
Functional Role of the RNA-Binding Protein Rbm24a and Its Target sox2 in Microphthalmia


1
Department of Biology, University of Iowa, Iowa City, IA 52245, USA
2
Mercury Data Science, Houston, TX 77098, USA
3
Department of Biostatistics, University of Iowa, Iowa City, IA 52245, USA
*
Author to whom correspondence should be addressed.
Biomedicines 2021, 9(2), 100; https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9020100
Submission received: 22 December 2020
/
Revised: 13 January 2021
/
Accepted: 16 January 2021
/
Published: 21 January 2021
(This article belongs to the Special Issue Zebrafish Models for Development and Disease 2.0)
Abstract
:Congenital eye defects represent a large class of disorders affecting roughly 21 million children worldwide. Microphthalmia and anophthalmia are relatively common congenital defects, with approximately 20% of human cases caused by mutations in SOX2. Recently, we identified the RNA-binding motif protein 24a (Rbm24a) which binds to and regulates sox2 in zebrafish and mice. Here we show that morpholino knockdown of rbm24a leads to microphthalmia and visual impairment. By utilizing sequential injections, we demonstrate that addition of exogenous sox2 RNA to rbm24a-deplete embryos is sufficient to suppress morphological and visual defects. This research demonstrates a critical role for understanding the post-transcriptional regulation of genes needed for development.
1. Introduction
Congenital eye defects represent a large class of disorders affecting roughly 21 million children worldwide [1]. Defects can affect every part of the eye from the retina to the lens, but can also include the eye as a whole. Microphthalmia, a smaller than normal eye(s), and anophthalmia, a lack of an eye(s), are both relatively common congenital defects affecting between 1 in 7000 and 1 in 30,000 live births, respectively [2,3,4,5].
Mutations in SRY (sex determining region Y)-box2 (SOX2) account for approximately 20% of human anophthalmia cases [6]. As a transcription factor, SOX2, often in cooperation with a partner transcription factor, is responsible for the regulation of many genes and is one of the Yamanaka reprogramming factors that regulates pluripotency [7,8,9]. Thus, understanding the regulation of SOX2 is crucial. While the transcriptional regulation of SOX2 has been explored, only recently has the post-transcriptional aspect of its regulation been appreciated [10].
Previously, we identified the RNA-binding protein, RBM24, as a post-transcriptional regulator of Sox2 [11,12]. Consistent with SOX2′s known role in microphthalmia and anophthalmia in human patients, knockdown and knockout Rbm24-deficient mice and zebrafish often displayed eye defects including microphthalmia and/or anophthalmia [12,13]. In addition to its role in eye development, rbm24a knockdown, mutation, and overexpression are also associated with cardiac defects. Previous inquiries found Rbm24 mutant mice to be embryonic lethal, dying between E7.5 and E14.5. The embryonic lethality observed is thought to be cardiomyopathy due to numerous cardiovascular malformations, which included ventricular septum defects, reduced trabeculation and compaction, dilated atria and ventricle chambers, thinner atrioventricular endocardial cushions, sarcomere disarray, and fibrosis [12,14,15,16]. Most previous research focused on the cardiac phenotypes associated with rbm24a depletion.
It is challenging to study global factors such as Rbm24a due to their wide-ranging impacts on a multitude of target RNAs. We chose to focus on a single proposed RNA target, sox2, to perform more in-depth analyses as to the relationship between Rbm24a and sox2. To do so, we analyzed the expression pattern of rbm24a and found, consistent with previous studies in zebrafish, Xenopus, mice, and chick, its expression in the somites, heart, and lens in a spatiotemporal manner [11,14,17,18,19,20,21,22,23]. To study the role of rbm24a in zebrafish development, we performed morpholino knockdown and CRISPR mutagenesis which phenocopied previously reported morphants and mutants [19,20,23]. We next performed sequential injection of rbm24a morpholino and exogenous sox2 RNA to determine the extent to which sox2 can suppress the rbm24a-depleted microphthalmia. We found phenotypic suppression of the rbm24a-induced microphthalmia. We also performed a visual assay which tests for light/dark detection on these embryos and found partial visually functional rescue by sox2 RNA. This work highlights the study of post-transcriptional modification and their target RNAs during eye development.
2. Experimental Section
2.1. Animal Care
Zebrafish are maintained in standard conditions under the approval of the University of Iowa Institutional Animal Care and Use Committee (#8071513, 13 August 2018). Embryos are collected from natural spawning and raised between 28 and 30 °C. No more than 50 embryos are kept per 100 mm plate. Embryo plates are cleaned of dead daily and water changes are made as needed.
2.2. Microinjection
Embryos at the 1–2 cell stage were injected with either a translation-blocking rbm24a morpholino (0.6–1.2, 1–1.3, or 1.7–2 ng), splice-blocking rbm24a morpholino, or AltR CRISPR construct (3 nL). The morpholinos were ordered from Gene Tools. rbm24a AUG MO sequence: 5′-GCATCCTCACGAAACGCTCAAGTGC-3′. rbm24a SB MO sequence: 5′-TTGATATAATCCTCACCTGGCTGCA-3′. The AltR crRNA was ordered from IDT [24]. rbm24a AltR crRNA sequence: 5′-GGACUUUCCAGUCUGUCUGUGUUUUAGAGCUAUGCU-3′. RNA for rbm24a (100–400 pg), sox2 (200–300 pg), and EGFP (200–300 pg) was generated from cDNA that was cloned into the pCR8/GW/TOPO vector (rbm24a, sox2; Invitrogen, Carlsbad, CA, USA) or pCS2 + (EGFP, RZPD) before linearized templates were transcribed using the SP6 Ambion mMessage mMachine kit (Invitrogen, Carlsbad, CA, USA). rbm24a and sox2 RNAs have N-terminal Myc tags. sox2 RNA lacks a 3′ UTR. Microinjection needles were measured via a capillary tube to ensure dosages fell in the aforementioned ranges. Sequential injections were performed utilizing the same needle between morpholino-injected embryos (morpholino-only and morpholino with RNA) and RNA-injected embryos (RNA-only and morpholino with RNA) to ensure consistent dosage between experimental groups. An app was used to calculate the amount of construct injected (https://play.google.com/store/apps/details?id=com.canthonyscott.microinjectioncalc&hl=en_US). When noted, sequential injections were performed on a single day per set and involved the usage of the same clutch of eggs, needle, and dosage for all embryos injected.
2.3. Mutagenesis Detection
Uninjected and injected embryos underwent gDNA extraction at 2–4 dpf. Briefly, 20 µL of 50 mM NaOH per embryo was added before embryos were heated at 95 °C for 15 min. Samples were cooled and neutralized with 1 µL of 1 M Tris pH 8.0 per 10 µL of NaOH. The resulting gDNA underwent PCR amplification before being sequenced and insertion/deletion (indels) detection was performed utilizing Synthego ICE [25]. rbm24a-Forward-5′-ATGCATACCACGCAAAAGGAC-3′, rbm24a-Reverse-5′-CAGTCTGTCTGTCGGTAATCA-3′.
2.4. Automated Startle Response
The automated vision startle response, VIZN, was performed on 5 or 6 days post-fertilization (dpf) larvae as previously described [26]. Phenotypically normal larvae were first tested for the ability to swim by being prodded to ensure touch responsiveness and swimming ability, before being sorted and placed in 48-well plates.
2.5. Statistical Analysis
Data in tables were analyzed statistically with Fisher’s exact test implement in R [27]. VIZN assays were statistically analyzed by Mann–Whitney.
3. Results
3.1. Mutation of rbm24a Leads to Microphthalmia and Cardiomyopathy
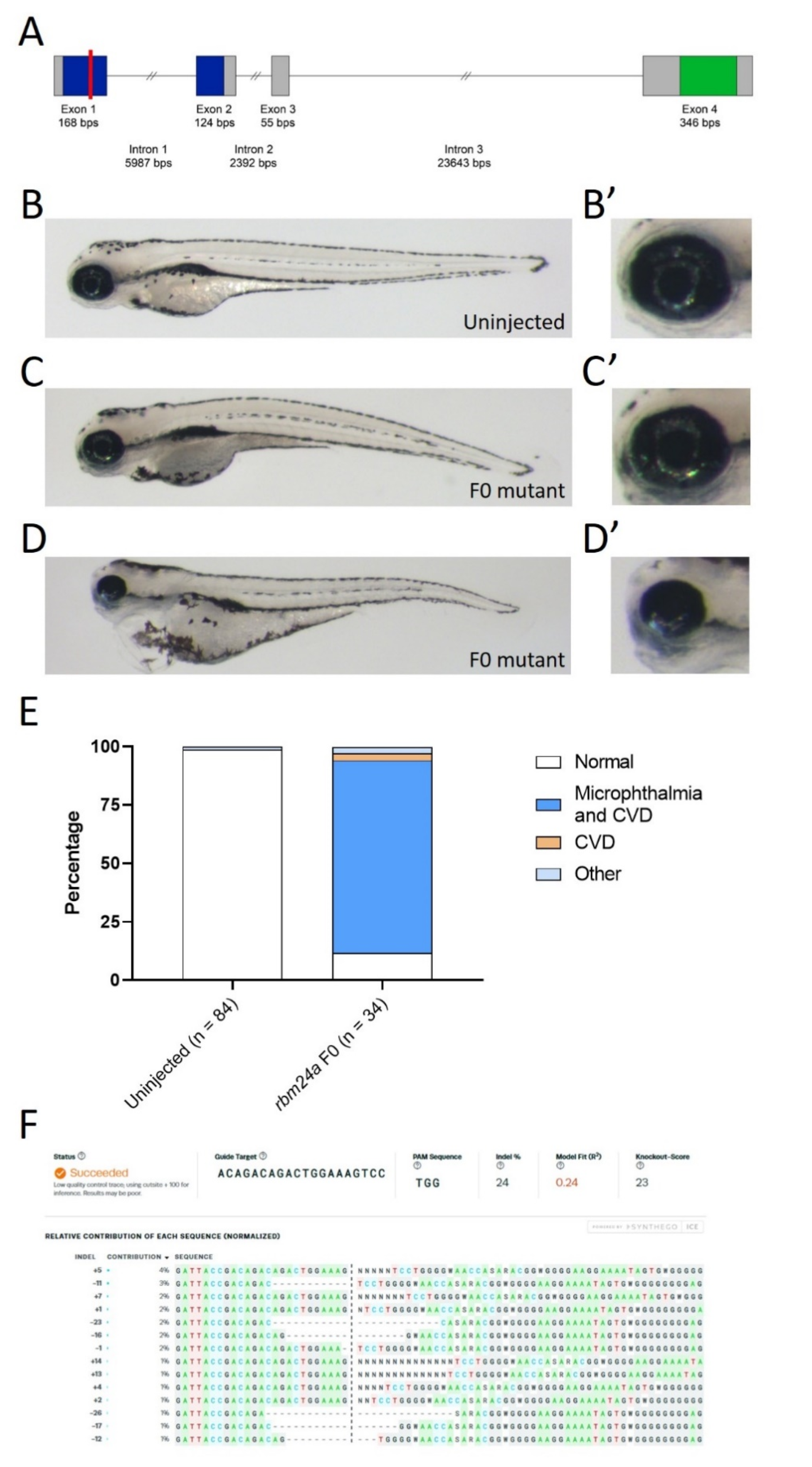
Zebrafish rbm24a is expressed in the developing heart, somites, and lens [19,20,22]. Translation and splice-blocking morpholino knockdown leads to dose-dependent defects, with lower doses exhibiting microphthalmia, while higher doses display microphthalmia and cardiac defects leading to edema (Supplemental Figure S1). We previously analyzed the microphthalmia phenotype both morphologically and histologically [13]. To validate the usage of the morpholino, we aimed to generate an rbm24a mutant. Similar to other organisms, including mice and humans, the zebrafish Rbm24a contains two domains: the RNA recognition motif (RRM) and an Alanine-rich region. The RRM domain spans exons 1 and 2 and is the domain responsible for binding to target RNAs. This domain is highly conserved, with 96.2% identity shared between zebrafish and humans. The Alanine-rich region is in exon 4 and has an unknown function in Rbm24a (Figure 1). Due to the key role of the RRM domain, we designed a CRISPR site in exon 1 which was a predicted null mutation (Figure 1). We performed CRISPR knockout and found phenotypes similar to those of morpholino knockdown. Most CRISPR-injected F0 embryos displayed microphthalmia (small eye(s)) and cardiac edema (fluid around the heart) which is consistent with the expression pattern of rbm24a, previously reported morphant phenotypes, and previously reported mutant phenotypes (Figure 1B–E) [19,20,23]. We extracted genomic DNA (gDNA) from both uninjected and CRISPR-injected F0, amplified the region flanking the target cut site, and sequenced the products. We next used Synthego ICE to determine the nature and frequency of indels [25]. Analysis of the CRISPR-injected F0 mutants showed an indel frequency between 20% and 24% when compared to the uninjected control embryos (Figure 1F). There is a statistically significant difference between the phenotypes of uninjected control and CRISPR-injected F0 embryos (p < 1 × 10−16) (Table 1).
3.2. rbm24a RNA Suppression of rbm24a Morpholino Knockdown
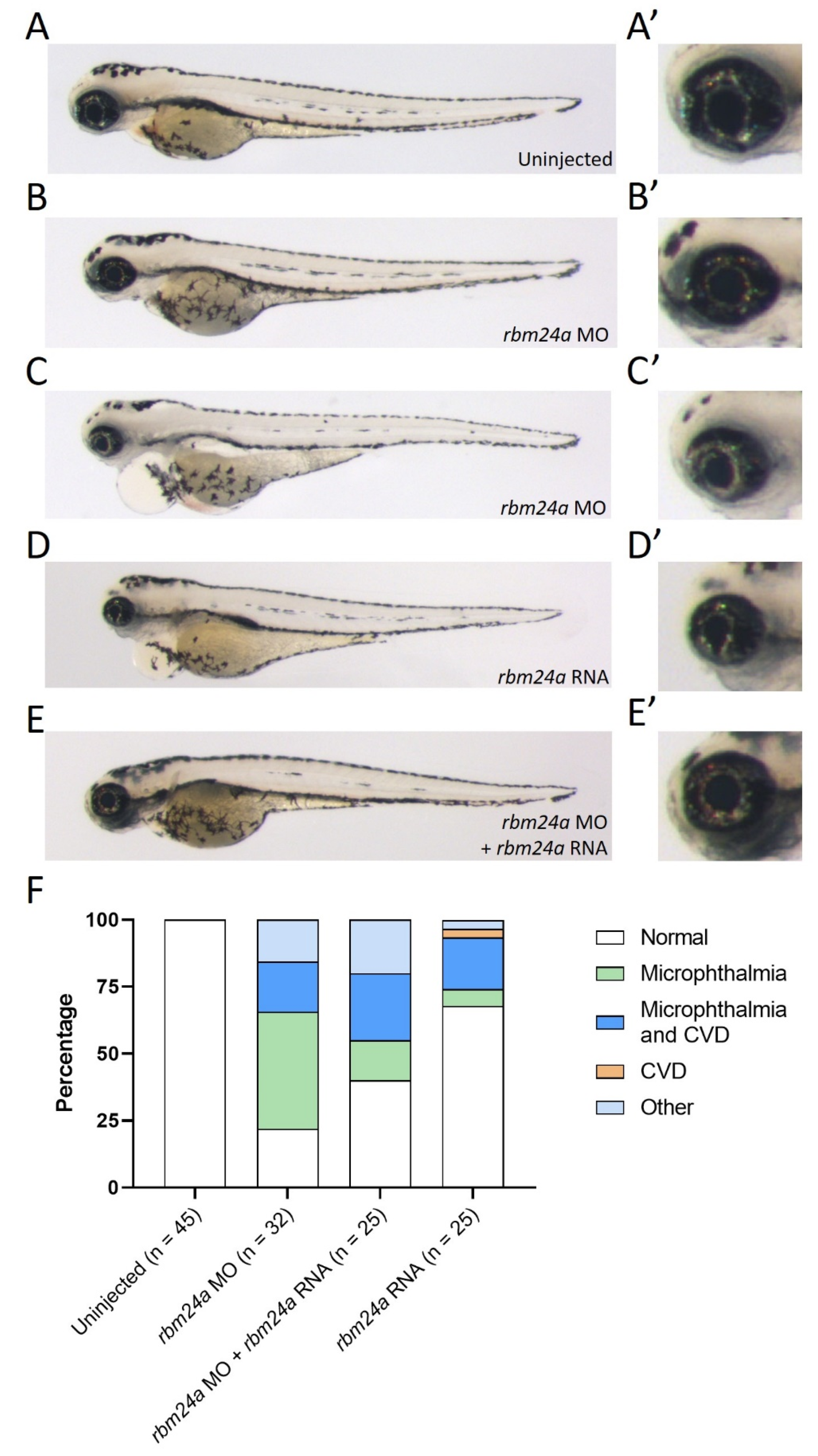
Due to the high number of CRISPR-injected F0 embryos exhibiting both microphthalmia and cardiac edema, we chose to utilize a low dose of translation-blocking morpholino in order to generate microphthalmia-only rbm24a knockdown phenotypes. When compared to uninjected embryos, morphant embryos injected with a low dose (1–1.3 ng) displayed only microphthalmia, while embryos injected at a higher dose (1.7–2 ng) phenocopied the CRISPR-injected F0 embryos with both microphthalmia and cardiac edema (Figure 2A–C’).
As an additional confirmation of the rbm24a knockdown phenotype, we next determined the extent to which exogenous rbm24a RNA can suppress the knockdown defects. We modified the rbm24a RNA construct so the morpholino could not bind to this exogenous rbm24a RNA. When injected alone, rbm24a RNA led to an overexpression phenotype very similar to the rbm24a knockdown phenotype (100–400 pg; Figure 2D–D’). The similarity of the rbm24a knockdown and overexpression phenotypes poses a challenge for phenotypic suppression studies, yet when we performed sequential injections of rbm24a morpholino and rbm24a RNA, we demonstrated phenotypic suppression (Figure 2E–E’). In morpholino knockdown alone, approximately 20% of the embryos were phenotypically normal, while the remaining 80% of embryos displayed defects. When embryos were sequentially injected with rbm24a morpholino and rbm24a RNA, the number with normal morphology doubled to nearly 40%, resulting in partial suppression of the knockdown microphthalmia defect (p = 0.0745) (Figure 2F, Table 2). The overlapping phenotypes generated by two different morpholinos, the suppression of the knockdown phenotype by exogenous rbm24a RNA, and the phenotypic similarity between the CRISPR-injected and morphant embryos indicate that the phenotypes observed are specific to rbm24a.
3.3. EGFP RNA Does Not Suppress rbm24a Morpholino Knockdown Phenotypes
Rbm24a is an RNA-binding protein. Therefore, we sought to determine if any exogenous RNA was sufficient to suppress the rbm24a knockdown phenotype. We utilized EGFP RNA as a control for RNA injection. EGFP is not naturally found in zebrafish and should have no function. Additionally, we can check for successful injection by screening embryos for EGFP fluorescence (Figure 3A). When compared to uninjected control embryos, both embryos injected with rbm24a morpholino (1.7–2 ng) and embryos injected with both rbm24a morpholino and EGFP RNA (200–300 pg) showed similar phenotypes, including many with microphthalmia and microphthalmia with cardiac edema (Figure 3B–D’). Injection of EGFP RNA alone yielded a phenotype similar to the uninjected control group (Figure 3E–E’). The number of normal embryos in both the uninjected group and EGFP RNA group is almost identical (p = 0.1050), while the number and nature of the affected embryos in both the rbm24a knockdown and rbm24a knockdown with EGFP RNA are also very similar (p = 0.5884) (Figure 3F, Table 3). Taken together, these data suggest that rbm24a knockdown defects cannot be suppressed by sequential injection of an unrelated exogenous RNA.
3.4. sox2 RNA Phenotypically Suppresses rbm24a Morpholino-Induced Microphthalmia
We previously identified sox2 as a target of Rbm24a in both zebrafish and mouse models [12]. In that study, we demonstrated that loss of rbm24a led to decreased levels of sox2. We hypothesize that supplying exogenous sox2 RNA would supplement the reduced endogenous sox2 RNA levels. We previously identified the binding site in the 3′ UTR of Sox2 for RBM24 via mouse cell culture studies [12]. Thus, we generated a zebrafish sox2 RNA construct which lacked the 3′ UTR to prevent Rbm24a binding (referred to as sox2 RNA). To test the functional role of sox2 as a target of Rbm24a, we performed sequential injection of rbm24a morpholino followed by sox2 RNA. Knockdown of rbm24a (1.7–2 ng) generated embryos with microphthalmia or microphthalmia with cardiac edema when compared against the uninjected control (Figure 4A–B’).
In contrast, embryos sequentially injected with both rbm24a morpholino and sox2 RNA (200–300 pg) were often normal in morphology (Figure 4C–C’). Injection of sox2 RNA alone also generated mostly phenotypically normal embryos (Figure 4D–D’). A majority of uninjected, rbm24a morpholino and sox2 RNA-injected, and sox2 RNA-injected embryos were morphologically normal with some exceptions in both injection groups. The rbm24a knockdown embryos displayed more embryos with microphthalmia or microphthalmia with cardiac edema when compared to the rbm24a knockdown with sox2 RNA, indicating that injection of exogenous sox2 RNA can partially suppress the rbm24a knockdown phenotype (Figure 4E, Table 4).
3.5. EGFP RNA Does Not Functionally Suppress rbm24a-Induced Visual Defects
We next wanted to determine the functional role of rbm24a in vision. Previously, we demonstrated that morpholino knockdown of rbm24a impacts the startle response in a dose-dependent manner [13]. When 5 to 6 days post-fertilization zebrafish larvae are exposed to an interruption of a constant light source, they perform a characteristic escape response which can be tracked with motion detection cameras and software. We utilized the automated startle response assay (VIZN) which generates five interruptions in light and records the zebrafish movement in response to the interruption in light [26]. Uninjected control larvae responded roughly four out of five times, indicating that they are visually responsive to the stark change in light in the startle response assay. Our previous studies indicated that low-dose knockdown of rbm24a maintained light detection [13]. For this study, we utilized a mid-range dose of rbm24a morpholino to better evaluate the effectiveness of RNA suppression. For all of our vision assays, we only utilized larvae that were touch responsive with a characteristic swimming response and had no cardiac edema. These rbm24a morphant larvae responded to the automated startle response assay (VIZN) significantly fewer times (approximately two out of five times) than the uninjected control (Figure 5). When sequentially injected with both rbm24a morpholino and EGFP RNA, larvae had a similar response to the rbm24a morpholino-alone larvae and responded about two out of five times. The difference between the rbm24a morpholino-only and rbm24a morpholino with EGFP RNA was not significantly different. The larvae injected with only EGFP RNA responded roughly four out of five times. There was no significant difference between the uninjected control and EGFP RNA-alone groups (Figure 5).
3.6. sox2 RNA Partially Functionally Suppresses rbm24a-Induced Visual Defects
We next investigated the extent to which sox2 RNA can restore visual functionality to rbm24a knockdown larvae. We performed VIZN on the uninjected control and rbm24a knockdown larvae. Uninjected control larvae responded approximately four out of five times, indicating that they are visually responsive to the startle response assay (Figure 6). As stated previously, we purposely utilized a dose of rbm24a morpholino which resulted in larvae responding statistically significantly fewer times (roughly two out of five) than the uninjected control. Sequential injection of rbm24a morpholino and sox2 RNA larvae responded significantly more times (about three out of five) than rbm24a morpholino-injected larvae. However, the rbm24a morpholino and sox2 RNA larvae were also statistically significantly different from the uninjected control larvae. Larvae injected with only sox2 RNA responded similarly (about four out of five times) to the uninjected control (Figure 6). Taken together, these data indicate sox2 partially restores visual function in rbm24a morpholino knockdown larvae.
4. Discussion
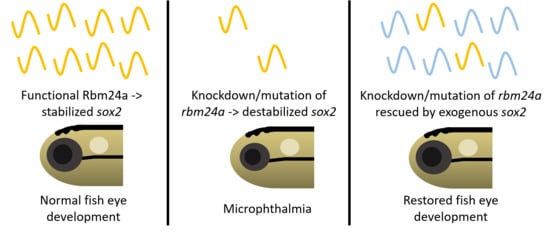
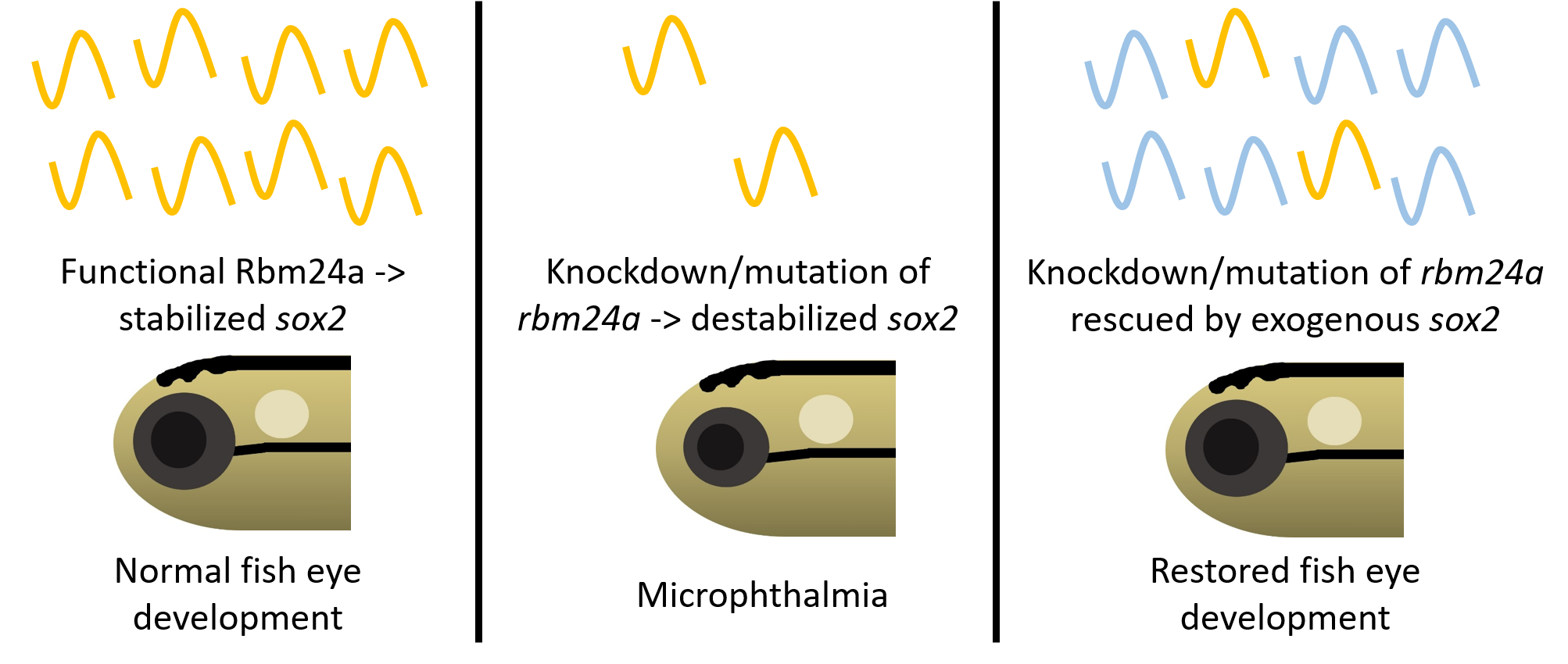
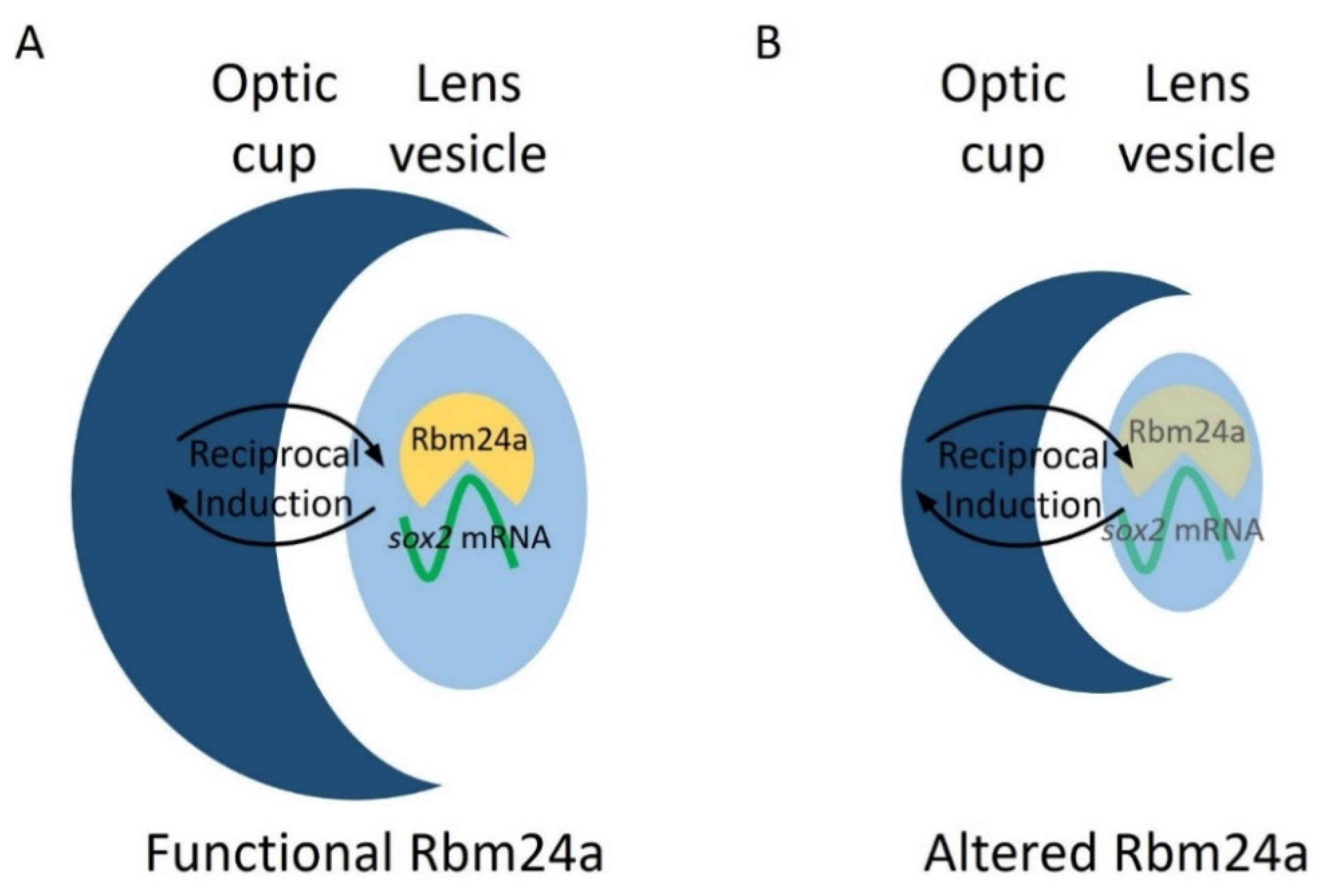
Our previous research indicated sox2 as a target for Rbm24a and suggests that Rbm24a binds to and stabilizes the sox2 mRNA transcript [12]. Localization data indicate that rbm24a is expressed exclusively in the lens. This suggests that Rbm24a binds to and regulates lens-expressed sox2 in the lens [17,18,19,20,21,28]. When knocked down via morpholino or mutated via CRISPR, the levels of Rbm24a are depleted which leads to a decrease in the stability and amount of sox2 mRNA. One possibility is that the reduced levels of sox2, a proliferative factor, lead to decreased lens vesicle size. Due to coordinated development between the lens vesicle and optic cup via reciprocal induction, the optic cup develops in tandem with the lens vesicle and results in microphthalmia (Figure 7).
Recently, a zebrafish TALEN mutant for rbm24a has been published which phenocopies both our morpholino knockdown and CRISPR-injected F0 defects [23]. In the Shao et al. 2020 study, the authors suggest that microphthalmia is the result of a cardiac morphological defect leading to poor blood flow to the eye. We hypothesized that the microphthalmia was due to target RNA disruption by decreased rbm24a expression. In our previously published studies, we identified that RBM24 binds to Sox2 in mouse cell cultures [12]. From this, we hypothesized that knockdown of rbm24a with the addition of exogenous sox2 would suppress the microphthalmia phenotype. We found that sequential knockdown of rbm24a and the addition of exogenous sox2 RNA generated a phenotype more similar to that of uninjected control embryos than rbm24a knockdown (Figure 4A–C’). Our data indicate sox2 is a main contributor to the rbm24a-induced microphthalmia. In terms of the previous study by Shao et al., it is possible that the TALEN mutant was not a null mutation which could have allowed for some function in the lens. Additionally, the utilization of heterotypic parabiosis allows for the potential transfer of more than just blood (including hormones, signaling molecules, etc.) between embryos. It is possible that the transfer of these biomolecules could have played a role in suppressing the microphthalmia phenotype [23]. Follow-up studies utilizing an organ-specific driver would resolve this contrast.
We occasionally observed cardiovascular defects in the sox2 overexpression embryos, perhaps stemming from the ubiquitous overexpression of sox2. Ubiquitously expressed exogenous sox2 may be able to substitute for cardiac members of the SOX family including Sox6 which has been shown to be a cardiomyocyte regulator in murine cells, Sox9 which is involved in heart valve development in mice, and Sox17 which has been shown to be essential for specification of cardiac mesoderm in mouse cell cultures [29,30,31]. In the future, an eye-specific promoter could be utilized to allow for the study of rbm24a/sox2 interactions with specificity to the eye.
In this study, we demonstrated that knockdown and mutation of the RNA-binding motif protein 24a gene, rbm24a, leads to microphthalmia at low-dose knockdown and microphthalmia with cardiac edema at increased knockdown doses. Additionally, we demonstrated that the microphthalmia induced by rbm24a functionally impacted the visual capabilities of the larvae, as knockdown fish performed worse in a visual behavior study. Previously, we identified sox2 as a target of Rbm24a [12]. When sequentially injected, exogenous sox2 RNA is able to phenotypically and partially functionally suppress the rbm24a knockdown larvae. This shows functional validation of a suspected target RNA by an RNA-binding protein. This work broadly demonstrates the key role RNA-binding proteins and post-transcriptional modification plays during development. It also highlights that genes not associated with disease, such as rbm24a, can impact a well-studied disease-associated gene (sox2), making it critical to better understand post-transcriptional regulation and the potential impacts of RNA-binding proteins and their associated target RNAs.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2227-9059/9/2/100/s1, Figure S1: Knockdown of rbm24a yields dose-dependent phenotypes at 4 days post-fertilization.
Author Contributions
Conceptualization, L.K.B. and D.C.S.; methodology, L.K.B.; investigation, L.K.B.; resources, C.A.S.; statistical analysis, L.K.B. and K.W.; writing—original draft preparation, L.K.B. and D.C.S.; writing—review and editing, L.K.B., C.A.S., and D.C.S.; project administration, D.C.S.; funding acquisition, D.C.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the American Heart Association, grant number 17GRNT33670684.
Institutional Review Board Statement
Zebrafish are maintained in standard conditions under the approval of the University of Iowa Institutional Animal Care and Use Committee (#8071513, 13 August 2018).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable.
Acknowledgments
The authors would like to thank Douglas Houston, Sheila Baker, Michael Dailey, and Bryan Phillips for their suggestions and expertise.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Johnson, G.J.; Minassian, D.C.; Weale, R.A.; West, S.K. The Epidemiology of Eye Disease; Arnold: London, UK, 2003. [Google Scholar]
- Graw, J. The genetic and molecular basis of congenital eye defects. Nat. Rev. Genet. 2003, 4, 876–888. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.P.; Taylor, A.E.; Sowden, J.C.; Ragge, N.K.; Russell-Eggitt, I.; Rahi, J.S.; Gilbert, C.E. Anophthalmos, microphthalmos, and typical coloboma in the United Kingdom: A prospective study of incidence and risk. Investig. Ophthalmol. Vis. Sci. 2011, 52, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Fry, M.; Al-Samarraie, M.; Gilbert, C.; Steinkuller, P.G. An update on progress and the changing epidemiology of causes of childhood blindness worldwide. J. Am. Assoc. Pediatric Ophthalmol. Strabismus 2012, 16, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Reis, L.M.; Semina, E.V. Conserved genetic pathways associated with microphthalmia, anophthalmia, and coloboma. Birth Defects Res. Part C Embryo Today Rev. 2015, 105, 96–113. [Google Scholar] [CrossRef] [Green Version]
- Fantes, J.; Ragge, N.K.; Lynch, S.-A.; McGill, N.I.; Collin, J.R.O.; Howard-Peebles, P.N.; Hayward, C.; Vivian, A.J.; Williamson, K.; van Heyningen, V. Mutations in SOX2 cause anophthalmia. Nat. Genet. 2003, 33, 462–463. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Chipperfield, H.; Melton, D.A.; Wong, W.H. A gene regulatory network in mouse embryonic stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 16438–16443. [Google Scholar] [CrossRef] [Green Version]
- Kondoh, H.; Lovell-Badge, R. Sox2: Biology and Role in Development and Disease; Academic Press: London, UK, 2015. [Google Scholar]
- Wu, J.; Belmonte, J.C.I. The molecular harbingers of early mammalian embryo patterning. Cell 2016, 165, 13–15. [Google Scholar] [CrossRef]
- Lachke, S.A.; Ho, J.W.; Kryukov, G.V.; O’Connell, D.J.; Aboukhalil, A.; Bulyk, M.L.; Park, P.J.; Maas, R.L. iSyTE: Integrated Systems Tool for Eye gene discovery. Investig. Ophthalmol. Vis. Sci. 2012, 53, 1617–1627. [Google Scholar] [CrossRef] [Green Version]
- Dash, S.; Brastrom, L.K.; Patel, S.D.; Scott, C.A.; Slusarski, D.C.; Lachke, S.A. The master transcription factor SOX2, mutated in anophthalmia/microphthalmia, is post-transcriptionally regulated by the conserved RNA-binding protein RBM24 in vertebrate eye development. Hum. Mol. Genet. 2020, 29, 591–604. [Google Scholar] [CrossRef]
- Brastrom, L.K.; Scott, C.A.; Dawson, D.V.; Slusarski, D.C. A high-throughput assay for congenital and age-related eye diseases in zebrafish. Biomedicines 2019, 7, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Hung, L.-H.; Licht, T.; Kostin, S.; Looso, M.; Khrameeva, E.; Bindereif, A.; Schneider, A.; Braun, T. RBM24 is a major regulator of muscle-specific alternative splicing. Dev. Cell 2014, 31, 87–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Zhang, Y.; Xu, E.; Mohibi, S.; de Anda, D.M.; Jiang, Y.; Zhang, J.; Chen, X. Rbm24, a target of p53, is necessary for proper expression of p53 and heart development. Cell Death Differ. 2018, 25, 1118–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Kong, X.; Zhang, M.; Yang, X.; Xu, X. RNA binding protein 24 deletion disrupts global alternative splicing and causes dilated cardiomyopathy. Protein Cell 2019, 10, 405–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fetka, I.; Radeghieri, A.; Bouwmeester, T. Expression of the RNA recognition motif-containing protein SEB-4 during Xenopus embryonic development. Mech. Dev. 2000, 94, 283–286. [Google Scholar] [CrossRef]
- Oberleitner, S. Seb4—An RNA-Binding Protein as a Novel Regulator of Myogenesis during Early Development in Xenopus laevis. Ph.D. Dissertation, Ludwig Maximilian University of Munich, Munich, Germany, 2008. [Google Scholar]
- Maragh, S.; Miller, R.A.; Bessling, S.L.; McGaughey, D.M.; Wessels, M.W.; De Graaf, B.; Stone, E.A.; Bertoli-Avella, A.M.; Gearhart, J.D.; Fisher, S. Identification of RNA binding motif proteins essential for cardiovascular development. BMC Dev. Biol. 2011, 11, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Maragh, S.; Miller, R.A.; Bessling, S.L.; Wang, G.; Hook, P.W.; McCallion, A.S. Rbm24a and Rbm24b are required for normal somitogenesis. PLoS ONE 2014, 9, e105460. [Google Scholar] [CrossRef] [Green Version]
- Grifone, R.; Saquet, A.; Xu, Z.; Shi, D.L. Expression patterns of Rbm24 in lens, nasal epithelium, and inner ear during mouse embryonic development. Dev. Dyn. 2018, 247, 1160–1169. [Google Scholar] [CrossRef] [Green Version]
- Wagner, D.E.; Weinreb, C.; Collins, Z.M.; Briggs, J.A.; Megason, S.G.; Klein, A.M. Single-cell mapping of gene expression landscapes and lineage in the zebrafish embryo. Science 2018, 360, 981–987. [Google Scholar] [CrossRef] [Green Version]
- Shao, M.; Lu, T.; Zhang, C.; Zhang, Y.-Z.; Kong, S.-H.; Shi, D.-L. Rbm24 controls poly (A) tail length and translation efficiency of crystallin mRNAs in the lens via cytoplasmic polyadenylation. Proc. Natl. Acad. Sci. USA 2020, 117, 7245–7254. [Google Scholar] [CrossRef] [Green Version]
- Vakulskas, C.A.; Dever, D.P.; Rettig, G.R.; Turk, R.; Jacobi, A.M.; Collingwood, M.A.; Bode, N.M.; McNeill, M.S.; Yan, S.; Camarena, J. A high-fidelity Cas9 mutant delivered as a ribonucleoprotein complex enables efficient gene editing in human hematopoietic stem and progenitor cells. Nat. Med. 2018, 24, 1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roginsky, J. Analyzing CRISPR Editing Results: Synthego Developed a Tool Called ICE to Be More Efficient Than Other Methods. Genet. Eng. Biotechnol. News 2018, 38, S24–S26. [Google Scholar] [CrossRef]
- Scott, C.A.; Marsden, A.N.; Slusarski, D.C. Automated, high-throughput, in vivo analysis of visual function using the zebrafish. Dev. Dyn. 2016, 245, 605–613. [Google Scholar] [CrossRef] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing; The R Foundation: Vienna, Austria, 2017. [Google Scholar]
- Poon, K.L.; Tan, K.T.; Wei, Y.Y.; Ng, C.P.; Colman, A.; Korzh, V.; Xu, X.Q. RNA-binding protein RBM24 is required for sarcomere assembly and heart contractility. Cardiovasc. Res. 2012, 94, 418–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen-Barak, O.; Yi, Z.; Hagiwara, N.; Monzen, K.; Komuro, I.; Brilliant, M.H. Sox6 regulation of cardiac myocyte development. Nucleic Acids Res. 2003, 31, 5941–5948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garside, V.C.; Cullum, R.; Alder, O.; Lu, D.Y.; Vander Werff, R.; Bilenky, M.; Zhao, Y.; Jones, S.J.; Marra, M.A.; Underhill, T.M. SOX9 modulates the expression of key transcription factors required for heart valve development. Development 2015, 142, 4340–4350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Asakura, M.; Inoue, H.; Nakamura, T.; Sano, M.; Niu, Z.; Chen, M.; Schwartz, R.J.; Schneider, M.D. Sox17 is essential for the specification of cardiac mesoderm in embryonic stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 3859–3864. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Mutation of rbm24a phenocopies morpholino knockdown. (A) Schematic for genetic structure of Rbm24a, with the RNA recognition motif (RRM) in blue and the Ala-rich region in green. The red line indicates the site of the Alt-R CRISPR/Cas0 mutation. (B) Uninjected control 4 dpf embryo with wild type morphology. (B’) Detail of eye shown in B. (C) and (D) are Alt-R CRISPR/Cas9-injected embryos showing the variation of the phenotypes. (C’) and (D’) are detail of eyes found in C and D, respectively. (E) Graph of uninjected and CRISPR-injected F0 embryos. (F) Synthego ICE analysis of mutations found in F0 mutants. Indels are listed. Images taken at 33×.
Figure 1.
Mutation of rbm24a phenocopies morpholino knockdown. (A) Schematic for genetic structure of Rbm24a, with the RNA recognition motif (RRM) in blue and the Ala-rich region in green. The red line indicates the site of the Alt-R CRISPR/Cas0 mutation. (B) Uninjected control 4 dpf embryo with wild type morphology. (B’) Detail of eye shown in B. (C) and (D) are Alt-R CRISPR/Cas9-injected embryos showing the variation of the phenotypes. (C’) and (D’) are detail of eyes found in C and D, respectively. (E) Graph of uninjected and CRISPR-injected F0 embryos. (F) Synthego ICE analysis of mutations found in F0 mutants. Indels are listed. Images taken at 33×.

Figure 2.
Suppression of rbm24a morpholino knockdown with rbm24a RNA. (A) Uninjected 4 dpf embryo with wild type morphology, (A’) detail of eye in A. (B) Knockdown of rbm24a at low doses leads to microphthalmia, (B’) detail of eye in B, while (C) is a higher dose showing microphthalmia with cardiac edema, (C’) detail of eye in C. (D) Injection of rbm24a RNA yields phenotypes similar to higher dose knockdown, (D’) detail of eye in D. (E) Sequential injection of rbm24a morpholino and rbm24a RNA suppresses the phenotypes, (E’) detail of eye in E. (F) Graph of phenotypes. Images taken at 33×.
Figure 2.
Suppression of rbm24a morpholino knockdown with rbm24a RNA. (A) Uninjected 4 dpf embryo with wild type morphology, (A’) detail of eye in A. (B) Knockdown of rbm24a at low doses leads to microphthalmia, (B’) detail of eye in B, while (C) is a higher dose showing microphthalmia with cardiac edema, (C’) detail of eye in C. (D) Injection of rbm24a RNA yields phenotypes similar to higher dose knockdown, (D’) detail of eye in D. (E) Sequential injection of rbm24a morpholino and rbm24a RNA suppresses the phenotypes, (E’) detail of eye in E. (F) Graph of phenotypes. Images taken at 33×.

Figure 3.
EGFP RNA does not suppress rbm24a knockdown. (A) Uninjected and EGFP RNA-injected 1 dpf embryos are shown in both brightfield (top) and with a GFP filter (bottom). (B) Uninjected 4 dpf embryo with wild type morphology, (B’) detail of eye in B. (C) rbm24a knockdown embryos display microphthalmia with cardiac edema, (C’) detail of eye in C. (D) Sequential injection of rbm24a morpholino and EGFP RNA yields phenotypes similar to knockdown alone, (D’) detail of eye in D. (E) EGFP RNA-injected embryos are morphologically wild type, (E’) detail of eye in E. (F) Graph of phenotypes. Image A taken at 62×. Images B-E’ taken at 33×.
Figure 3.
EGFP RNA does not suppress rbm24a knockdown. (A) Uninjected and EGFP RNA-injected 1 dpf embryos are shown in both brightfield (top) and with a GFP filter (bottom). (B) Uninjected 4 dpf embryo with wild type morphology, (B’) detail of eye in B. (C) rbm24a knockdown embryos display microphthalmia with cardiac edema, (C’) detail of eye in C. (D) Sequential injection of rbm24a morpholino and EGFP RNA yields phenotypes similar to knockdown alone, (D’) detail of eye in D. (E) EGFP RNA-injected embryos are morphologically wild type, (E’) detail of eye in E. (F) Graph of phenotypes. Image A taken at 62×. Images B-E’ taken at 33×.

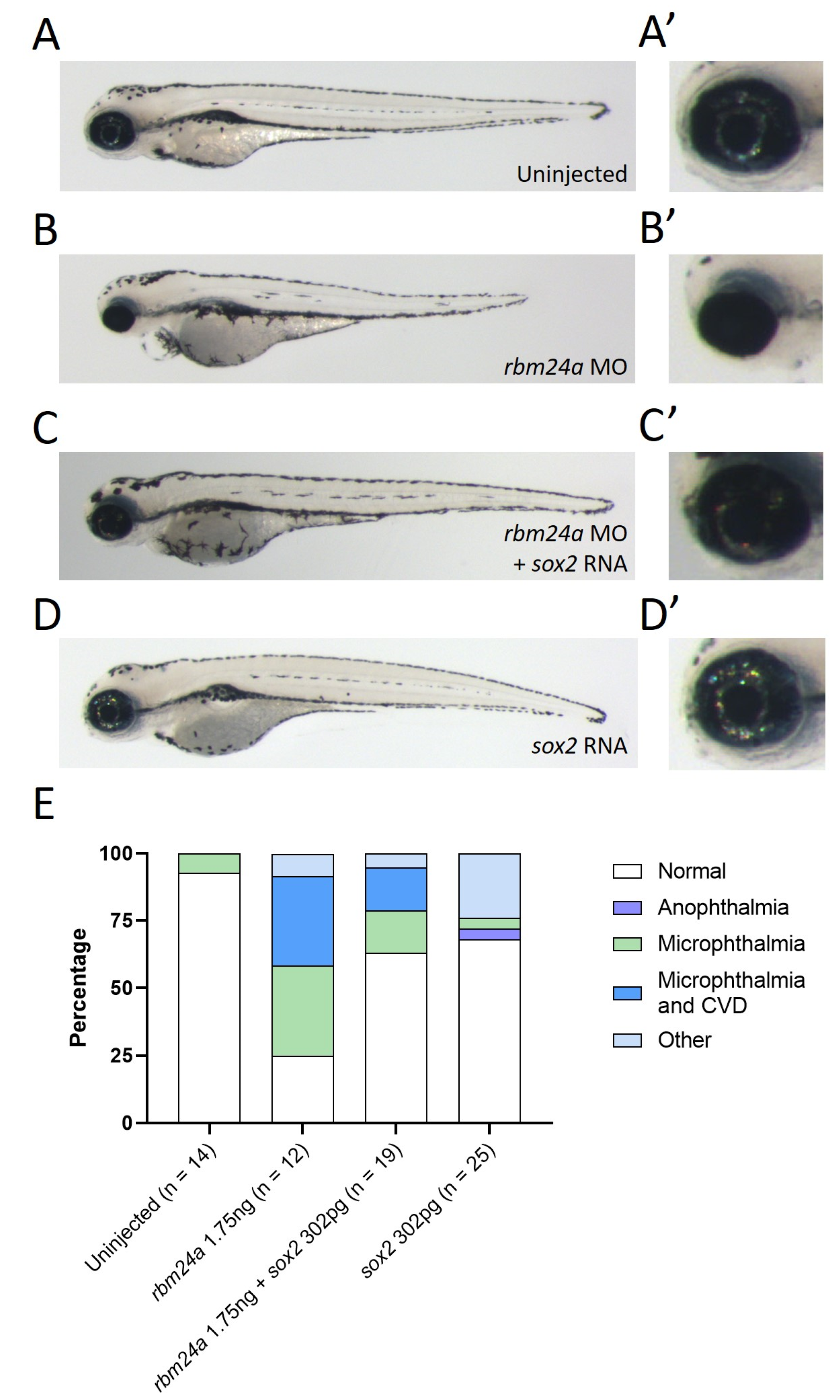
Figure 4.
Exogenous sox2 can suppress rbm24a-associated microphthalmia. (A) Uninjected 4 dpf embryo with wild type morphology, (A’) detail of eye in A. (B) Knockdown of rbm24a at low doses leads to microphthalmia, (B’) detail of eye in B. (C) Sequential injection of rbm24a morpholino and sox2 RNA results in a phenotype more similar to wild type than rbm24a morphant, (C’) detail of eye in C. (D) Injection of sox2 RNA alone results in wild type morphology, (D’) detail of eye in D. (E) Graph of phenotypes. Images taken at 33×.
Figure 4.
Exogenous sox2 can suppress rbm24a-associated microphthalmia. (A) Uninjected 4 dpf embryo with wild type morphology, (A’) detail of eye in A. (B) Knockdown of rbm24a at low doses leads to microphthalmia, (B’) detail of eye in B. (C) Sequential injection of rbm24a morpholino and sox2 RNA results in a phenotype more similar to wild type than rbm24a morphant, (C’) detail of eye in C. (D) Injection of sox2 RNA alone results in wild type morphology, (D’) detail of eye in D. (E) Graph of phenotypes. Images taken at 33×.

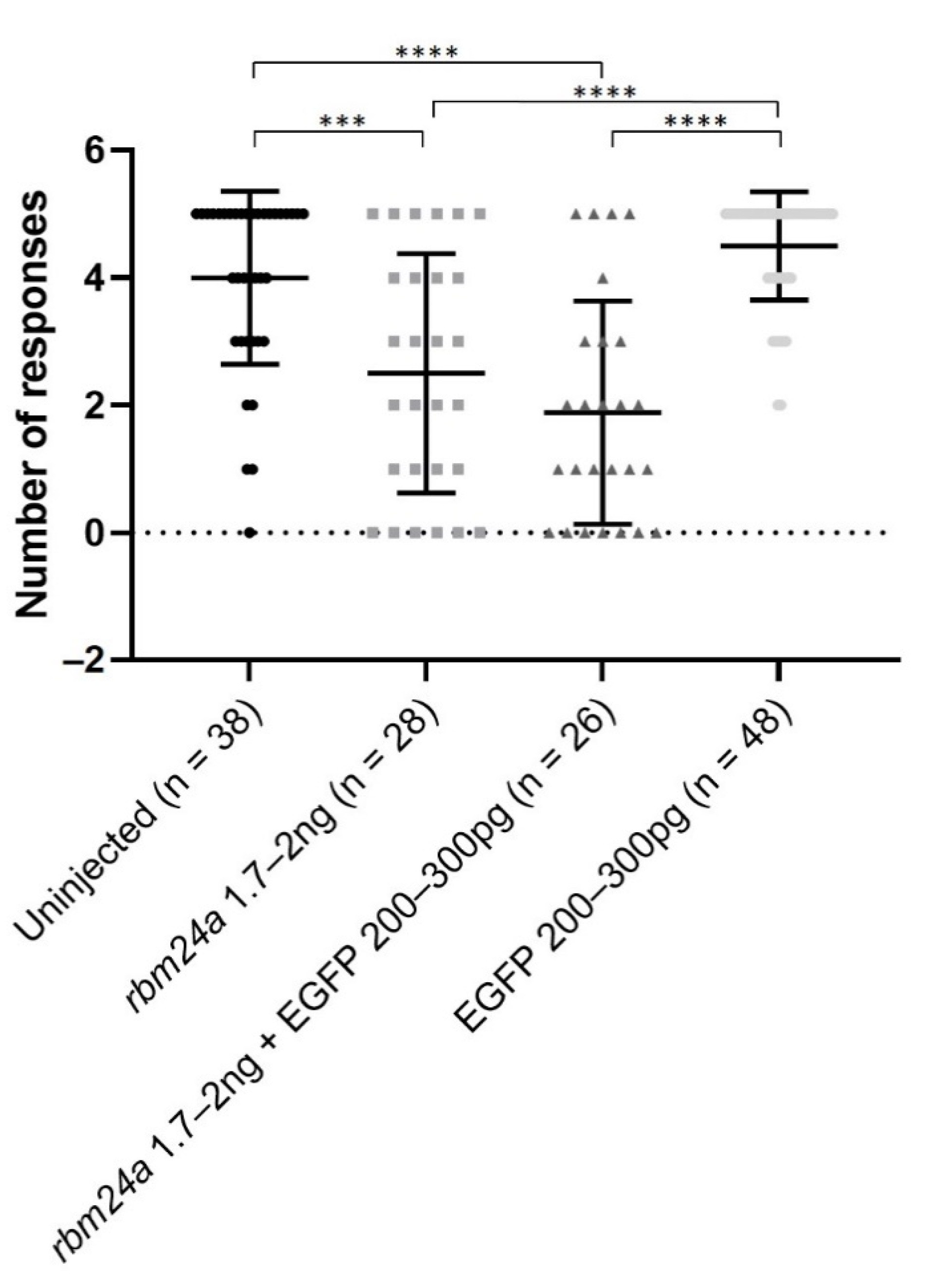
Figure 5.
EGFP RNA does not improve visual function of rbm24a knockdown embryos. Automated startle response assay (VIZN) was performed on uninjected, rbm24a knockdown, rbm24a knockdown with EGFP RNA, and EGFP RNA larvae. Knockdown of rbm24a inhibits visual function, which was statistically significant compared to uninjected. Addition of EGFP RNA to rbm24a knockdown does not statistically significantly increase visual function when compared to rbm24a knockdown alone. Injection of EGFP RNA alone does not statistically significantly alter visual function from uninjected. Mann–Whitney *** p < 0.001, **** p < 0.0001. Nonsignificant interactions not shown.
Figure 5.
EGFP RNA does not improve visual function of rbm24a knockdown embryos. Automated startle response assay (VIZN) was performed on uninjected, rbm24a knockdown, rbm24a knockdown with EGFP RNA, and EGFP RNA larvae. Knockdown of rbm24a inhibits visual function, which was statistically significant compared to uninjected. Addition of EGFP RNA to rbm24a knockdown does not statistically significantly increase visual function when compared to rbm24a knockdown alone. Injection of EGFP RNA alone does not statistically significantly alter visual function from uninjected. Mann–Whitney *** p < 0.001, **** p < 0.0001. Nonsignificant interactions not shown.

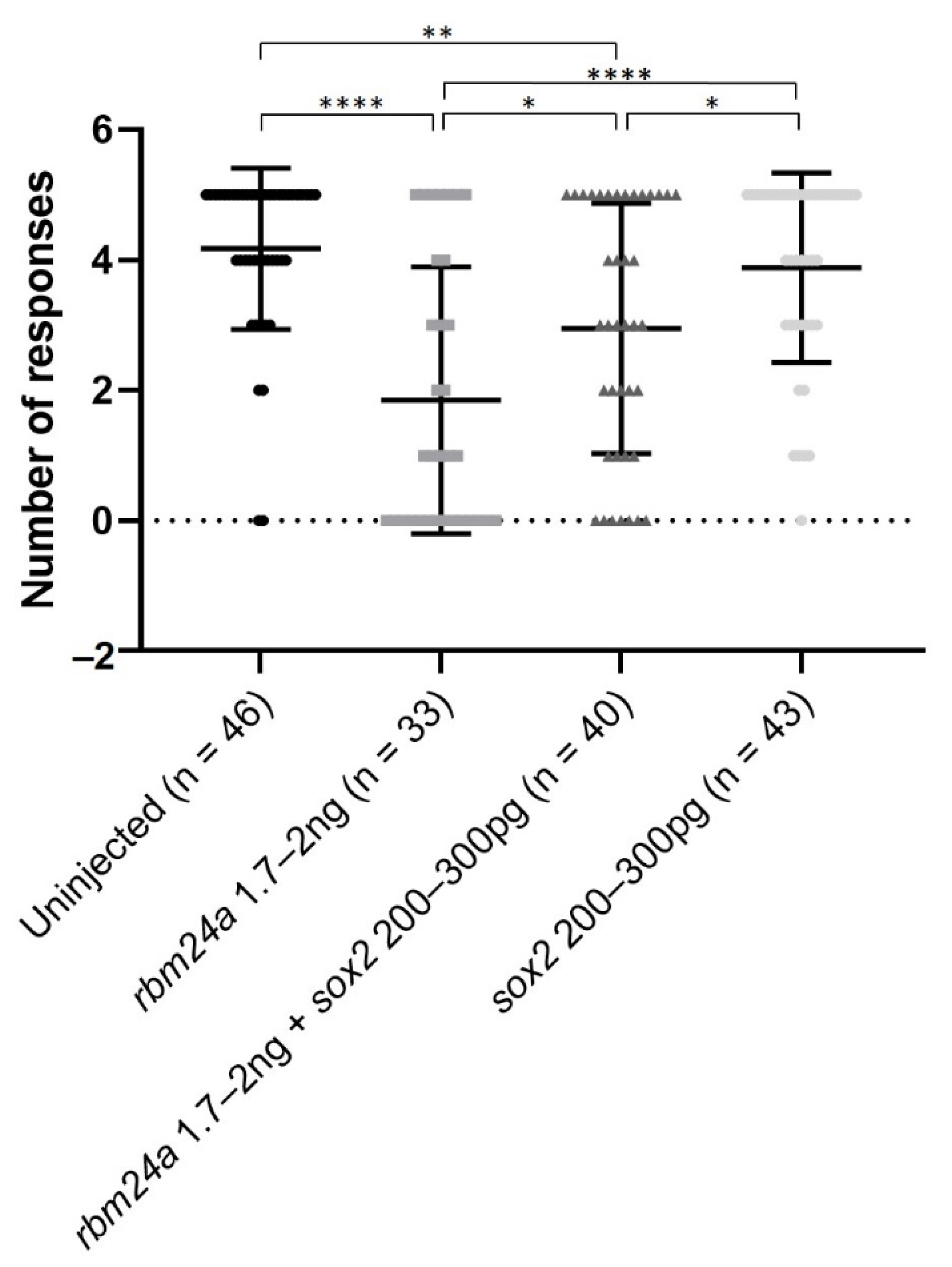
Figure 6.
Injection of sox2 RNA can partially restore visual function by VIZN to rbm24a knockdown embryos. VIZN was performed on uninjected, rbm24a knockdown, rbm24a knockdown with sox2 RNA, and sox2 RNA larvae. Knockdown of rbm24a statistically significantly inhibits visual function when compared to uninjected. Addition of sox2 RNA to rbm24a knockdown statistically significantly improves visual function when compared to rbm24a knockdown, but not to the same level as uninjected. Injection of sox2 RNA alone does not statistically significantly alter visual function from uninjected. Mann–Whitney * p < 0.05, ** p < 0.01, **** p < 0.0001. Nonsignificant interactions not shown.
Figure 6.
Injection of sox2 RNA can partially restore visual function by VIZN to rbm24a knockdown embryos. VIZN was performed on uninjected, rbm24a knockdown, rbm24a knockdown with sox2 RNA, and sox2 RNA larvae. Knockdown of rbm24a statistically significantly inhibits visual function when compared to uninjected. Addition of sox2 RNA to rbm24a knockdown statistically significantly improves visual function when compared to rbm24a knockdown, but not to the same level as uninjected. Injection of sox2 RNA alone does not statistically significantly alter visual function from uninjected. Mann–Whitney * p < 0.05, ** p < 0.01, **** p < 0.0001. Nonsignificant interactions not shown.

Figure 7.
Model for Rbm24a in zebrafish eye development. (A) Functional Rbm24a is found in the developing lens of zebrafish. There, the protein acts to bind to and stabilize sox2 mRNA. This interaction leads to the development of a normal sized lens and, with reciprocal induction signaling between the lens and retina, the retina also develops normally to size-match the lens. (B) With either knockdown or mutation of rbm24a, there is lessened Rbm24a protein in the lens which in turn cannot stabilize as many sox2 mRNA molecules (represented as transparent shapes). The lack of the correct amount of sox2, a proliferative factor, causes the lens to develop smaller than normal. However, reciprocal induction has not been affected, which leads to the lens and retina developing small in tandem.
Figure 7.
Model for Rbm24a in zebrafish eye development. (A) Functional Rbm24a is found in the developing lens of zebrafish. There, the protein acts to bind to and stabilize sox2 mRNA. This interaction leads to the development of a normal sized lens and, with reciprocal induction signaling between the lens and retina, the retina also develops normally to size-match the lens. (B) With either knockdown or mutation of rbm24a, there is lessened Rbm24a protein in the lens which in turn cannot stabilize as many sox2 mRNA molecules (represented as transparent shapes). The lack of the correct amount of sox2, a proliferative factor, causes the lens to develop smaller than normal. However, reciprocal induction has not been affected, which leads to the lens and retina developing small in tandem.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Number of phenotypes associated with uninjected control and CRISPR-injected F0 embryos.
| Uninjected | rbm24a CRISPR F0 | |
|---|---|---|
| Normal | 83 | 4 |
| Microphthalmia | 0 | |
| Microphthalmia and CVD 1 | 28 | |
| CVD | 1 | |
| Other 2 | 1 | 1 |
| Total | 84 | 34 |
1 CVD stands for cardiovascular defect. 2 Other is a category reserved for phenotypes that do not fall under the above categories. They are frequently trunk defects.
Table 2.
Number of phenotypes associated with uninjected, rbm24a knockdown, rbm24a knockdown with rbm24a RNA, and rbm24a RNA alone.
Table 2.
Number of phenotypes associated with uninjected, rbm24a knockdown, rbm24a knockdown with rbm24a RNA, and rbm24a RNA alone.
| Uninjected | rbm24a MO | rbm24a MO + rbm24a RNA | rbm24a RNA | |
|---|---|---|---|---|
| Normal | 45 | 7 | 10 | 21 |
| Microphthalmia | 14 | 4 | 2 | |
| Microphthalmia and CVD | 6 | 6 | 6 | |
| CVD | 1 | |||
| Other | 5 | 5 | 1 | |
| Total | 45 | 32 | 25 | 31 |
Table 3.
Number of phenotypes associated with uninjected, rbm24a knockdown, rbm24a knockdown with EGFP RNA, and EGFP RNA alone.
Table 3.
Number of phenotypes associated with uninjected, rbm24a knockdown, rbm24a knockdown with EGFP RNA, and EGFP RNA alone.
| Uninjected | rbm24a MO | rbm24a MO + EGFP RNA | EGFP RNA | |
|---|---|---|---|---|
| Normal | 63 | 2 | 1 | 28 |
| Microphthalmia | 2 | 6 | 10 | 2 |
| Microphthalmia and CVD | 13 | 6 | ||
| CVD | ||||
| Other | 4 | 9 | ||
| Total | 63 | 25 | 26 | 30 |
Table 4.
Number of phenotypes associated with uninjected, rbm24a knockdown, rbm24a knockdown with sox2 RNA, and sox2 RNA alone.
Table 4.
Number of phenotypes associated with uninjected, rbm24a knockdown, rbm24a knockdown with sox2 RNA, and sox2 RNA alone.
| Uninjected | rbm24a MO | rbm24a MO + sox2 RNA | sox2 RNA | |
|---|---|---|---|---|
| Normal | 13 | 3 | 12 | 17 |
| Microphthalmia | 2 | |||
| Microphthalmia and CVD | 4 | 3 | ||
| CVD | ||||
| Other | 1 | 5 | 4 | 6 |
| Total | 14 | 12 | 17 | 25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Brastrom, L.K.; Scott, C.A.; Wang, K.; Slusarski, D.C. Functional Role of the RNA-Binding Protein Rbm24a and Its Target sox2 in Microphthalmia. Biomedicines 2021, 9, 100. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9020100
AMA Style
Brastrom LK, Scott CA, Wang K, Slusarski DC. Functional Role of the RNA-Binding Protein Rbm24a and Its Target sox2 in Microphthalmia. Biomedicines. 2021; 9(2):100. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9020100
Chicago/Turabian StyleBrastrom, Lindy K., C. Anthony Scott, Kai Wang, and Diane C. Slusarski. 2021. "Functional Role of the RNA-Binding Protein Rbm24a and Its Target sox2 in Microphthalmia" Biomedicines 9, no. 2: 100. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9020100
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.