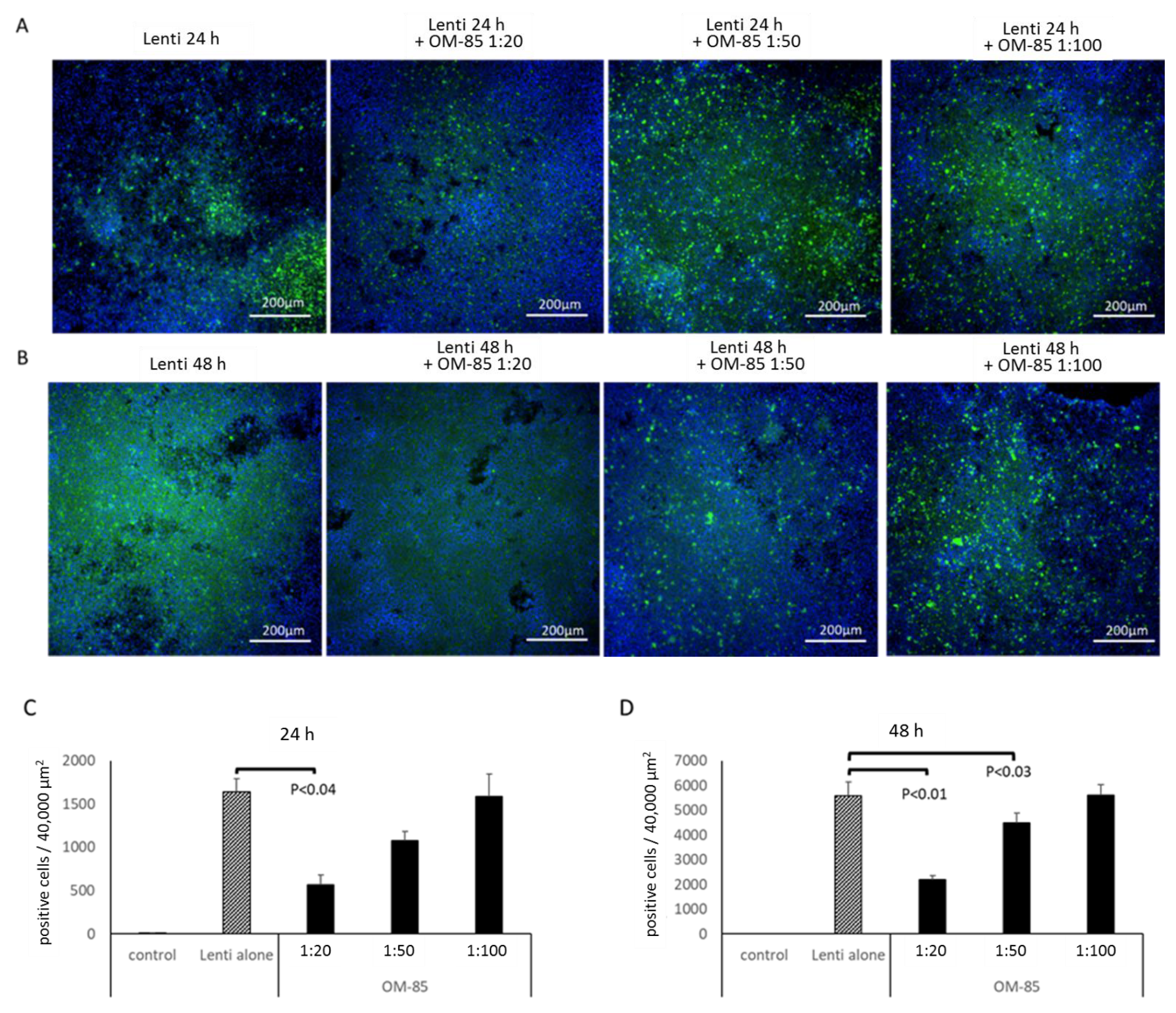
3.4. OM-85 Pretreatment Reduces Infection of Epithelial Cells with Pseudo-Typed S-Protein Lentivirus
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lukassen, S.; Chua, R.L.; Trefzer, T.; Kahn, N.C.; Schneider, M.A.; Muley, T.; Winter, H.; Meister, M.; Veith, C.; Boots, A.W.; et al. SARS-CoV-2 receptor ACE2 and TMPRSS2 are primarily expressed in bronchial transient secretory cells. EMBO J. 2020, 39, e105114. [Google Scholar] [CrossRef]
- Ruaro, B.; Salton, F.; Braga, L.; Wade, B.; Confalonieri, P.; Volpe, M.C.; Baratella, E.; Maiocchi, S.; Confalonieri, M. The History and Mystery of Alveolar Epithelial Type II Cells: Focus on Their Physiologic and Pathologic Role in Lung. Int. J. Mol. Sci. 2021, 22, 2566. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Jiang, S.C.; Zhang, Z.W.; Fu, Y.F.; Hu, J.; Li, Z.L. The Role of Alveolar Edema in COVID-19. Cells 2021, 10, 1897. [Google Scholar] [CrossRef]
- Subbarayan, K.; Ulagappan, K.; Wickenhauser, C.; Bachmann, M.; Seliger, B. Immune Interaction Map of Human SARS-CoV-2 Target Genes: Implications for Therapeutic Avenues. Front. Immunol. 2021, 12, 597399. [Google Scholar] [CrossRef]
- Raghav, P.K.; Kalyanaraman, K.; Kumar, D. Human cell receptors: Potential drug targets to combat COVID-19. Amino Acids 2021, 53, 813–842. [Google Scholar] [CrossRef]
- Zhang, C.; Verma, A.; Feng, Y.; Melo, M.C.R.; McQuillan, M.; Hansen, M.; Lucas, A.; Park, J.; Ranciaro, A.; Thompson, S.; et al. Global patterns of genetic variation and association with clinical phenotypes at genes involved in SARS-CoV-2 infection. medRxiv 2021. [Google Scholar] [CrossRef]
- Zoufaly, A.; Poglitsch, M.; Aberle, J.H.; Hoepler, W.; Seitz, T.; Traugott, M.; Grieb, A.; Pawelka, E.; Laferl, H.; Wenisch, C.; et al. Human recombinant soluble ACE2 in severe COVID-19. Lancet Respir. Med. 2020, 8, 1154–1158. [Google Scholar] [CrossRef]
- Mukerjee, S.; Gao, H.; Xu, J.; Sato, R.; Zsombok, A.; Lazartigues, E. ACE2 and ADAM17 Interaction Regulates the Activity of Presympathetic Neurons. Hypertension 2019, 74, 1181–1191. [Google Scholar] [CrossRef]
- Stolarczyk, M.; Veit, G.; Schnúr, A.; Veltman, M.; Lukacs, G.L.; Scholte, B.J. Extracellular oxidation in cystic fibrosis airway epithelium causes enhanced EGFR/ADAM17 activity. Am. J. Physiol. Lung Cell. Mol. Physiol. 2018, 314, L555–L568. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, K.S.; Crue, T.; Nall, J.M.; Foster, D.; Sajuthi, S.; Correll, K.A.; Nakamura, M.; Everman, J.; Downey, G.P.; Seibold, M.A.; et al. Influenza virus infection increases ACE2 expression and shedding in human small airway epithelial cells. Eur. Respir. J. 2021, 58, 2003988. [Google Scholar] [CrossRef] [PubMed]
- Hristova, M.; Stanilova, S.; Miteva, L. Serum concentration of renin-angiotensin system components in association with ACE I/D polymorphism among hypertensive subjects in response to ACE inhibitor therapy. Clin. Exp. Hypertens. 2019, 41, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Dart, A.B.; Wicklow, B.; Scholey, J.; Sellers, E.A.; Dyck, J.; Mahmud, F.; Sochett, E.; Hamilton, J.; Blydt-Hansen, T.; Burns, K. An evaluation of renin-angiotensin system markers in youth with type 2 diabetes and associations with renal outcomes. Pediatr. Diabetes 2020, 21, 1102–1109. [Google Scholar] [CrossRef]
- Hamet, P.; Pausova, Z.; Attaoua, R.; Hishmih, C.; Haloui, M.; Shin, J.; Paus, T.; Abrhamowicz, M.; Gaudet, D.; Santucci, L.; et al. SARS-COV-2 RECEPTOR ACE2 GENE is associated with hypertension and SEVERity of COVID 19: Interaction with sex, obesity and SMOKING. Am. J. Hypertens. 2021, 34, 367–376. [Google Scholar] [CrossRef]
- Emilsson, V.; Gudmundsson, E.F.; Aspelund, T.; Jonsson, B.G.; Gudjonsson, A.; Launer, L.J.; Lamb, J.R.; Gudmundsdottir, V.; Jennings, L.L.; Gudnason, V. ACE2 levels are altered in comorbidities linked to severe outcome in COVID-19. medRxiv 2020. [Google Scholar] [CrossRef]
- Rahman, M.M.; Hasan, M.; Ahmed, A. Potential detrimental role of soluble ACE2 in severe COVID-19 comorbid patients. Rev. Med. Virol. 2021, 31, 1–12. [Google Scholar] [CrossRef]
- Yalcin, H.C.; Sukumaran, V.; Al-Ruweidi, M.K.A.A.; Shurbaji, S. Do Changes in ACE-2 Expression Affect SARS-CoV-2 Virulence and Related Complications: A Closer Look into Membrane-Bound and Soluble Forms. Int. J. Mol. Sci. 2021, 22, 6703. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Pöhlmann, S. A Multibasic Cleavage Site in the Spike Protein of SARS-CoV-2 Is Essential for Infection of Human Lung Cells. Mol. Cell 2020, 78, 779–784.e5. [Google Scholar] [CrossRef]
- McKee, D.L.; Sternberg, A.; Stange, U.; Laufer, S.; Naujokat, C. Candidate drugs against SARS-CoV-2 and COVID-19. Pharmacol. Res. 2020, 157, 104859. [Google Scholar] [CrossRef]
- Pons, S.; Fodil, S.; Azoulay, E.; Zafrani, L. The vascular endothelium: The cornerstone of organ dysfunction in severe SARS-CoV-2 infection. Crit. Care 2020, 24, 353. [Google Scholar] [CrossRef]
- Radzikowska, U.; Ding, M.; Tan, G.; Zhakparov, D.; Peng, Y.; Wawrzyniak, P.; Wang, M.; Li, S.; Morita, H.; Altunbulakli, C.; et al. Distribution of ACE2, CD147, CD26, and other SARS-CoV-2 associated molecules in tissues and immune cells in health and in asthma, COPD, obesity, hypertension, and COVID-19 risk factors. Allergy 2020, 75, 2829–2845. [Google Scholar] [CrossRef] [PubMed]
- Cameron, K.; Rozano, L.; Falasca, M.; Mancera, R.L. Does the SARS-CoV-2 Spike Protein Receptor Binding Domain Interact Effectively with the DPP4 (CD26) Receptor? A Molecular Docking Study. Int. J. Mol. Sci. 2021, 22, 7001. [Google Scholar] [CrossRef] [PubMed]
- Clausen, T.; Sandoval, D.R.; Spliid, C.B.; Pihl, J.; Perrett, H.R.; Painter, C.D.; Narayanan, A.; Majowicz, S.A.; Kwong, E.M.; McVicar, R.N.; et al. SARS-CoV-2 Infection Depends on Cellular Heparan Sulfate and ACE2. Cell 2020, 183, 1043–1057.e15. [Google Scholar] [CrossRef]
- Rovas, A.; Osiaevi, I.; Buscher, K.; Sackarnd, J.; Tepasse, P.R.; Fobker, M.; Kühn, J.; Braune, S.; Göbel, U.; Thölking, G.; et al. Microvascular dysfunction in COVID-19: The MYSTIC study. Angiogenesis 2020, 24, 145–157. [Google Scholar] [CrossRef]
- Ontong, P.; Prachayasittikul, V. Unraveled roles of hyaluronan in severe COVID-19. EXCLI J. 2021, 20, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Esposito, S.; Jones, M.H.; Feleszko, W.; Martell, J.A.O.; Falup-Pecurariu, O.; Geppe, N.; Martinón-Torres, F.; Shen, K.L.; Roth, M.; Principi, N. Prevention of New Respiratory Episodes in Children with Recurrent Respiratory Infections: An Expert Consensus Statement. Microorganisms 2020, 8, 1810. [Google Scholar] [CrossRef]
- Cardinale, F.; Lombardi, E.; Rossi, O.; Bagnasco, D.; Bellocchi, A.; Menzella, F. Epithelial dysfunction, respiratory infections and asthma: The importance of immunomodulation. A focus on OM-85. Expert Rev. Respir. Med. 2020, 14, 1019–1026. [Google Scholar] [CrossRef]
- Dang, A.T.; Pasquali, C.; Ludigs, K.; Guarda, G. OM-85 is an immunomodulator of interferon-β production and inflammasome activity. Sci. Rep. 2017, 7, 43844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentley, J.K.; Han, M.; Jaipalli, S.; Hinde, J.L.; Lei, J.; Ishikawa, T.; Goldsmith, A.M.; Rajput, C.; Hershenson, M.B. Myristoylated rhinovirus VP4 protein activates TLR2-dependent proinflammatory gene expression. Am. J. Physiol. Lung Cell. Mol. Physiol. 2019, 317, L57–L70. [Google Scholar] [CrossRef]
- Luan, H.; Zhang, Q.; Wang, L.; Wang, C.; Zhang, M.; Xu, X.; Zhou, H.; Li, X.; Xu, Q.; He, F.; et al. OM85-BV induced the productions of IL-1β, IL-6, and TNF-α via TLR4- and TLR2-mediated ERK1/2/NF-κB pathway in RAW264.7 cells. J. Interferon Cytokine Res. 2014, 34, 526–536. [Google Scholar] [CrossRef]
- Parola, C.; Salogni, L.; Vaira, X.; Scutera, S.; Somma, P.; Salvi, V.; Musso, T.; Tabbia, G.; Bardessono, M.; Pasquali, C.; et al. Selective activation of human dendritic cells by OM-85 through a NF-kB and MAPK dependent pathway. PLoS ONE 2013, 8, e82867. [Google Scholar] [CrossRef]
- Roth, M.; Pasquali, C.; Stolz, D.; Tamm, M. Broncho Vaxom (OM-85) modulates rhinovirus docking proteins on human airway epithelial cells via Erk1/2 mitogen activated protein kinase and cAMP. PLoS ONE 2017, 12, e0188010. [Google Scholar] [CrossRef] [Green Version]
- Cao, C.; Wang, J.; Li, Y.; Li, Y.; Ma, L.; Abdelrahim, M.E.A.; Zhu, Y. Efficacy and safety of OM-85 in paediatric recurrent respiratory tract infections which could have a possible protective effect on COVID-19 pandemic: A meta-analysis. Int. J. Clin. Pract. 2021, 75, e13981. [Google Scholar] [CrossRef] [PubMed]
- Esposito, S.; Bianchini, S.; Bosis, S.; Tagliabue, C.; Coro, I.; Argentiero, A.; Principi, N. A randomized, placebo-controlled, double-blinded, single-centre, phase IV trial to assess the efficacy and safety of OM-85 in children suffering from recurrent respiratory tract infections. J. Transl. Med. 2019, 17, 284. [Google Scholar] [CrossRef]
- Sly, P.D.; Galbraith, S.; Islam, Z.; Holt, B.; Troy, N.; Holt, P.G. Primary prevention of severe lower respiratory illnesses in at-risk infants using the immunomodulatory OM-85. J. Allergy Clin. Immunol. 2019, 144, 870–872.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koatz, A.M.; Coe, N.A.; Cicerán, A.; Alter, A.J. Clinical and Immunological Benefits of OM-85 Bacterial Lysate in Patients with Allergic Rhinitis, Asthma, and COPD and Recurrent Respiratory Infections. Lung 2016, 194, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Li, Y.; Xu, L.; Xia, M.; Cao, L. Bacterial lysate increases the percentage of natural killer T cells in peripheral blood and alleviates asthma in children. Pharmacology 2015, 95, 139–144. [Google Scholar] [CrossRef]
- Pan, L.; Jiang, X.G.; Guo, J.; Tian, Y.; Liu, C.T. Effects of OM-85 BV in patients with chronic obstructive pulmonary disease: A systematic review and meta-analysis. J. Clin. Pharmacol. 2015, 55, 1086–1092. [Google Scholar] [CrossRef]
- Marcellini, A.; Swieboda, D.; Guedán, A.; Farrow, S.N.; Casolari, P.; Contoli, M.; Johnston, S.L.; Papi, A.; Solari, R. Glucocorticoids impair type I IFN signalling and enhance rhinovirus replication. Eur. J. Pharmacol. 2021, 893, 173839. [Google Scholar] [CrossRef]
- Ekbom, E.; Quint, J.; Schöler, L.; Malinovschi, A.; Franklin, K.; Holm, M.; Torén, K.; Lindberg, E.; Jarvis, D.; Janson, C. Asthma and treatment with inhaled corticosteroids: Associations with hospitalisations with pneumonia. BMC Pulm. Med. 2019, 19, 254. [Google Scholar] [CrossRef]
- Van den Eynde, E.; Gasch, O.; Oliva, J.C.; Prieto, E.; Calzado, S.; Gomila, A.; Machado, M.L.; Falgueras, L.; Ortonobes, S.; Morón, A.; et al. Corticosteroids and tocilizumab reduce in-hospital mortality in severe COVID-19 pneumonia: A retrospective study in a Spanish hospital. Infect. Dis. 2021, 53, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Luis, B.M.; Miguel, M.B.; Pedro, D.L.; David, I.P.; Itziar, A.; Ana, G.H.; Enrique, I.J.; María, L.V.; La Torre-Ferrera Noelia, D.; Julio César, B.B.; et al. Benefits of early aggressive immunomodulatory therapy (tocilizumab and methylprednisolone) in COVID-19: Single center cohort study of 685 patients. J. Transl. Autoimmun. 2021, 4, 100086. [Google Scholar] [CrossRef]
- Finney, L.J.; Glanville, N.; Farne, H.; Aniscenko, J.; Fenwick, P.; Kemp, S.V.; Trujillo-Torralbo, M.B.; Loo, S.L.; Calderazzo, M.A.; Wedzicha, J.A.; et al. Inhaled corticosteroids downregulate the SARS-CoV-2 receptor ACE2 in COPD through suppression of type I interferon. J. Allergy Clin. Immunol. 2021, 147, 510–519.e5. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, S.; Wang, J.; Xue, Z.; Wang, C.; Wang, N. Dexamethasone inhibits SARS-CoV-2 spike pseudotyped virus viropexis by binding to ACE2. Virology 2021, 554, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Z.; Liu, J.; Shi, D.; Chen, W.; Li, J.; Yan, R.; Bi, Y.; Hu, W.; Zhu, Z.; Yu, Y.; et al. Glucocorticoids improve severe or critical COVID-19 by activating ACE2 and reducing IL-6 levels. Int. J. Biol. Sci. 2020, 16, 2382–2391. [Google Scholar] [CrossRef]
- Papamanoli, A.; Yoo, J.; Grewal, P.; Predun, W.; Hotelling, J.; Jacob, R.; Mojahedi, A.; Skopicki, H.A.; Mansour, M.; Marcos, L.A.; et al. High-dose methylprednisolone in nonintubated patients with severe COVID-19 pneumonia. Eur. J. Clin. Investig. 2021, 51, e13458. [Google Scholar] [CrossRef] [PubMed]
- Salton, F.; Confalonieri, P.; Meduri, G.U.; Santus, P.; Harari, S.; Scala, R.; Lanini, S.; Vertui, V.; Oggionni, T.; Caminati, A.; et al. Prolonged Low-Dose Methylprednisolone in Patients With Severe COVID-19 Pneumonia. Open Forum Infect. Dis. 2020, 7, ofaa421. [Google Scholar] [CrossRef] [PubMed]
- Chhapola Shukla, S. ACE2 expression in allergic airway disease may decrease the risk and severity of COVID-19. Eur. Arch. Oto-Rhino-Laryngol. 2021, 278, 2637–2640. [Google Scholar] [CrossRef] [PubMed]
- Drago, L.; Cappelletti, L.; De Vecchi, E.; Pignataro, L.; Torretta, S.; Mattina, R. Antiadhesive and antibiofilm activity of hyaluronic acid against bacteria responsible for respiratory tract infections. APMIS 2014, 122, 1013–1019. [Google Scholar] [CrossRef]
- Cermelli, C.; Cuoghi, A.; Scuri, M.; Bettua, C.; Neglia, R.G.; Ardizzoni, A.; Blasi, E.; Iannitti, T.; Palmieri, B. In vitro evaluation of antiviral and virucidal activity of a high molecular weight hyaluronic acid. Virol. J. 2011, 8, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, P.; Bhardwaj, T.; Kumar, A.; Gehi, B.R.; Kapuganti, S.K.; Garg, N.; Nath, G.; Giri, R. Reprofiling of approved drugs against SARS-CoV-2 main protease: An in-silico study. J. Biomol. Struct. Dyn. 2020, 1–15. [Google Scholar] [CrossRef]
- Hellman, U.; Karlsson, M.G.; Engström-Laurent, A.; Cajander, S.; Dorofte, L.; Ahlm, C.; Laurent, C.; Blomberg, A. Presence of hyaluronan in lung alveoli in severe Covid-19: An opening for new treatment options? J. Biol. Chem. 2020, 295, 15418–15422. [Google Scholar] [CrossRef] [PubMed]
- Andonegui-Elguera, S.; Taniguchi-Ponciano, K.; Gonzalez-Bonilla, C.R.; Torres, J.; Mayani, H.; Herrera, L.A.; Peña-Martínez, E.; Silva-Román, G.; Vela-Patiño, S.; Ferreira-Hermosillo, A.; et al. Molecular Alterations Prompted by SARS-CoV-2 Infection: Induction of Hyaluronan, Glycosaminoglycan and Mucopolysaccharide Metabolism. Arch. Med. Res. 2020, 51, 645–653. [Google Scholar] [CrossRef]
- Evans, J.P.; Liu, S.L. Role of host factors in SARS-CoV-2 entry. J. Biol. Chem. 2021, 297, 100847. [Google Scholar] [CrossRef]
- Yu, M.; Zhang, T.; Zhang, W.; Sun, Q.; Li., H.; Li, J.P. Elucidating the Interactions Between Heparin/Heparan Sulfate and SARS-CoV-2-Related Proteins-An Important Strategy for Developing Novel Therapeutics for the COVID-19 Pandemic. Front. Mol. Biosci. 2021, 7, 628551. [Google Scholar] [CrossRef]
- Hu, Y.; Meng, X.; Zhang, F.; Xiang, Y.; Wang, J. The in vitro antiviral activity of lactoferrin against common human coronaviruses and SARS-CoV-2 is mediated by targeting the heparan sulfate co-receptor. Emerg. Microbes Infect. 2021, 10, 317–330. [Google Scholar] [CrossRef]
- Kalra, R.S.; Kandimalla, R. Engaging the spikes: Heparan sulfate facilitates SARS-CoV-2 spike protein binding to ACE2 and potentiates viral infection. Signal Transduct. Target. Ther. 2021, 6, 39. [Google Scholar] [CrossRef]
- Martino, C.; Kellman, B.P.; Sandoval, D.R.; Clausen, T.M.; Marotz, C.A.; Song, S.J.; Wandro, S.; Zaramela, L.S.; Salido Benítez, R.A.; Zhu, Q.; et al. Bacterial modification of the host glycosaminoglycan heparan sulfate modulates SARS-CoV-2 infectivity. bioRxiv 2020, 18. [Google Scholar] [CrossRef]
- Cheudjeu, A. Antiviral strategies should focus on stimulating the biosynthesis of heparan sulfates, not their inhibition. Life Sci. 2021, 277, 119508. [Google Scholar] [CrossRef]
- Mycroft-West, C.J.; Su, D.; Pagani, I.; Rudd, T.R.; Elli, S.; Gandhi, N.S.; Guimond, S.E.; Miller, G.J.; Meneghetti, M.C.Z.; Nader, H.B.; et al. Heparin Inhibits Cellular Invasion by SARS-CoV-2: Structural Dependence of the Interaction of the Spike S1 Receptor-Binding Domain with Heparin. Thromb. Haemost. 2020, 120, 1700–1715. [Google Scholar] [CrossRef] [PubMed]
- Tandon, R.; Sharp, J.S.; Zhang, F.; Pomin, V.H.; Ashpole, N.M.; Mitra, D.; Jin, W.; Liu, H.; Sharma, P.; Linhardt, R.J. Effective Inhibition of SARS-CoV-2 Entry by Heparin and Enoxaparin Derivatives. bioRxiv 2020, 95, e01987-20. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Species/Clonality | Cat # | Supplier |
|---|---|---|---|
| ACE2 | Mouse monoclonal | MAB933 | R&D SYSTEMS |
| TMPRSS2 | Rabbit polyclonal | HAP035787 | Sigma-Aldrich |
| ADAM17 | Rabbit monoclonal | NBP2-67179 | Novus Biologicals |
| DPP4 | Mouse monoclonal | ab114033 | Abcam |
| Primers | Sequence (5′–3′) | Length | Tm | Location | Amplicon Size |
|---|---|---|---|---|---|
| ACE2_Forward | CGAAGCCGAAGACCTGTTCTA | 21 | 61.5 | 102–122 | 140 |
| ACE2_Reverse | GGGCAAGTGTGGACTGTTCC | 20 | 63 | 241–222 | |
| TMPRSS2_Forward | CAAGTGCTCCAACTCTGGGAT | 21 | 61.8 | 444–464 | 115 |
| TMPRSS2_Reverse | AACACACCGATTCTCGTCCTC | 21 | 61.8 | 558–538 | |
| DPP4_Forward | TACAAAAGTGACATGCCTCAGTT | 23 | 60.1 | 1317–1339 | 134 |
| DPP4_Reverse | TGTGTAGAGTATAGAGGGGCAGA | 23 | 61.1 | 1450–1428 | |
| ADAM17_Forward | GTGGATGGTAAAAACGAAAGCG | 22 | 60.4 | 295–316 | 93 |
| ADAM17_Reverse | GGCTAGAACCCTAGAGTCAGG | 21 | 60.4 | 387–367 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, L.; Zhou, L.; Tamm, M.; Roth, M. OM-85 Broncho-Vaxom®, a Bacterial Lysate, Reduces SARS-CoV-2 Binding Proteins on Human Bronchial Epithelial Cells. Biomedicines 2021, 9, 1544. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9111544
Fang L, Zhou L, Tamm M, Roth M. OM-85 Broncho-Vaxom®, a Bacterial Lysate, Reduces SARS-CoV-2 Binding Proteins on Human Bronchial Epithelial Cells. Biomedicines. 2021; 9(11):1544. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9111544
Chicago/Turabian StyleFang, Lei, Liang Zhou, Michael Tamm, and Michael Roth. 2021. "OM-85 Broncho-Vaxom®, a Bacterial Lysate, Reduces SARS-CoV-2 Binding Proteins on Human Bronchial Epithelial Cells" Biomedicines 9, no. 11: 1544. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9111544