
Dysbiosis and Enhanced Beta-Defensin Production in Hair Follicles of Patients with Lichen Planopilaris and Frontal Fibrosing Alopecia

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Metagenomic Analysis
- Sequence quality control and feature table construction: basic sequencing quality and further noise removal were achieved using the DADA2 QIIME2 plugins;
- Tree generation for phylogenetic diversity analyses: a rooted phylogenetic tree was generated using the mafft program to perform a multiple sequence alignment. The alignment was filtered to remove highly variable positions. FastTree was then applied to generate a phylogenetic tree from the masked alignment. Finally, midpoint rooting was applied to place the root of the tree;
- Taxonomic analysis: a pre-trained naive Bayes classifier was used to explore the taxonomic distribution of the samples. This classifier was trained on the SILVA (release v132, 2017) 99% 16S rRNA Operational Taxonomic Units (OTUs), on full length sequences of the 16S gene [31];
- Alpha rarefaction plotting: alpha diversity was explored as a function of sampling depth using the QIIME2 diversity alpha-rarefaction visualizer;
- Count table generation: finally, the results in BIOM format generated by QIIME2 were processed into count tables for further statistical analysis and processing.
2.3. Statistical Analysis
2.4. Histological Assessment
3. Results
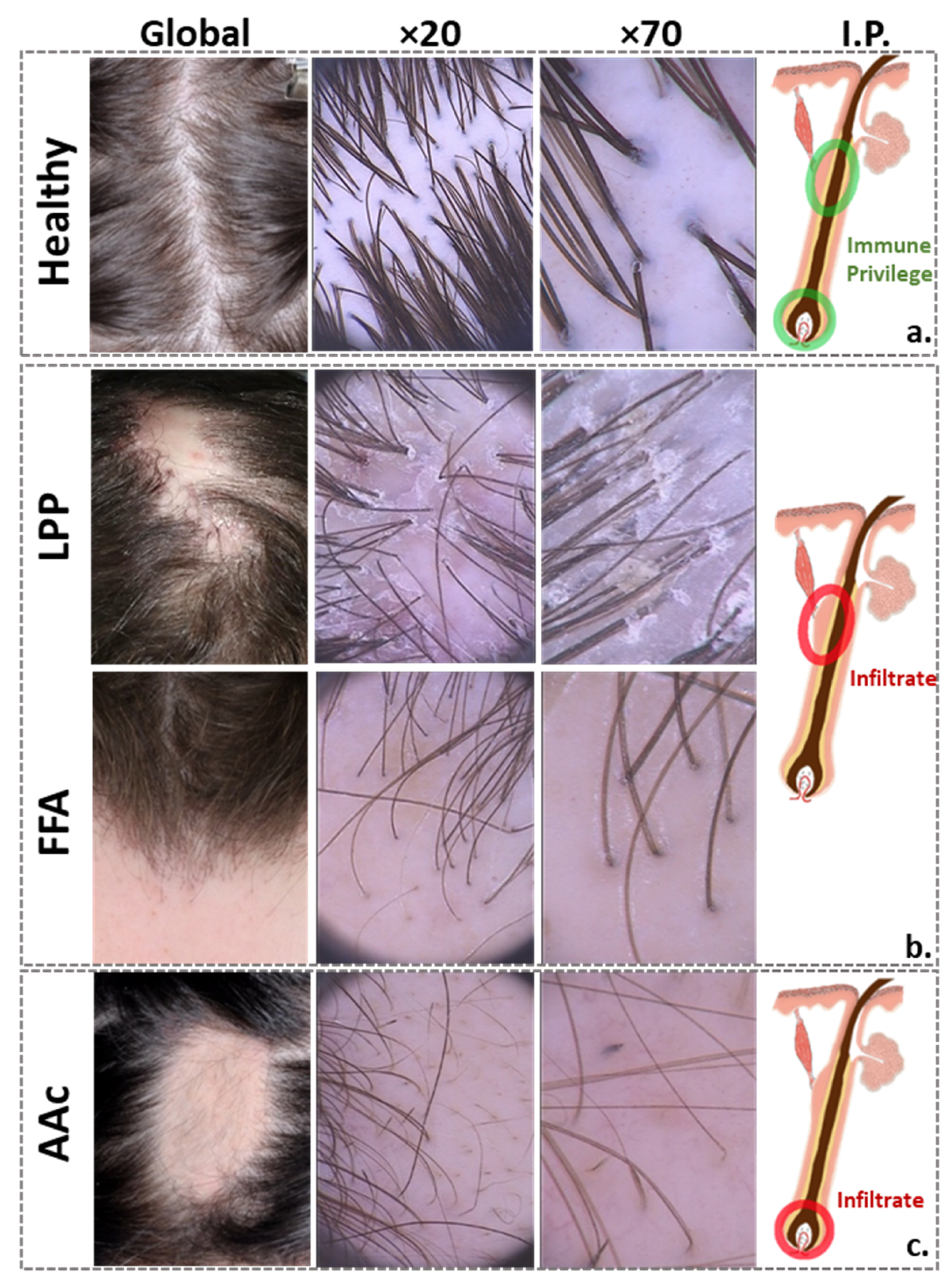
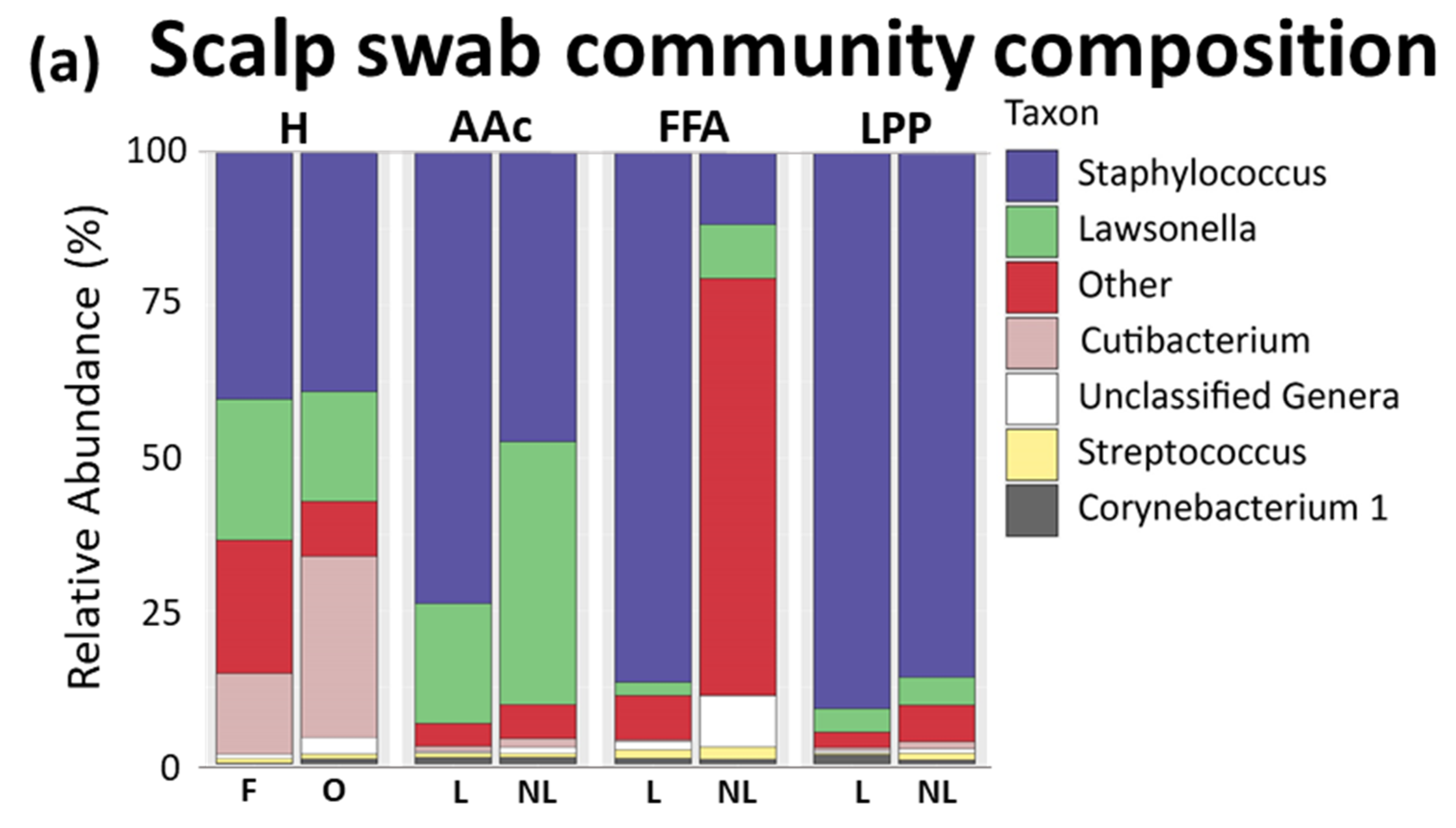
3.1. Scalp Surface
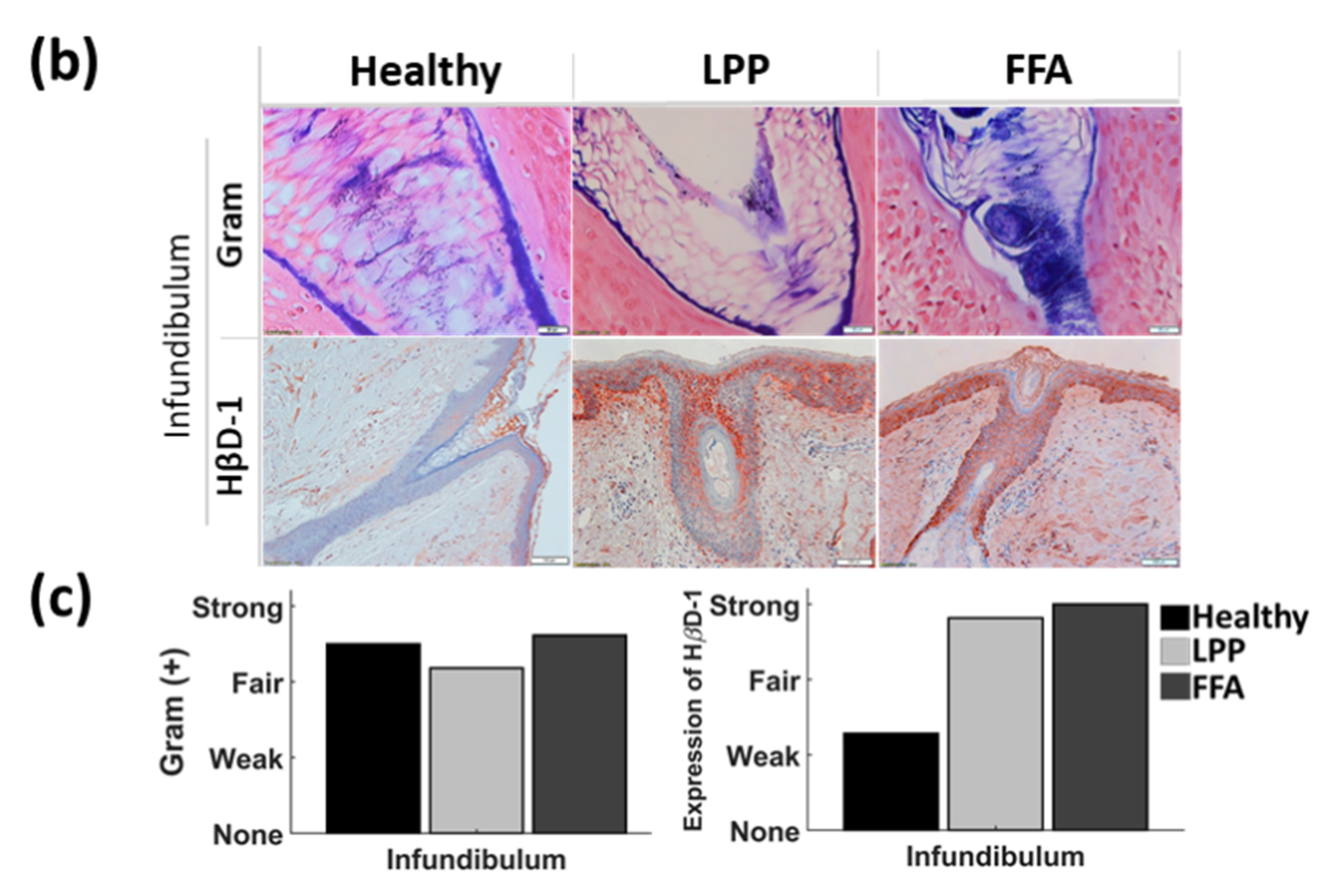
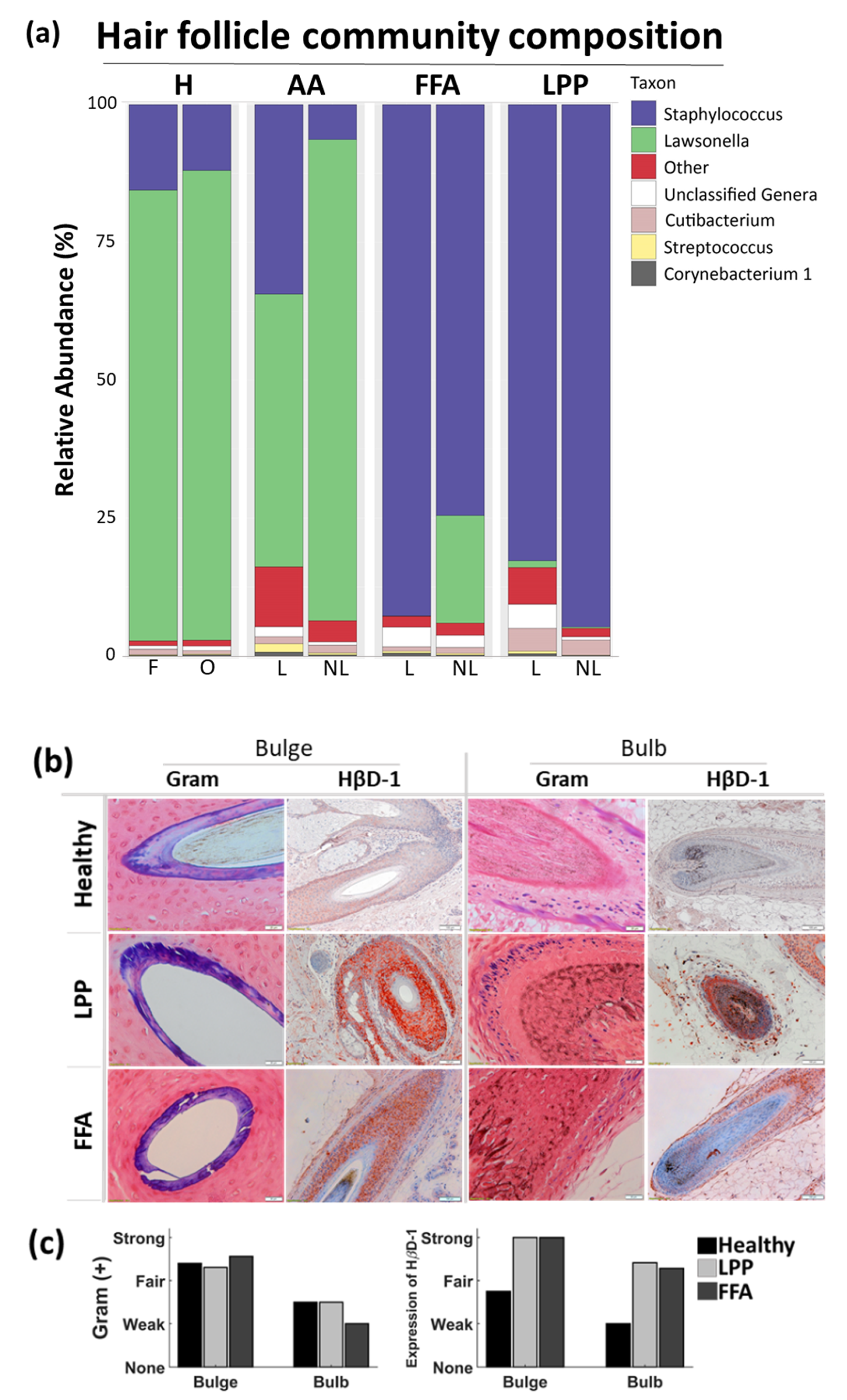
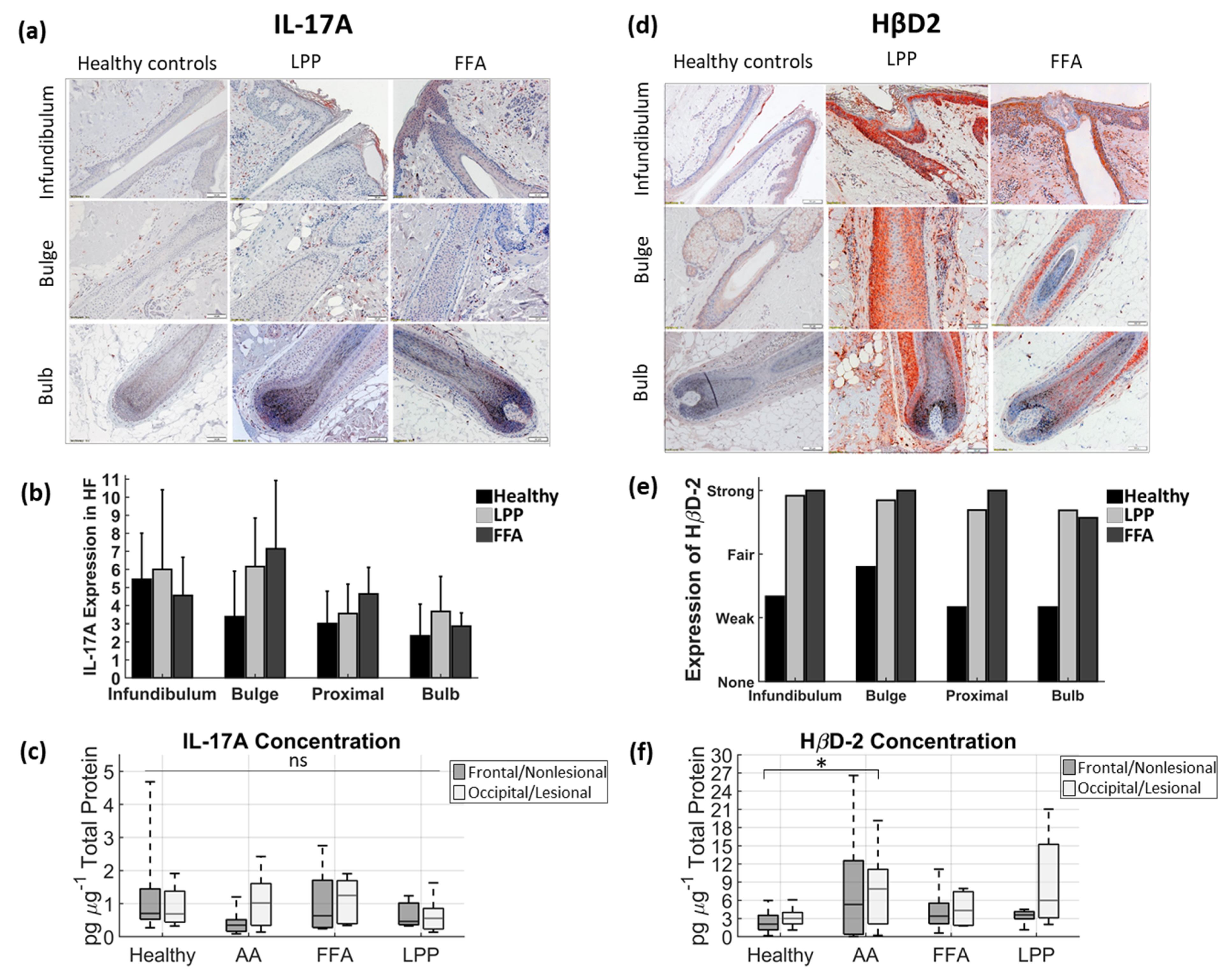
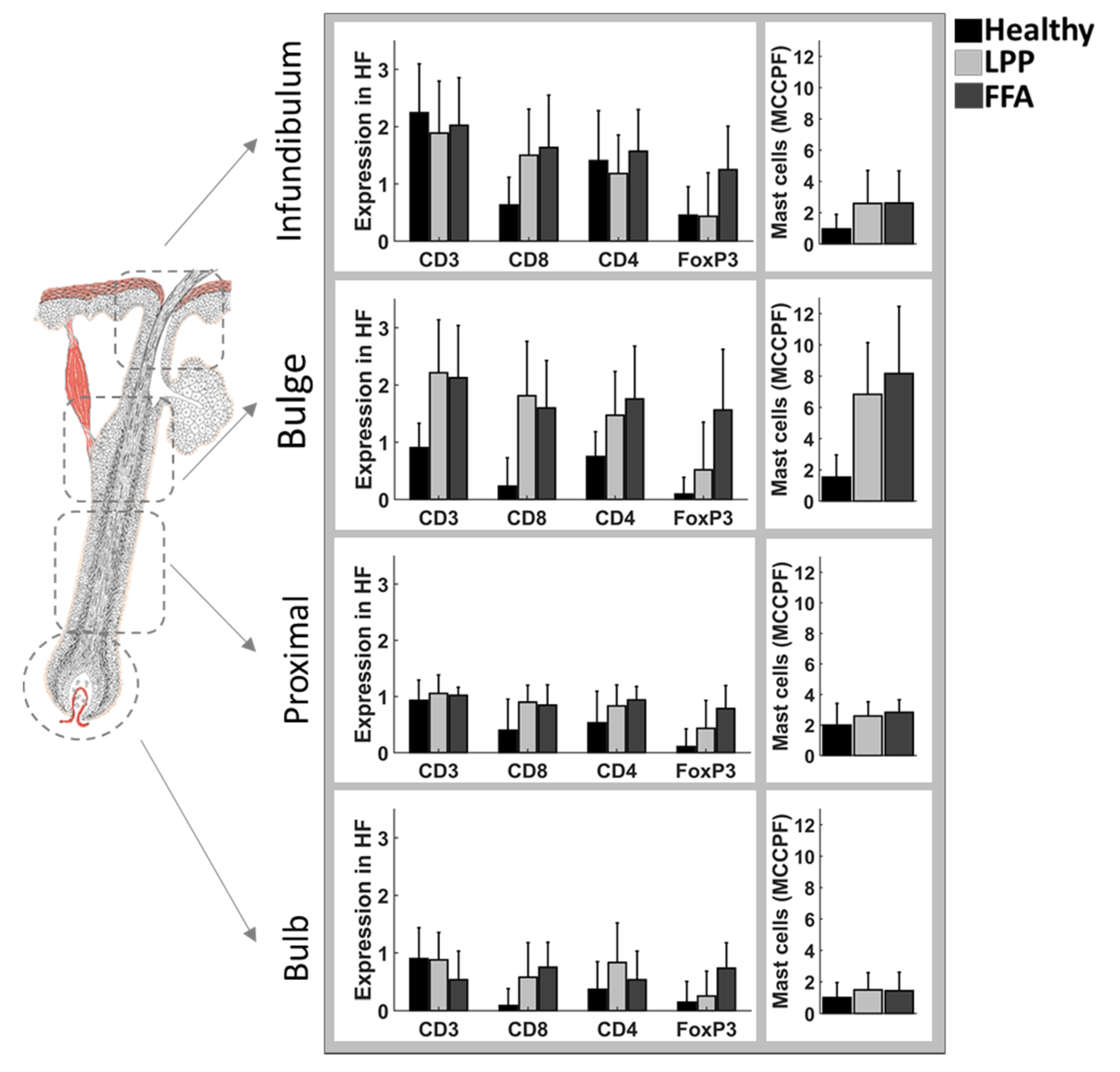
3.2. Below the Surface
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vano-Galvan, S.; Saceda-Corralo, D.; Blume-Peytavi, U.; Cucchia, J.; Dlova, N.C.; Gavazzoni Dias, M.F.R.; Grimalt, R.; Guzman-Sanchez, D.; Harries, M.; Ho, A.; et al. Frequency of the Types of Alopecia at Twenty-Two Specialist Hair Clinics: A Multicenter Study. Skin Appendage Disord 2019, 5, 309–315. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, A.; Clark, C.; Holmes, S. Frontal fibrosing alopecia: A review of 60 cases. J. Am. Acad. Dermatol. 2012, 67, 955–961. [Google Scholar] [CrossRef] [PubMed]
- Kanti, V.; Constantinou, A.; Reygagne, P.; Vogt, A.; Kottner, J.; Blume-Peytavi, U. Frontal fibrosing alopecia: Demographic and clinical characteristics of 490 cases. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 1976–1983. [Google Scholar] [CrossRef] [PubMed]
- Saceda-Corralo, D.; Pindado-Ortega, C.; Moreno-Arrones, O.M.; Ortega-Quijano, D.; Fernandez-Nieto, D.; Jimenez-Cauhe, J.; Vano-Galvan, S. Association of Inflammation With Progression of Hair Loss in Women With Frontal Fibrosing Alopecia. JAMA Dermatol. 2020, 156, 700. [Google Scholar] [CrossRef] [PubMed]
- Cerqueira, E.R.; Valente, N.; Sotto, M.N.; Romiti, R. Comparative Analysis of Immunopathological Features of Lichen Planopilaris and Female Patients with Frontal Fibrosing Alopecia. Int. J. Trichology 2016, 8, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Galvez-Canseco, A.; Sperling, L. Lichen planopilaris and frontal fibrosing alopecia cannot be differentiated by histopathology. J. Cutan. Pathol. 2018, 45, 313–317. [Google Scholar] [CrossRef]
- Chen, C.L.; Huang, W.Y.; Wang, E.H.C.; Tai, K.Y.; Lin, S.J. Functional complexity of hair follicle stem cell niche and therapeutic targeting of niche dysfunction for hair regeneration. J. Biomed. Sci. 2020, 27, 43. [Google Scholar] [CrossRef] [Green Version]
- Imanishi, H.; Ansell, D.M.; Cheret, J.; Harries, M.; Bertolini, M.; Sepp, N.; Biro, T.; Poblet, E.; Jimenez, F.; Hardman, J.; et al. Epithelial-to-Mesenchymal Stem Cell Transition in a Human Organ: Lessons from Lichen Planopilaris. J. Invest. Dermatol. 2018, 138, 511–519. [Google Scholar] [CrossRef] [Green Version]
- Harries, M.J.; Jimenez, F.; Izeta, A.; Hardman, J.; Panicker, S.P.; Poblet, E.; Paus, R. Lichen Planopilaris and Frontal Fibrosing Alopecia as Model Epithelial Stem Cell Diseases. Trends Mol. Med. 2018, 24, 435–448. [Google Scholar] [CrossRef]
- Aldoori, N.; Dobson, K.; Holden, C.R.; McDonagh, A.J.; Harries, M.; Messenger, A.G. Frontal fibrosing alopecia: Possible association with leave-on facial skin care products and sunscreens; a questionnaire study. Br. J. Dermatol. 2016, 175, 762–767. [Google Scholar] [CrossRef]
- Tavakolpour, S.; Mahmoudi, H.; Abedini, R.; Kamyab Hesari, K.; Kiani, A.; Daneshpazhooh, M. Frontal fibrosing alopecia: An update on the hypothesis of pathogenesis and treatment. Int. J. Womens Dermatol. 2019, 5, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Polak-Witka, K.; Rudnicka, L.; Blume-Peytavi, U.; Vogt, A. The role of the microbiome in scalp hair follicle biology and disease. Exp. Dermatol. 2019, 29, 286–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constantinou, A.; Kanti, V.; Polak-Witka, K.; Blume-Peytavi, U.; Spyrou, G.M.; Vogt, A. The Potential Relevance of the Microbiome to Hair Physiology and Regeneration: The Emerging Role of Metagenomics. Biomedicines 2021, 9, 236. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuji, T.; Chiang, H.I.; Jiang, S.B.; Nagarajan, H.; Zengler, K.; Gallo, R.L. The microbiome extends to subepidermal compartments of normal skin. Nat. Commun. 2013, 4, 1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polak-Witka, K.; Constantinou, A.; Schwarzer, R.; Helmuth, J.; Wiessner, A.; Hadam, S.; Kanti, V.; Rancan, F.; Andruck, A.; Richter, C.; et al. Identification of anti-microbial peptides and traces of microbial DNA in infrainfundibular compartments of human scalp terminal hair follicles. Eur. J. Dermatol. 2020. [Google Scholar] [CrossRef]
- Lademann, J.; Knorr, F.; Richter, H.; Blume-Peytavi, U.; Vogt, A.; Antoniou, C.; Sterry, W.; Patzelt, A. Hair follicles--an efficient storage and penetration pathway for topically applied substances. Summary of recent results obtained at the Center of Experimental and Applied Cutaneous Physiology, Charite -Universitatsmedizin Berlin, Germany. Skin Pharmacol. Physiol 2008, 21, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Blume-Peytavi, U.; Vogt, A. Human hair follicle: Reservoir function and selective targeting. Br. J. Dermatol. 2011, 165 (Suppl. S2), 13–17. [Google Scholar] [CrossRef]
- Chen, C.C.; Plikus, M.V.; Tang, P.C.; Widelitz, R.B.; Chuong, C.M. The Modulatable Stem Cell Niche: Tissue Interactions during Hair and Feather Follicle Regeneration. J. Mol. Biol. 2016, 428, 1423–1440. [Google Scholar] [CrossRef] [Green Version]
- Vogt, A.; Constantinou, A.; Rancan, F.; Ghoreschi, K.; Blume-Peytavi, U.; Combadiere, B. A niche in the spotlight: Could external factors critically disturb hair follicle homeostasis and contribute to inflammatory hair follicle diseases? Exp. Dermatol. 2020. [Google Scholar] [CrossRef]
- Otberg, N.; Kang, H.; Alzolibani, A.A.; Shapiro, J. Folliculitis decalvans. Dermatol. Ther. 2008, 21, 238–244. [Google Scholar] [CrossRef]
- Chiarini, C.; Torchia, D.; Bianchi, B.; Volpi, W.; Caproni, M.; Fabbri, P. Immunopathogenesis of folliculitis decalvans: Clues in early lesions. Am. J. Clin. Pathol. 2008, 130, 526–534. [Google Scholar] [CrossRef] [Green Version]
- Campbell, D.J.; Koch, M.A. Living in Peace: Host-Microbiota Mutualism in the Skin. Cell Host Microbe 2017, 21, 419–420. [Google Scholar] [CrossRef] [Green Version]
- Scharschmidt, T.C.; Vasquez, K.S.; Pauli, M.L.; Leitner, E.G.; Chu, K.; Truong, H.A.; Lowe, M.M.; Sanchez Rodriguez, R.; Ali, N.; Laszik, Z.G.; et al. Commensal Microbes and Hair Follicle Morphogenesis Coordinately Drive Treg Migration into Neonatal Skin. Cell Host Microbe 2017, 21, 467–477.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillmann, K.; Blume-Peytavi, U. Diagnosis of hair disorders. Semin. Cutan. Med. Surg 2009, 28, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Finlay, A.Y.; Khan, G.K. Dermatology Life Quality Index (DLQI)—A simple practical measure for routine clinical use. Clin. Exp. Dermatol. 1994, 19, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Vano-Galvan, S.; Molina-Ruiz, A.M.; Fernandez-Crehuet, P.; Rodrigues-Barata, A.R.; Arias-Santiago, S.; Serrano-Falcon, C.; Martorell-Calatayud, A.; Barco, D.; Perez, B.; Serrano, S.; et al. Folliculitis decalvans: A multicentre review of 82 patients. J. Eur. Acad. Dermatol. Venereol. 2015, 29, 1750–1757. [Google Scholar] [CrossRef]
- Olsen, E.A.; Hordinsky, M.K.; Price, V.H.; Roberts, J.L.; Shapiro, J.; Canfield, D.; Duvic, M.; King, L.E., Jr.; McMichael, A.J.; Randall, V.A.; et al. Alopecia areata investigational assessment guidelines--Part II. National Alopecia Areata Foundation. J. Am. Acad. Dermatol. 2004, 51, 440–447. [Google Scholar] [CrossRef]
- Chiang, C.; Sah, D.; Cho, B.K.; Ochoa, B.E.; Price, V.H. Hydroxychloroquine and lichen planopilaris: Efficacy and introduction of Lichen Planopilaris Activity Index scoring system. J. Am. Acad. Dermatol. 2010, 62, 387–392. [Google Scholar] [CrossRef]
- Holmes, S.; Ryan, T.; Young, D.; Harries, M.; British, H.; Nail, S. Frontal Fibrosing Alopecia Severity Index (FFASI): A validated scoring system for assessing frontal fibrosing alopecia. Br. J. Dermatol. 2016, 175, 203–207. [Google Scholar] [CrossRef] [Green Version]
- Graziano, M.U.; Graziano, K.U.; Pinto, F.M.; Bruna, C.Q.; de Souza, R.Q.; Lascala, C.A. Effectiveness of disinfection with alcohol 70% (w/v) of contaminated surfaces not previously cleaned. Rev. Lat Am. Enferm. 2013, 21, 618–623. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Team, R.C. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online: https://www.R-project.org (accessed on 2 March 2021).
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leo, L.; Shetty, S.S. Tools for Microbiome Analysis in R. Microbiome Package Version. Bioconductor 2017–2019. Available online: http://microbiome.github.io/microbiome (accessed on 5 March 2021).
- Ssekagiri, A.; Sloan, W.T.; Ijaz, U.Z. microbiomeSeq: An R package for analysis of microbial communities in an environmental context. In Proceedings of the ISCB Africa ASBCB Conference, Kumasi, Ghana, 9 August 2017. [Google Scholar] [CrossRef]
- Ocejo, M.; Oporto, B.; Hurtado, A. 16S rRNA amplicon sequencing characterization of caecal microbiome composition of broilers and free-range slow-growing chickens throughout their productive lifespan. Sci. Rep. 2019, 9, 2506. [Google Scholar] [CrossRef] [Green Version]
- Grice, E.A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Davis, J.; Young, A.C.; Program, N.C.S.; Bouffard, G.G.; Blakesley, R.W.; Murray, P.R.; et al. Topographical and temporal diversity of the human skin microbiome. Science 2009, 324, 1190–1192. [Google Scholar] [CrossRef] [Green Version]
- Fahlen, A.; Engstrand, L.; Baker, B.S.; Powles, A.; Fry, L. Comparison of bacterial microbiota in skin biopsies from normal and psoriatic skin. Arch. Dermatol. Res. 2012, 304, 15–22. [Google Scholar] [CrossRef]
- Haskin, A.; Fischer, A.H.; Okoye, G.A. Prevalence of Firmicutes in Lesions of Hidradenitis Suppurativa in Obese Patients. JAMA Dermatol. 2016, 152, 1276–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ring, H.C.; Riis Mikkelsen, P.; Miller, I.M.; Jenssen, H.; Fuursted, K.; Saunte, D.M.; Jemec, G.B. The bacteriology of hidradenitis suppurativa: A systematic review. Exp. Dermatol. 2015, 24, 727–731. [Google Scholar] [CrossRef]
- Sillani, C.; Bin, Z.; Ying, Z.; Zeming, C.; Jian, Y.; Xingqi, Z. Effective treatment of folliculitis decalvans using selected antimicrobial agents. Int. J. Trichology 2010, 2, 20–23. [Google Scholar] [CrossRef] [Green Version]
- Annessi, G. Tufted folliculitis of the scalp: A distinctive clinicohistological variant of folliculitis decalvans. Br. J. Dermatol. 1998, 138, 799–805. [Google Scholar] [CrossRef]
- Leyden, J.J.; Marples, R.R.; Kligman, A.M. Staphylococcus aureus in the lesions of atopic dermatitis. Br. J. Dermatol. 1974, 90, 525–530. [Google Scholar] [CrossRef]
- Pinto, D.; Sorbellini, E.; Marzani, B.; Rucco, M.; Giuliani, G.; Rinaldi, F. Scalp bacterial shift in Alopecia areata. PLoS ONE 2019, 14, e0215206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, K.; Gallo, R.L. Antimicrobial peptides in human skin disease. Eur. J. Dermatol. 2008, 18, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Ali, R.S.; Falconer, A.; Ikram, M.; Bissett, C.E.; Cerio, R.; Quinn, A.G. Expression of the peptide antibiotics human beta defensin-1 and human beta defensin-2 in normal human skin. J. Invest. Dermatol. 2001, 117, 106–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niyonsaba, F.; Ogawa, H.; Nagaoka, I. Human beta-defensin-2 functions as a chemotactic agent for tumour necrosis factor-alpha-treated human neutrophils. Immunology 2004, 111, 273–281. [Google Scholar] [CrossRef]
- Abiko, Y.; Jinbu, Y.; Noguchi, T.; Nishimura, M.; Kusano, K.; Amaratunga, P.; Shibata, T.; Kaku, T. Upregulation of human beta-defensin 2 peptide expression in oral lichen planus, leukoplakia and candidiasis. An immunohistochemical study. Pathol. Res. Pract. 2002, 198, 537–542. [Google Scholar] [CrossRef]
- Nishimura, M.; Abiko, Y.; Kusano, K.; Yamazaki, M.; Saitoh, M.; Mizoguchi, I.; Jinbu, Y.; Noguchi, T.; Kaku, T. Localization of human beta-defensin 3 mRNA in normal oral epithelium, leukoplakia, and lichen planus: An in situ hybridization study. Med. Electron Microsc. 2003, 36, 94–97. [Google Scholar] [CrossRef]
- Chung, W.O.; Dommisch, H.; Yin, L.; Dale, B.A. Expression of defensins in gingiva and their role in periodontal health and disease. Curr. Pharm. Des. 2007, 13, 3073–3083. [Google Scholar] [CrossRef]
- Gursoy, U.K.; Kononen, E. Understanding the roles of gingival beta-defensins. J. Oral Microbiol. 2012, 4. [Google Scholar] [CrossRef]
- Shibagaki, N.; Suda, W.; Clavaud, C.; Bastien, P.; Takayasu, L.; Iioka, E.; Kurokawa, R.; Yamashita, N.; Hattori, Y.; Shindo, C.; et al. Aging-related changes in the diversity of women’s skin microbiomes associated with oral bacteria. Sci. Rep. 2017, 7, 10567. [Google Scholar] [CrossRef]
- Juge, R.; Rouaud-Tinguely, P.; Breugnot, J.; Servaes, K.; Grimaldi, C.; Roth, M.P.; Coppin, H.; Closs, B. Shift in skin microbiota of Western European women across aging. J. Appl. Microbiol. 2018, 125, 907–916. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, J.J.; Myeong, N.R.; Kim, T.; Kim, D.; An, S.; Kim, H.; Park, T.; Jang, S.I.; Yeon, J.H.; et al. Segregation of age-related skin microbiome characteristics by functionality. Sci. Rep. 2019, 9, 16748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Park, T.; Yun, J.I.; Lim, H.W.; Han, N.R.; Lee, S.T. Investigation of Age-Related Changes in the Skin Microbiota of Korean Women. Microorganisms 2020, 8, 1581. [Google Scholar] [CrossRef] [PubMed]
- Pouralibaba, F.; Babaloo, Z.; Pakdel, F.; Aghazadeh, M. Serum Level of Interleukin 17 in Patients with Erosive and Non erosive Oral Lichen Planus. J. Dent. Res. Dent. Clin. Dent. Prospect. 2013, 7, 91–94. [Google Scholar] [CrossRef]
- Solimani, F.; Pollmann, R.; Schmidt, T.; Schmidt, A.; Zheng, X.; Savai, R.; Muhlenbein, S.; Pickert, J.; Eubel, V.; Mobs, C.; et al. Therapeutic Targeting of Th17/Tc17 Cells Leads to Clinical Improvement of Lichen Planus. Front. Immunol 2019, 10, 1808. [Google Scholar] [CrossRef] [Green Version]
- Bain, K.A.; McDonald, E.; Moffat, F.; Tutino, M.; Castelino, M.; Barton, A.; Cavanagh, J.; Ijaz, U.Z.; Siebert, S.; McInnes, I.B.; et al. Alopecia areata is characterized by dysregulation in systemic type 17 and type 2 cytokines, which may contribute to disease-associated psychological morbidity. Br. J. Dermatol. 2020, 182, 130–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanda, N.; Kamata, M.; Tada, Y.; Ishikawa, T.; Sato, S.; Watanabe, S. Human beta-defensin-2 enhances IFN-gamma and IL-10 production and suppresses IL-17 production in T cells. J. Leukoc. Biol. 2011, 89, 935–944. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Subjects | Healthy Controls | AAc | LPP | FFA | ||||
|---|---|---|---|---|---|---|---|---|
| n = 12 | % | n = 7 | % | n = 6 | % | n = 6 | % | |
| Male | 6 | 50 | 0 | 0 | 0 | 0 | 0 | 0 |
| Female | 6 | 50 | 7 | 100 | 6 | 100 | 6 | 100 |
| Mean age, years (SD) | 28.7 (7.2) | 41.3 (15.9) | 54.8 (18.1) | 56.8 (7.4) | ||||
| Mean Weight, kg (SD) | 67.1 (10.8) | 70 (6.5) | 62.7 (17.8) | 71.8 (7.7) | ||||
| Mean Height, cm (SD) | 170.8 (10.2) | 171.4 (4.2) | 166 (6.4) | 167.4 (7.1) | ||||
| Atopic Dermatitis | 0 | 0 | 0 | 0 | ||||
| Psoriasis | 0 | 0 | 0 | 0 | ||||
| Fitzpatrick Phototype | ||||||||
| Type II | 5 | 42% | 2 | 29% | 1 | 17% | 1 | 17 |
| Type II/III | 0 | 0% | 0 | 0% | 1 | 17% | 1 | 17 |
| Type III | 3 | 25% | 3 | 43% | 4 | 67% | 1 | 33 |
| Type III/IV | 0 | 0% | 2 | 29% | 0 | 0% | 0 | 0 |
| Type IV | 3 | 25% | 0 | 0% | 0 | 0% | 2 | 33 |
| Type V | 1 | 8% | 0 | 0% | 0 | 0% | 0 | 0 |
| Scalp pH (SD) | ||||||||
| Frontal | 6 (1.5) | n/a | n/a | n/a | ||||
| Occipital | 5.7 (1.2) | n/a | n/a | n/a | ||||
| Affected | n/a | 6.9 (1.1) | 6.8 (0.8) | 6.5 (0.8) | ||||
| Not Affected | n/a | 6.9 (2) | 6.9 (1.3) | 6.3 (0.6) | ||||
| Scalp Sebum (SD) | ||||||||
| Frontal | 107.4 (96.5) | n/a | n/a | n/a | ||||
| Occipital | 78.8 (73.6) | n/a | n/a | n/a | ||||
| Affected | n/a | 44.3 (49.5) | 71.3 (65.9) | 29 (39.1) | ||||
| Not Affected | n/a | 16.1 (14.5) | 60.2 (20.1) | 41 (37.9) | ||||
| DLQI (mean, SD) | n/a | 5 (3.8) | 5.8 (2.1) | 4.2 (3.7) | ||||
| Severity Score1 (mean, SD) | n/a | 20.3 (15.2) | 6.6 (2.4) | 33.9 (6.2) | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Constantinou, A.; Polak-Witka, K.; Tomazou, M.; Oulas, A.; Kanti, V.; Schwarzer, R.; Helmuth, J.; Edelmann, A.; Blume-Peytavi, U.; Spyrou, G.M.; et al. Dysbiosis and Enhanced Beta-Defensin Production in Hair Follicles of Patients with Lichen Planopilaris and Frontal Fibrosing Alopecia. Biomedicines 2021, 9, 266. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9030266
Constantinou A, Polak-Witka K, Tomazou M, Oulas A, Kanti V, Schwarzer R, Helmuth J, Edelmann A, Blume-Peytavi U, Spyrou GM, et al. Dysbiosis and Enhanced Beta-Defensin Production in Hair Follicles of Patients with Lichen Planopilaris and Frontal Fibrosing Alopecia. Biomedicines. 2021; 9(3):266. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9030266
Chicago/Turabian StyleConstantinou, Andria, Katarzyna Polak-Witka, Marios Tomazou, Anastasis Oulas, Varvara Kanti, Rolf Schwarzer, Johannes Helmuth, Anke Edelmann, Ulrike Blume-Peytavi, George M. Spyrou, and et al. 2021. "Dysbiosis and Enhanced Beta-Defensin Production in Hair Follicles of Patients with Lichen Planopilaris and Frontal Fibrosing Alopecia" Biomedicines 9, no. 3: 266. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9030266