
Repeated α-GalCer Administration Induces a Type 2 Cytokine-Biased iNKT Cell Response and Exacerbates Atopic Skin Inflammation in Vα14Tg NC/Nga Mice

 , , , , ,
, , , , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Mice
2.3. Cell Isolation and Culture
2.4. Flow Cytometry
2.5. Intracellular Cytokine Staining
2.6. ELISA
2.7. Analysis of Skin Sections
2.8. Scoring the Severity of Skin Lesions
2.9. Intraperitoneal Injection of α-GalCer into Mice
2.10. Statistical Analysis
3. Results
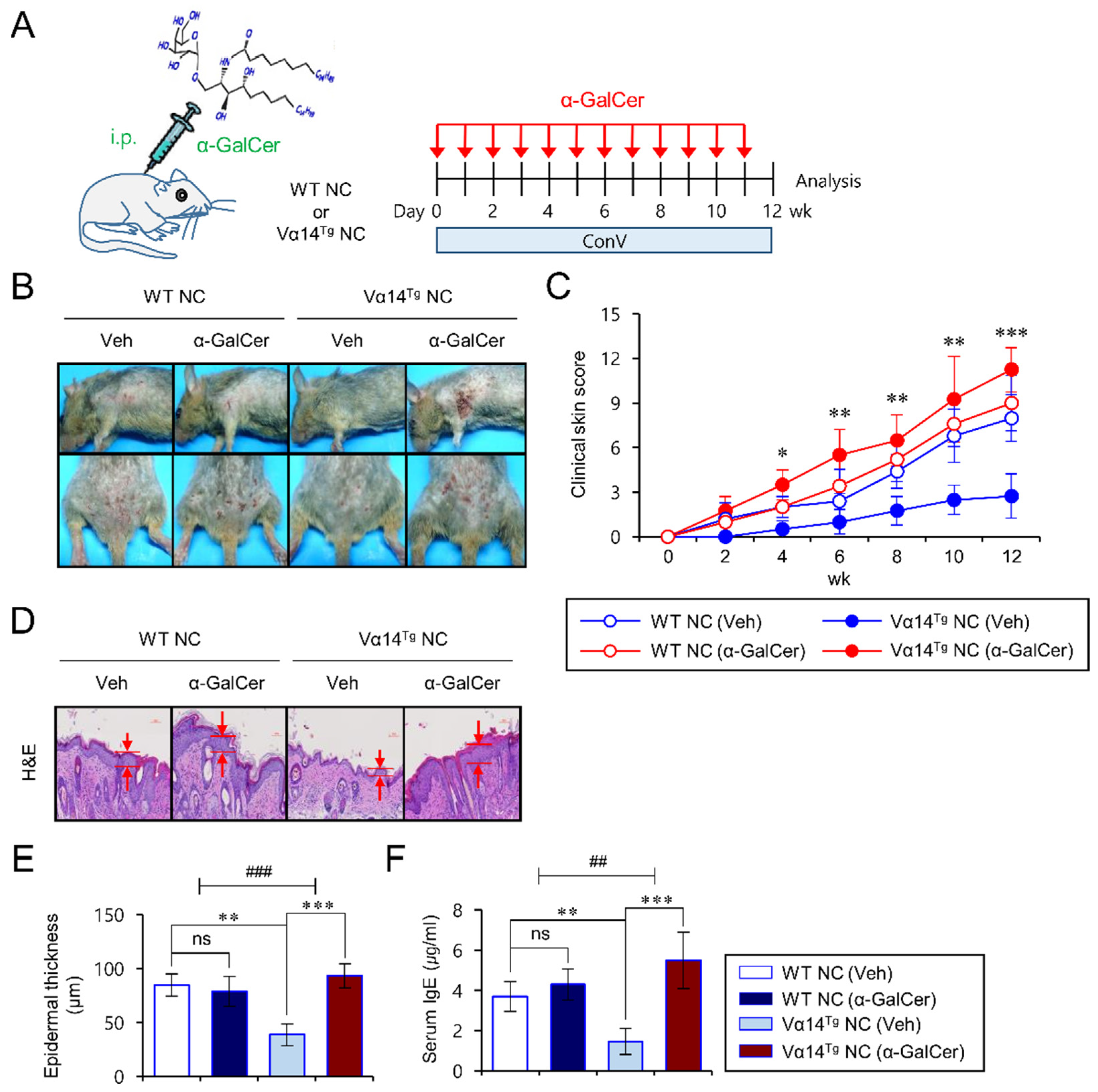
3.1. Vα14Tg NC Mice Become Susceptible to AD upon Repeated α-GalCer Treatment
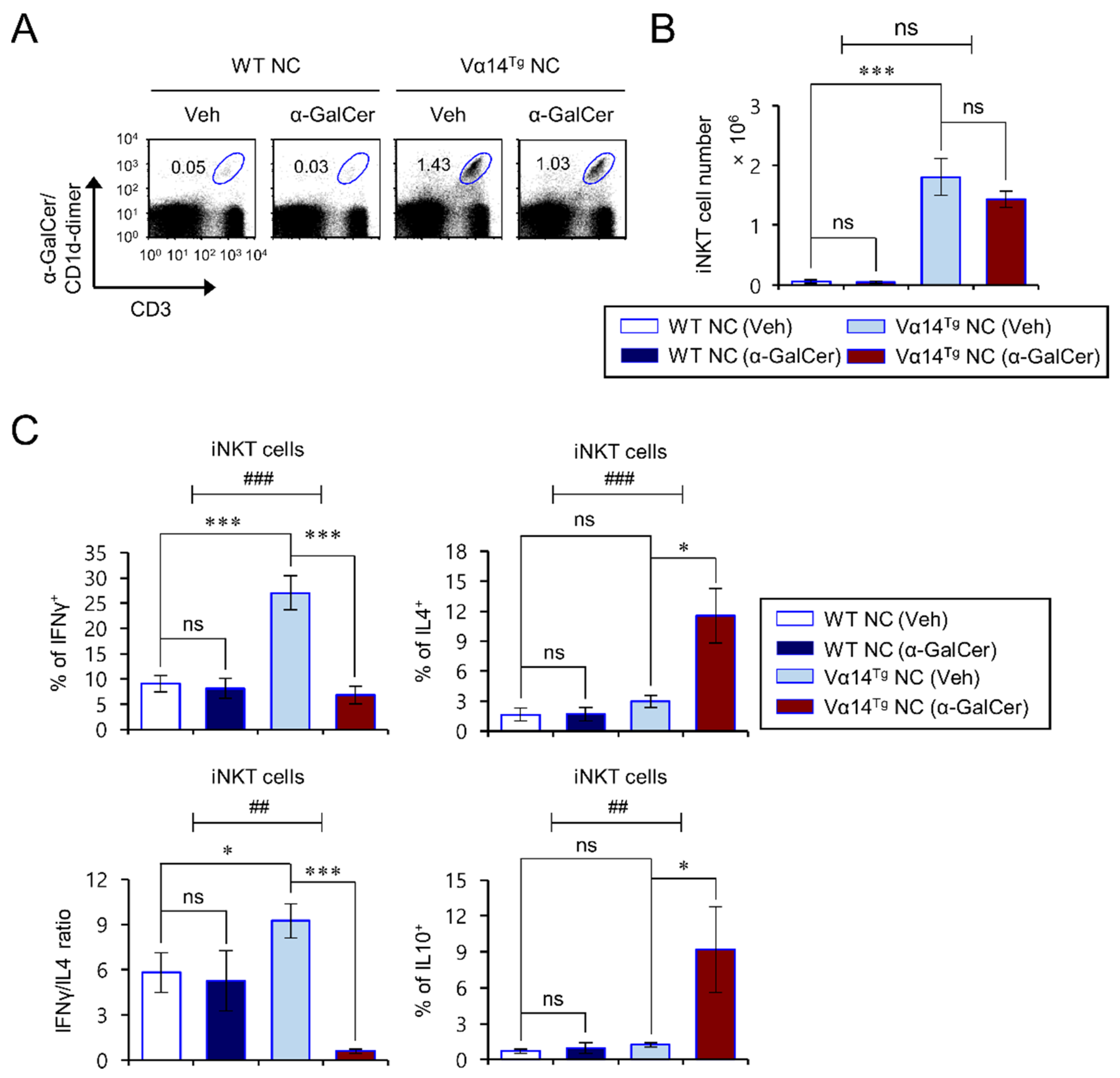
3.2. Repeated In Vivo α-GalCer Treatment Biases iNKT Cells of Vα14Tg NC Mice towards Type 2 Cytokine Production
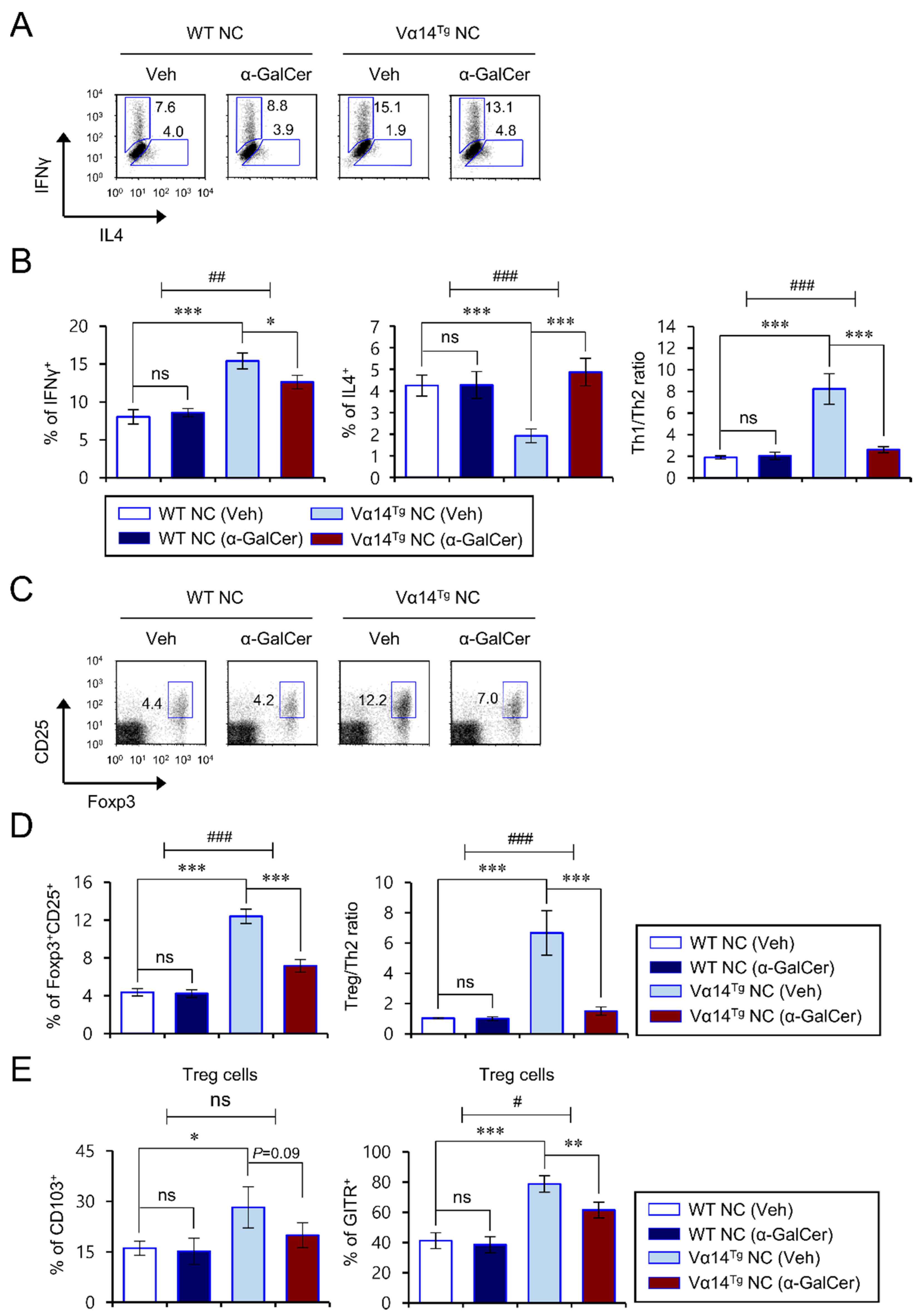
3.3. Alpha-GalCer-Mediated Exacerbation of AD Pathogenesis Correlates with a Reduced Th1/Th2 Cytokine Ratio of CD4+ T Cells in Vα14Tg NC Mice
3.4. Repeated α-GalCer Treatment Contracts Splenic Treg Cells in Vα14Tg NC Mice
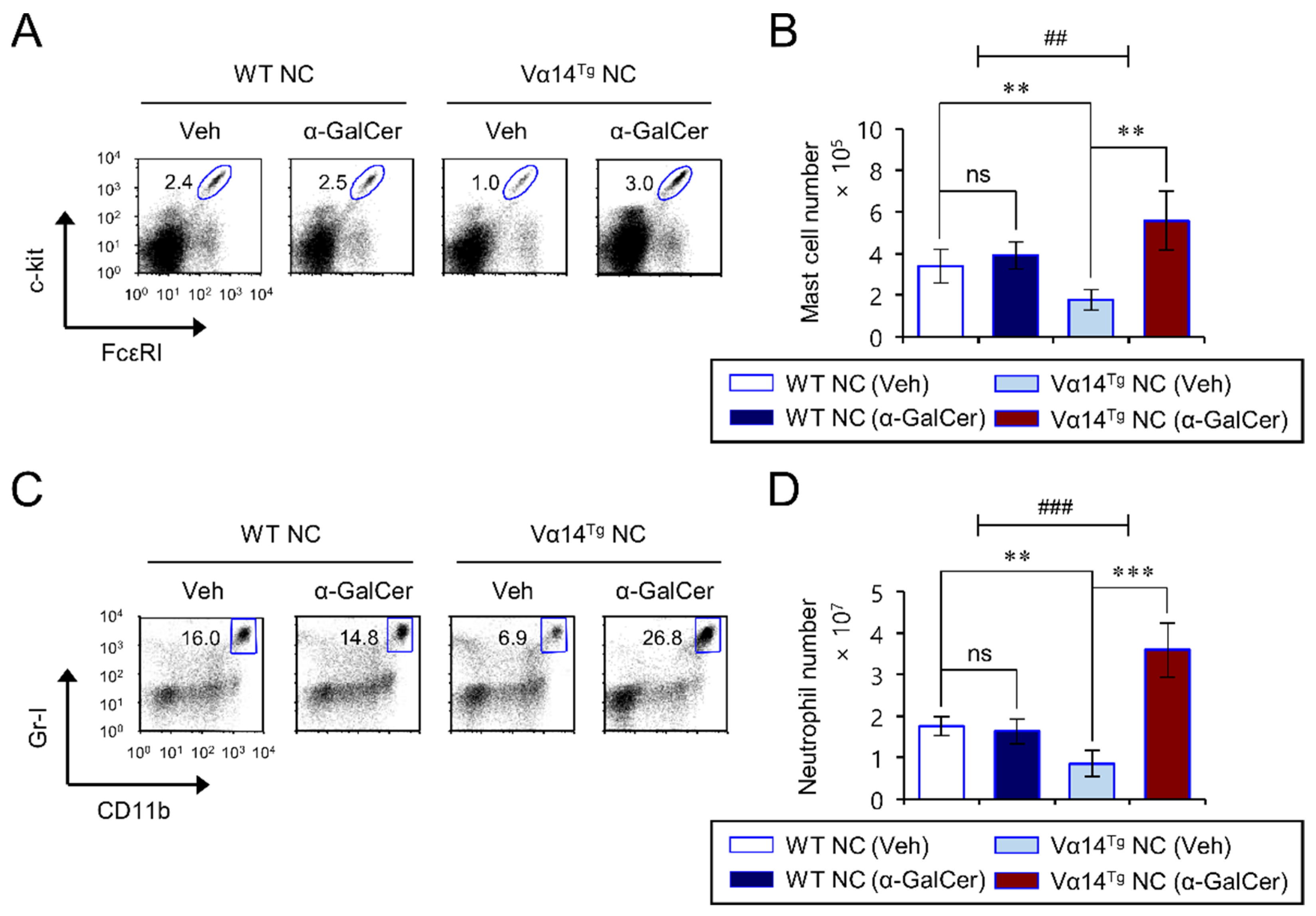
3.5. Repeated α-GalCer-Injection Expands Splenic Mast Cells and Neutrophils in Vα14Tg NC Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Park, H.J.; Lee, S.W.; Hong, S. Regulation of Allergic Immune Responses by Microbial Metabolites. Immune Netw. 2018, 18, e15. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Park, H.J.; Jeon, J.; Park, Y.H.; Kim, T.C.; Jeon, S.H.; Seong, R.H.; Van Kaer, L.; Hong, S. Ubiquitous Overexpression of Chromatin Remodeling Factor SRG3 Exacerbates Atopic Dermatitis in NC/Nga Mice by Enhancing Th2 Immune Responses. Int. J. Mol. Sci. 2021, 22, 1553. [Google Scholar] [PubMed]
- Furue, M. Regulation of Skin Barrier Function via Competition between AHR Axis versus IL-13/IL-4JAKSTAT6/STAT3 Axis: Pathogenic and Therapeutic Implications in Atopic Dermatitis. J. Clin. Med. 2020, 9, 3741. [Google Scholar]
- Weidinger, S.; Beck, L.A.; Bieber, T.; Kabashima, K.; Irvine, A.D. Atopic dermatitis. Nat. Rev. Dis. Primers 2018, 4, 1. [Google Scholar] [CrossRef]
- Hattori, K.; Nishikawa, M.; Watcharanurak, K.; Ikoma, A.; Kabashima, K.; Toyota, H.; Takahashi, Y.; Takahashi, R.; Watanabe, Y.; Takakura, Y. Sustained exogenous expression of therapeutic levels of IFN-gamma ameliorates atopic dermatitis in NC/Nga mice via Th1 polarization. J. Immunol. 2010, 184, 2729–2735. [Google Scholar] [PubMed] [Green Version]
- Jin, H.; He, R.; Oyoshi, M.; Geha, R.S. Animal models of atopic dermatitis. J. Investig. Dermatol. 2009, 129, 31–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takano, N.; Arai, I.; Kurachi, M. Analysis of the spontaneous scratching behavior by NC/Nga mice: A possible approach to evaluate antipruritics for subjects with atopic dermatitis. Eur. J. Pharmacol. 2003, 471, 223–228. [Google Scholar] [CrossRef]
- Vestergaard, C.; Yoneyama, H.; Murai, M.; Nakamura, K.; Tamaki, K.; Terashima, Y.; Imai, T.; Yoshie, O.; Irimura, T.; Mizutani, H.; et al. Overproduction of Th2-specific chemokines in NC/Nga mice exhibiting atopic dermatitis-like lesions. J. Clin. Investig. 1999, 104, 1097–1105. [Google Scholar] [CrossRef]
- Van Kaer, L.; Wu, L. Therapeutic Potential of Invariant Natural Killer T Cells in Autoimmunity. Front. Immunol. 2018, 9, 519. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Driver, J.P.; Van Kaer, L. The Role of Autophagy in iNKT Cell Development. Front. Immunol. 2018, 9, 2653. [Google Scholar] [CrossRef]
- Kumar, A.; Suryadevara, N.; Hill, T.M.; Bezbradica, J.S.; Van Kaer, L.; Joyce, S. Natural Killer T Cells: An Ecological Evolutionary Developmental Biology Perspective. Front. Immunol. 2017, 8, 1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, Y.T.; Leung, K.; Chang, Y.J.; DeKruyff, R.H.; Savage, P.B.; Cruse, R.; Benoit, C.; Elewaut, D.; Baumgarth, N.; Umetsu, D.T. A natural killer T-cell subset that protects against airway hyperreactivity. J. Allergy Clin. Immunol. 2019, 143, 565–576.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.J.; Lee, S.W.; Park, S.H.; Van Kaer, L.; Hong, S. Selective Expansion of Double Negative iNKT Cells Inhibits the Development of Atopic Dermatitis in Valpha14 TCR Transgenic NC/Nga Mice by Increasing Memory-Type CD8(+) T and Regulatory CD4(+) T Cells. J. Investig. Dermatol. 2020, 141, 1512–1521. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Lee, S.W.; Van Kaer, L.; Hong, S. CD1d-Dependent iNKT Cells Control DSS-Induced Colitis in a Mouse Model of IFNgamma-Mediated Hyperinflammation by Increasing IL22-Secreting ILC3 Cells. Int. J. Mol. Sci. 2021, 22, 1250. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Park, H.J.; Cheon, J.H.; Wu, L.; Van Kaer, L.; Hong, S. iNKT Cells Suppress Pathogenic NK1.1(+)CD8(+) T Cells in DSS-Induced Colitis. Front. Immunol. 2018, 9, 2168. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.J.; Kim, H.Y.; Albacker, L.A.; Lee, H.H.; Baumgarth, N.; Akira, S.; Savage, P.B.; Endo, S.; Yamamura, T.; Maaskant, J.; et al. Influenza infection in suckling mice expands an NKT cell subset that protects against airway hyperreactivity. J. Clin. Investig. 2011, 121, 57–69. [Google Scholar] [CrossRef]
- Morita, M.; Motoki, K.; Akimoto, K.; Natori, T.; Sakai, T.; Sawa, E.; Yamaji, K.; Koezuka, Y.; Kobayashi, E.; Fukushima, H. Structure-activity relationship of alpha-galactosylceramides against B16-bearing mice. J. Med. Chem. 1995, 38, 2176–2187. [Google Scholar] [CrossRef]
- Wang, H.; Feng, D.; Park, O.; Yin, S.; Gao, B. Invariant NKT cell activation induces neutrophil accumulation and hepatitis: Opposite regulation by IL-4 and IFN-gamma. Hepatology 2013, 58, 1474–1485. [Google Scholar]
- Lee, S.W.; Park, H.J.; Van Kaer, L.; Hong, S.; Hong, S. Graphene oxide polarizes iNKT cells for production of TGFbeta and attenuates inflammation in an iNKT cell-mediated sepsis model. Sci. Rep. 2018, 8, 10081. [Google Scholar] [CrossRef]
- Park, H.J.; Lee, S.W.; Im, W.; Kim, M.; Van Kaer, L.; Hong, S. iNKT Cell Activation Exacerbates the Development of Huntington’s Disease in R6/2 Transgenic Mice. Mediat. Inflamm. 2019, 2019, 3540974. [Google Scholar]
- Sag, D.; Krause, P.; Hedrick, C.C.; Kronenberg, M.; Wingender, G. IL-10-producing NKT10 cells are a distinct regulatory invariant NKT cell subset. J. Clin. Investig. 2014, 124, 3725–3740. [Google Scholar] [CrossRef] [PubMed]
- Uchida, T.; Nakashima, H.; Yamagata, A.; Ito, S.; Ishikiriyama, T.; Nakashima, M.; Seki, S.; Kumagai, H.; Oshima, N. Repeated administration of alpha-galactosylceramide ameliorates experimental lupus nephritis in mice. Sci. Rep. 2018, 8, 8225. [Google Scholar] [CrossRef]
- Ju, A.; Lee, S.W.; Lee, Y.E.; Han, K.C.; Kim, J.C.; Shin, S.C.; Park, H.J.; EunKyeong Kim, E.; Hong, S.; Jang, M. A carrier-free multiplexed gene editing system applicable for suspension cells. Biomaterials 2019, 217, 119298. [Google Scholar] [PubMed]
- Hachem, P.; Lisbonne, M.; Michel, M.L.; Diem, S.; Roongapinun, S.; Lefort, J.; Marchal, G.; Herbelin, A.; Askenase, P.W.; Dy, M.; et al. Alpha-galactosylceramide-induced iNKT cells suppress experimental allergic asthma in sensitized mice: Role of IFN-gamma. Eur. J. Immunol. 2005, 35, 2793–2802. [Google Scholar] [CrossRef] [PubMed]
- Gaspar-Elsas, M.I.; Queto, T.; Masid-de-Brito, D.; Vieira, B.M.; de Luca, B.; Cunha, F.Q.; Xavier-Elsas, P. alpha-Galactosylceramide suppresses murine eosinophil production through interferon-gamma-dependent induction of NO synthase and CD95. Br. J. Pharmacol. 2015, 172, 3313–3325. [Google Scholar]
- Takahashi, N.; Akahoshi, M.; Matsuda, A.; Ebe, K.; Inomata, N.; Obara, K.; Hirota, T.; Nakashima, K.; Shimizu, M.; Tamari, M.; et al. Association of the IL12RB1 promoter polymorphisms with increased risk of atopic dermatitis and other allergic phenotypes. Hum. Mol. Genet. 2005, 14, 3149–3159. [Google Scholar] [CrossRef] [Green Version]
- Itazawa, T.; Adachi, Y.; Okabe, Y.; Hamamichi, M.; Adachi, Y.S.; Toyoda, M.; Morohashi, M.; Miyawaki, T. Developmental changes in interleukin-12-producing ability by monocytes and their relevance to allergic diseases. Clin. Exp. Allergy 2003, 33, 525–530. [Google Scholar] [CrossRef]
- Fyhrquist, N.; Lehtimaki, S.; Lahl, K.; Savinko, T.; Lappetelainen, A.M.; Sparwasser, T.; Wolff, H.; Lauerma, A.; Alenius, H. Foxp3+ cells control Th2 responses in a murine model of atopic dermatitis. J. Investig. Dermatol. 2012, 132, 1672–1680. [Google Scholar]
- Kalekar, L.A.; Rosenblum, M.D. Regulatory T cells in inflammatory skin disease: From mice to humans. Int. Immunol. 2019, 31, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Oyoshi, M.K.; He, R.; Li, Y.; Mondal, S.; Yoon, J.; Afshar, R.; Chen, M.; Lee, D.M.; Luo, H.R.; Luster, A.D.; et al. Leukotriene B4-driven neutrophil recruitment to the skin is essential for allergic skin inflammation. Immunity 2012, 37, 747–758. [Google Scholar] [CrossRef]
- Voisin, T.; Chiu, I.M. Molecular link between itch and atopic dermatitis. Proc. Natl. Acad. Sci. USA 2018, 115, 12851–12853. [Google Scholar]
- Walsh, C.M.; Hill, R.Z.; Schwendinger-Schreck, J.; Deguine, J.; Brock, E.C.; Kucirek, N.; Rifi, Z.; Wei, J.; Gronert, K.; Brem, R.B.; et al. Neutrophils promote CXCR3-dependent itch in the development of atopic dermatitis. Elife 2019, 8, e48448. [Google Scholar] [CrossRef] [PubMed]
- Brennan, P.J.; Tatituri, R.V.; Brigl, M.; Kim, E.Y.; Tuli, A.; Sanderson, J.P.; Gadola, S.D.; Hsu, F.F.; Besra, G.S.; Brenner, M.B. Invariant natural killer T cells recognize lipid self antigen induced by microbial danger signals. Nat. Immunol. 2011, 12, 1202–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darmoise, A.; Teneberg, S.; Bouzonville, L.; Brady, R.O.; Beck, M.; Kaufmann, S.H.; Winau, F. Lysosomal alpha-galactosidase controls the generation of self lipid antigens for natural killer T cells. Immunity 2010, 33, 216–228. [Google Scholar] [CrossRef] [Green Version]
- Wingender, G.; Rogers, P.; Batzer, G.; Lee, M.S.; Bai, D.; Pei, B.; Khurana, A.; Kronenberg, M.; Horner, A.A. Invariant NKT cells are required for airway inflammation induced by environmental antigens. J. Exp. Med. 2011, 208, 1151–1162. [Google Scholar] [CrossRef]
- Albacker, L.A.; Chaudhary, V.; Chang, Y.J.; Kim, H.Y.; Chuang, Y.T.; Pichavant, M.; DeKruyff, R.H.; Savage, P.B.; Umetsu, D.T. Invariant natural killer T cells recognize a fungal glycosphingolipid that can induce airway hyperreactivity. Nat. Med. 2013, 19, 1297–1304. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Zeng, B.; Chen, J.; Cui, G.; Lu, C.; Wu, W.; Yang, J.; Wei, H.; Xue, R.; Bai, L.; et al. Enterogenous bacterial glycolipids are required for the generation of natural killer T cells mediated liver injury. Sci. Rep. 2016, 6, 36365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wingender, G.; Stepniak, D.; Krebs, P.; Lin, L.; McBride, S.; Wei, B.; Braun, J.; Mazmanian, S.K.; Kronenberg, M. Intestinal microbes affect phenotypes and functions of invariant natural killer T cells in mice. Gastroenterology 2012, 143, 418–428. [Google Scholar] [CrossRef] [Green Version]
- Lai, A.C.; Chi, P.Y.; Thio, C.L.; Han, Y.C.; Kao, H.N.; Hsieh, H.W.; Gervay-Hague, J.; Chang, Y.J. alpha-Lactosylceramide Protects Against iNKT-Mediated Murine Airway Hyperreactivity and Liver Injury Through Competitive Inhibition of Cd1d Binding. Front. Chem. 2019, 7, 811. [Google Scholar] [CrossRef] [Green Version]
- Lombardi, V.; Stock, P.; Singh, A.K.; Kerzerho, J.; Yang, W.; Sullivan, B.A.; Li, X.; Shiratsuchi, T.; Hnatiuk, N.E.; Howell, A.R.; et al. A CD1d-dependent antagonist inhibits the activation of invariant NKT cells and prevents development of allergen-induced airway hyperreactivity. J. Immunol. 2010, 184, 2107–2115. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.J.; Wang, H.; Starrett, G.J.; Phuong, V.; Jameson, S.C.; Hogquist, K.A. Tissue-Specific Distribution of iNKT Cells Impacts Their Cytokine Response. Immunity 2015, 43, 566–578. [Google Scholar]
- Wedman, P.A.; Aladhami, A.; Chumanevich, A.P.; Fuseler, J.W.; Oskeritzian, C.A. Mast cells and sphingosine-1-phosphate underlie prelesional remodeling in a mouse model of eczema. Allergy 2018, 73, 405–415. [Google Scholar] [CrossRef]
- Sehra, S.; Serezani, A.P.M.; Ocana, J.A.; Travers, J.B.; Kaplan, M.H. Mast Cells Regulate Epidermal Barrier Function and the Development of Allergic Skin Inflammation. J. Investig. Dermatol. 2016, 136, 1429–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hershko, A.Y.; Suzuki, R.; Charles, N.; Alvarez-Errico, D.; Sargent, J.L.; Laurence, A.; Rivera, J. Mast cell interleukin-2 production contributes to suppression of chronic allergic dermatitis. Immunity 2011, 35, 562–571. [Google Scholar] [PubMed] [Green Version]
- Li, W.; Ji, F.; Zhang, Y.; Wang, Y.; Yang, N.; Ge, H.; Wang, F. Cooperation of invariant NKT cells and CD4+CD25+ T regulatory cells in prevention of autoimmune diabetes in non-obese diabetic mice treated with alpha-galactosylceramide. Acta Biochim. Biophys. Sin. 2008, 40, 381–390. [Google Scholar]
- Venken, K.; Decruy, T.; Aspeslagh, S.; Van Calenbergh, S.; Lambrecht, B.N.; Elewaut, D. Bacterial CD1d-restricted glycolipids induce IL-10 production by human regulatory T cells upon cross-talk with invariant NKT cells. J. Immunol. 2013, 191, 2174–2183. [Google Scholar] [CrossRef] [PubMed]
- Magnan, A.; Mely, L.; Prato, S.; Vervloet, D.; Romagne, F.; Camilla, C.; Necker, A.; Casano, B.; Montero-Jullian, F.; Fert, V.; et al. Relationships between natural T cells, atopy, IgE levels, and IL-4 production. Allergy 2000, 55, 286–290. [Google Scholar] [CrossRef]
- Oishi, Y.; Sakamoto, A.; Kurasawa, K.; Nakajima, H.; Nakao, A.; Nakagawa, N.; Tanabe, E.; Saito, Y.; Iwamoto, I. CD4-CD8- T cells bearing invariant Valpha24JalphaQ TCR alpha-chain are decreased in patients with atopic diseases. Clin. Exp. Immunol. 2000, 119, 404–411. [Google Scholar] [CrossRef]
- Takahashi, T.; Nakamura, K.; Chiba, S.; Kanda, Y.; Tamaki, K.; Hirai, H. V alpha 24+ natural killer T cells are markedly decreased in atopic dermatitis patients. Hum. Immunol. 2003, 64, 586–592. [Google Scholar] [CrossRef]
- Dardalhon, V.; Awasthi, A.; Kwon, H.; Galileos, G.; Gao, W.; Sobel, R.A.; Mitsdoerffer, M.; Strom, T.B.; Elyaman, W.; Ho, I.C.; et al. IL-4 inhibits TGF-beta-induced Foxp3+ T cells and, together with TGF-beta, generates IL-9+ IL-10+ Foxp3(−) effector T cells. Nat. Immunol. 2008, 9, 1347–1355. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, H.J.; Kim, T.-C.; Park, Y.H.; Lee, S.W.; Jeon, J.; Park, S.-H.; Van Kaer, L.; Hong, S. Repeated α-GalCer Administration Induces a Type 2 Cytokine-Biased iNKT Cell Response and Exacerbates Atopic Skin Inflammation in Vα14Tg NC/Nga Mice. Biomedicines 2021, 9, 1619. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9111619
Park HJ, Kim T-C, Park YH, Lee SW, Jeon J, Park S-H, Van Kaer L, Hong S. Repeated α-GalCer Administration Induces a Type 2 Cytokine-Biased iNKT Cell Response and Exacerbates Atopic Skin Inflammation in Vα14Tg NC/Nga Mice. Biomedicines. 2021; 9(11):1619. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9111619
Chicago/Turabian StylePark, Hyun Jung, Tae-Cheol Kim, Yun Hoo Park, Sung Won Lee, Jungmin Jeon, Se-Ho Park, Luc Van Kaer, and Seokmann Hong. 2021. "Repeated α-GalCer Administration Induces a Type 2 Cytokine-Biased iNKT Cell Response and Exacerbates Atopic Skin Inflammation in Vα14Tg NC/Nga Mice" Biomedicines 9, no. 11: 1619. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9111619