
MiR-7 in Cancer Development


1
Department of Biology, Division of Molecular Biology, Faculty of Science, University of Zagreb, Horvatovac 102, 10000 Zagreb, Croatia
2
Division of Molecular Biology, Rudjer Bosković Institute, Bijenička 54, 10000 Zagreb, Croatia
*
Author to whom correspondence should be addressed.
Biomedicines 2021, 9(3), 325; https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9030325
Submission received: 26 February 2021
/
Revised: 20 March 2021
/
Accepted: 22 March 2021
/
Published: 23 March 2021
(This article belongs to the Special Issue MicroRNA in Solid Tumor and Hematological Diseases)
Abstract
:MicroRNAs (miRNAs) are short non-coding RNA involved in the regulation of specific mRNA translation. They participate in cellular signaling circuits and can act as oncogenes in tumor development, so-called oncomirs, as well as tumor suppressors. miR-7 is an ancient miRNA involved in the fine-tuning of several signaling pathways, acting mainly as tumor suppressor. Through downregulation of PI3K and MAPK pathways, its dominant role is the suppression of proliferation and survival, stimulation of apoptosis and inhibition of migration. Besides these functions, it has numerous additional roles in the differentiation process of different cell types, protection from stress and chromatin remodulation. One of the most investigated tissues is the brain, where its downregulation is linked with glioblastoma cell proliferation. Its deregulation is found also in other tumor types, such as in liver, lung and pancreas. In some types of lung and oral carcinoma, it can act as oncomir. miR-7 roles in cell fate determination and maintenance of cell homeostasis are still to be discovered, as well as the possibilities of its use as a specific biotherapeutic.
1. Introduction
MicroRNAs are short non-coding RNAs involved in the regulation of specific mRNA translation. Through this process, they regulate numerous cellular functions, participate in signaling circuits and fine-tune cellular differentiation.
miRNAs (miRs) have a complex pathway of biogenesis and regulation of their function. While final miRNAs are short single-stranded noncoding RNAs of 20–23 nt, they start as pri-miRNAs, several hundred base pairs long with a complex formation pathway. These primary miRNAs are first processed by a microprocessor containing Drosha, an enzyme that cleaves the stem of a hairpin structure formed by future miR sequence and producing pre-miRNA. After nuclear export, further processing is done by Dicer in the cytoplasm, which removes the loop region and produces miRNA duplex. Only one strand of the duplex is chosen to become the mature miRNA, loaded on an RNA-induced silencing complex (RISC) containing the Argonaute protein. RISC complex with specific miR targets complementary mRNAs and fully complementary mRNA are degraded. Since mature miRNAs in higher eukaryotic cells most often are not fully complementary to their target mRNA, they can lead to translation inhibition [1].
Usually, one mRNA can be targeted by several miRNAs on its 3’UTR. It is supposed that the target site spacing can influence cooperative repression. Although a great number of genes can be influenced by a single miR, in general, miRs act according to the cellular program in a specific cell type and target only a subset of transcripts [2]. However, the regulation of these processes is still poorly understood.
One of the first known, and also most investigated miRNAs is miR-7. The seed sequence GGAAGA is evolutionarily conserved and is found in Nematodes, Insects and Vertebrates [3]. In Mammals miR-7 dominantly acts as a tumor suppressor and regulates several basic cellular processes, which include proliferation, differentiation, apoptosis, migration and expression of stem cell features. It was also one of the miRs used for the classification of the regulatory clusters. Most discoveries are in regard to its role in the brain and sensory cell differentiation in man and Drosophila, respectively. Li X, and his collaborators (2009) suggest that miRs, in general, may stabilize different regulatory networks depending on the conditions of environmental fluctuation during development [4]. This hypothesis was developed on an example of miR-7, participating in Notch and Epidermal growth factor receptor (EGFR) coherent and incoherent feedforward loops during photoreceptor determination in Drosophila. On the other hand, miR-7 downregulation is linked to cell proliferation in many tumors, and its regulation is tightly connected with differentiation processes in the pancreas, brain and other organs [5,6].
miR-7 is encoded in three different sites in the human genome. MIR7-1 sequence is present inside the last intron of the heterogeneous nuclear ribonucleoprotein K (hnRNPK) gene, on chromosome 9 (9q21.32) and is considered to be the dominant gene responsible for miR-7 expression. MIR7-2 sequence is present in the intergenic region on chromosome 15, and MIR7-3 in the intron of pituitary gland specific factor 1 gene (PGSF1) or MIR-7 host gene on chromosome 19 [7].
2. Regulation of MiR-7 Expression
miRNA genes, like the protein-coding genes, have a regulated promoter and their products are members of signaling circuits of different cellular processes. miRs are also regulated at several steps during processing into their active form by means of binding to different proteins [8]. miRs can bind different long non-coding RNAs and circular RNAs either to be degraded or to be “preserved” for later function. Different RNA classes can function as miR “sponges” and bind miRs to keep them out of function: 3’UTR mRNA [9], long non-coding RNAs (lncRNAs) and circular RNAs. Different proteins can also regulate pri-miR degradation [1].
miR-7 is considered to be a network stabilizer, connecting different signaling pathways through feedback and feedforward loops [4]. Its function in buffering gene expression and providing robustness in cell response was demonstrated. Caygill and Brand (2017) showed on the Drosophila model where miR-7 targeted the Notch pathway, that miR-7 buffers the differentiation of the neuroepithelial cells into neuroblasts. Its role was to enable precision in the process despite conditions of environmental stress [10].
As a tumor suppressor, miR-7 expression is often downregulated in different cancer cells (i.e., in brain, lung and colon cancer cells [11,12,13]). Interestingly, it is also involved in signaling circuits directing differentiation in different tissues and it is regulated by specific transcription factors [6,14,15,16]. miR-7 promoter was found to be silenced by DNA methylation in cancer stem cells [17]. In breast carcinoma, its expression is estrogen-dependent [18]. Duex et al. found miR-7 to be in a signaling loop with EGFR through Usp18 (Ubp43), a ubiquitin-specific peptidase, whose downregulation elevates miR-7 levels [19]. It was also found that Hepatitis B virus (HBV) protein HBx can upregulate miR-7 expression through EGFR [20] and in breast cells by hepatocyte growth factor (HGF) [21]. However, miR-7 inhibition promotes breast cancer metastasis [22].
miR-7 was found to belong to a p53-dependent non-coding RNA network [23,24], as well as the Myc signaling circuit [25]. Hansen et al. described the existence of circular RNAs, which can pair with complementary miRNAs [26]. Circular RNAs have a structure of covalently closed single-stranded RNA molecules, produced by a specific type of splicing. These molecules are more stable than linear. Some of them can act as miR sponges: RNA molecules, which contain multiple target sites complementary to a specific miR and influence its activities by binding to it. The first such molecule was detected in neurons and it was Cdr1as (ciRS-7) regulating miR-7. It contained miR-7 sequences transcribed in the antisense orientation from the CDR1 gene, forming circular RNA (circRNA) Cdr1as with more than 70 binding sites for miR-7 and one perfectly complementary site for miR-671 [26,27,28]. It seems that Cdr1as binds miR-7s and serves as their reservoir, and their release is regulated with miR-671, which causes cleavage of Cdr1as and liberation of miR-7s to exert their activities. Furthermore, Kleaveland et al. ( found miR-7 to be a member of a regulatory network consisting of four ncRNAs: one long ncRNA, one circular and two microRNAs, in the mouse cerebellum [29]. Cyrano is a long ncRNA, which pairs to miR-7 and triggers its destruction. At the same time, this long ncRNA enables upregulation of circular Cdr1as, otherwise downregulated by miR-7. miR-671 was found to be involved in Cdr1as destruction.
Numerous long noncoding RNAs were found to bind to miR-7 and downregulate its activities: LINC00115 and XIST in breast cancer [22,30], LINC00240 in lung cancer [31], RSC1-AS1 in hepatocellular carcinoma, TINCR and Zing Finger Antisense 1 (ZFAS1) in breast and colorectal cancer [32,33,34], LPP-AS2 in glioma cells [35], etc. LncRNA SOX21-AS1 influenced cervical cancer progression by inhibiting miR-7/VDAC1 (voltage-dependent anion channel 1) [36]. lncRNA KCNQ1OT1 modulated cell resistance to chemotherapy [37], and lncRNA FOXD2-AS1 was found to bind miR-7 in thyroid cancer, upregulating the expression of hTERT [38]. lncRNA UCA1 downregulated miR-7, influencing the EGFR axis in gastric cancer cells resistant to hypoxia [39]. Upregulation of long noncoding RNA ANRIL caused by hypoxia modulated miR-7/SIRT1 axis and protected cells from cell death [40]. lncRNA CASC21 influenced miR-7/YAP1 signaling in colorectal cancer [41], and lncRNA Oip5-as1 in stem cells was found to modulate NANOG expression [42].
Several circular RNAs, besides ciRS, also regulate miR-7 and its downstream targets: circHIPK3 in colorectal cancer, circ-0015756 in hepatocellular carcinoma influencing downstream FAK [43,44], hsa_circRNA_0006528 in breast cancer influencing proliferation through MAPK/ERK pathway [45], circ-U2AF1 in gliomas influencing the expression of NOVA 2 [46], circ-TFCP2L1 decreasing mir-7-PAK1 signaling [47], circAkap17b regulating FSH secretion in pituitary gland [48]. circSNCA, SNCA and miR-7 were found to be regulated by endogenous competition and could influence the progression of Parkinson’s disease [49].
Other types of RNA can also modulate miR-7 activity: 3′UTR Ube3a-1 mRNA [9] and Small Nucleolar RNA Host Gene 15 (SNHG15) regulating Klf4 through miR-7 [50].
There are also several proteins, which influence miR-7 maturation. Wang et al. described miR-7 regulation by protein quaking isoforms (QKI) [51]. The QKI proteins have heteronuclear ribonucleoprotein particle K (hnRNPK) homology KH and belong to RNA binding proteins. These proteins interact with a QKI response element sequence in introns and mature mRNAs, and it was shown that nuclear isoforms QKI-5 and QKI-6 associated with pri-miR-7-1 to prevent its processing. They were also found to interact with Ago2, during stress conditions. Similarly, it was shown that miR-7 can be downregulated by NF90-NF45 complex, through the binding of this double-stranded RNA-binding protein complex to primary miR-7 [52]. miR-7, in turn, targeted the coding region of NF90 mRNA. Nerve cells have a posttranscriptional regulation of miR-7 through the expression of Musashi homolog 2 (MSI2) and Hu antigen R (HuR), miR processing inhibitors and tissue-specific factors, regulating miR-7 expression and activity during neural differentiation [8]. A similar regulation was found in human lung cancer cells as a response to TLR9 signaling [53]. In addition, mechanisms of miR-7 targeted degradation linked with its 3′ end modifications were recently discovered [54].
On the other side, SF2/ASF increases Drosha cleavage of primary miR-7 transcript and promotes miR-7 maturation, and miR-7 in the feedback loop can decrease SF2/ASF expression. This molecule does not only affect miR-7, but also other miRs, coordinating their splicing regulation and gene repression [55] (Figure 1).
3. MiR-7 and Chromatin Regulation
miR-7 was found to regulate a number of genes involved in chromatin modulation. It can downregulate histone methyl-transferase gene, SETDB1 in different types of cancer cells [56,57], as well as TET2 and SMARCD1 [58,59]. It can also influence global cellular expression through the regulation of master transcription factors, such as KLF4, and thus impact the fate of cancer stem cells and human embryonic stem cells [60]. miR-7 is also found in extracellular vesicles and besides the possibility to influence the fate of the cell where it is expressed, it could also interfere with the biology of the cells to which it is delivered [61].
4. MiR-7 in Nerve Cells and Glioblastoma
In brain development, a fine regulation of cell proliferation, cell differentiation and regulation of symmetric and asymmetric division, as well as cell migration is necessary. It seems that miR-7 has a role in fine-tuning of these processes, in general as a suppressor of proliferation (Table 1, Table 2, Table 3 and Table 4, Figure 2).
It has spatiotemporal-dependent expression and regulation [28], and it is found in discrete brain regions [3]. It can also have specific subcellular localization, different in the cell body and neurites. One example is miR-7 role in dopaminergic neuron differentiation by fine-tuning Pax6 expression [5]. miR-7 also regulates other neural fate markers, elements of the Wnt pathway, interferes with Hedgehog and Notch signaling and takes part in the differentiation process [134]. miR-7 regulates both, specific nerve functions (such as synaptic [144]) and master regulators (such as HoxD family members). It is detected as one of the miRNAs forming “miR signature” in neural stem and neural cancer stem cells [154], which is in accordance with its role in differentiation and proliferation. Interestingly, miR-7 is 40 times more abundant in neurons than in astrocytes (Table 1).
Besides influencing cell differentiation, and thus indirectly interfering with it, miR-7 can also directly inhibit cell proliferation (Figure 2). In glioblastoma and neuroblastoma miR-7 was found to be downregulated compared to normal tissue, indicating its role as a tumor suppressor [11,62]. The functions of miR-7 in glioblastomas are mainly linked to its influence on cell proliferation, differentiation, apoptosis and migration. Although some glioblastoma cells can be refractory to miR-7 expression, its downregulation is often found in nerve cell tumors. Saydam et al. found its downregulation to be the typical miR schwannoma characteristic signature [11].
One of the first detected and most investigated targets of miR-7 is Epidermal growth factor receptor, EGFR, whose protein expression is decreased by miR activity. EGFR is linked to several important proliferation-inducing pathways, such as PI3K/Akt and MAPK and their downregulation leads to decreased activation of the Akt and ERK1/2. Kefas found that miR-7 directly regulates EGFR expression [62].
miR-7 targets are also several other proteins involved in downstream signaling. In the PI3K/Akt pathway, these are Akt pathway regulators IRS-1 and IRS-2, PI3K subunits (PIK3R3 and PIK3CD), mTOR [66], and PAK1 (p21/Cdc42/Rac1-activated kinase) [67,155]. The latter is potentially involved not only in oncogenic signaling through EGFR/Akt, but also in motility, regulation of cytoskeleton and apoptosis [63]. On the MAPK pathway, miR-7 influences Raf1 and ARF4 (ADP-ribosylation factor 4) expression, which modulates activation of phospholipase D2 (PLD2) and downstream activation of AP-1 [67,155]. Webster et al. found its influence on JNK and CAMK pathways [63]. In addition, Duex et al. found miR-7 to be involved in the signaling loop with EGFR through Usp18 (Ubp43), a ubiquitin-specific peptidase, whose downregulation elevates miR-7 levels [19].
miR-7 is also involved in the regulation of cell survival [67,84] as it downregulates pro-survival proteins IRS-1, IGF-1R, PAK1, and Raf-1 and leads to the reduction in cell viability. Zhang X et al. found that the expression pattern of miR-7 correlates with the glioblastoma cells’ sensitivity to apoptosis induced by TRAIL, a TNF family member [110]. XIAP, an apoptosis inhibitor, was detected as a direct miR-7 target (Table 1, Table 2 and Table 3).
In another experimental setting, Kabaria et al. found that miR-7 targeted 3’UTR of Keap1 in human neuroblastoma cells [119]. Keap1 takes part in the regulation of Nrf2, a transcription factor involved in the expression of many antioxidant and detoxifying genes in reactive oxygen species (ROS) defense. miR-7, therefore, participated in cellular protection from oxidative stress. In neuroblastoma cells, Sirtuin (Sirt 1) was found to be a direct target of miR-7, and a link to the regulation of oxygen-glucose deprivation and cerebral injury [116]. It was found that miR-7 can target VDAC1, voltage-dependent anion channel, a part of the mitochondrial permeability transition pore, leading to the decrease in the intracellular ROS and protection against mitochondrial dysfunction and cytotoxicity [117]. Jia et al. compared RNA expression in glioblastoma cell lines differently sensitive on alkylation DNA damage and found miR-7 to be downregulated in the resistant cells [115]. They showed that miR-7 upregulation increased the cell sensitivity to alkylation. As a direct target, transcription factor YY1 was identified. However, it is also possible that in glioblastoma cell lines cell-specific regulation exists and that not all cell lines are responsive to miR-7 expression [8,156].
miR-7 also targets the expression of proteins involved in migration and metastasis [99]. Increased expression of miR-7 inhibited migration and invasion through downregulation of MMP-2, MMP-9 and FAK, a kinase involved in motility. Different targets were found to link miR-7 to actin cytoskeleton: Rho GTPases, Ack1 and PAK. In addition, in glioblastoma its target was a special AT-rich sequence binding protein 1 (SATB1), a protein able to promote migration and invasion [105].
Pan CM et al. found miR-7 to target TBX2 mRNA, and due to miR-7 downregulation in glioblastomas, TBX2 is increased [96]. Its high expression correlated with poor prognosis and higher invasivity of glioblastoma cells, EMT features and pulmonary metastasis. TBX2 is involved in the developmental processes and morphogenesis of different organs. It represses E-cadherin and increases the invasiveness of breast cancer cells. miR-7 also influenced TFF3, a signaling molecule downstream of PI3K/Akt pathway. Its downregulation decreased migration and invasion. This process can be reversed by a glioblastoma cell treatment with a glycolytic inhibitor which reduces the expression of miR-7 [97].
5. MiR-7 Role in Gastrointestinal Tumours
In gastric cancer (GC) patients, miR-7 deregulation consequently leads to increased cell proliferation, tumorigenesis and poor survival. In gastric cancer cells, besides targeting the EGFR pathway, miR-7 targets the IGF1R and downstream RELA and FOS [77]. miR-7 indirectly influences RELA activation, through targeting IKKeta. Through the feedback circuit, the NF-kappaB pathway regulates the miR-7 expression. In addition, miR-7 can downregulate the IGF1R-Snail pathway, which is involved in epithelial-mesenchymal transition [74,77]. Similar pathways were influenced in tongue squamous cell carcinoma [71]. Recently it was found that miR-7 could target lactate dehydrogenase A (LDH-A) in gastric cancer cells, so its downregulation can influence glycolysis, cell proliferation and sensitivity to chemotherapy [150].
In oral squamous cell carcinoma cells miR-7 regulated the expression of RECK, which acts as a metalloproteinase inhibitor and can suppress cell proliferation and migration. Therefore, miR-7 acted as an oncogene, and RECK inhibition was associated with poor prognosis and aggressiveness of tumors [87] (Figure 2).
miR-7 has also been reported to target a specific set of genes in the liver. Some of them code for proteins involved in cell cycle and apoptosis regulation, such as CCNE1 [83], Bcl-2 and XIAP. In hepatocellular carcinoma cells miR-7 directly regulates CUL5, influencing cell proliferation and inducing cell cycle arrest [92]. As miR-7 targets Notch3, its downregulation leads to Notch signaling activation in the same type of cancer cells [157]. Besides Notch3, Notch4 and VE cadherin were also found to be miR-7 targets [107]. miR-7 also downregulates VDAC1 in hepatocellular carcinoma and influences proliferation and migration [118], as well as the fibroblast growth factor receptor FGFR4, a key molecule for liver protection from chronic injury. In the conditions of increased fibrosis miR-7 was found to be upregulated and promoted HSC proliferation and activation as a consequence of FGFR4 downregulation [132].
miR-7 is involved in the differentiation of pancreatic endocrine cells [6]. In pancreatic carcinoma, miR-7 can suppress NFAT. This transcription factor can regulate epithelial-mesenchymal transition and act as an oncogene in pancreatic carcinoma cells [106]. Downing et al. found miR-7 to directly target REG1, a protein that increases proliferation and influences apoptosis and differentiation of pancreatic cells [124]. miR-7 was found to suppress SOX18 and to influence the gp130/JAK2/STAT3 pathway. Wang et al. found miR-7 to target members of the mTOR signaling pathway (p70S6K, eIF4E, Mapkap1, Mknk1 and MknK2) [75], influencing cell proliferation, as well as MAP3K9 [76]. In addition, miR-7 targets also SET8, a histone methyltransferase, thus potentially influencing the expression of a number of downstream genes [56] (Table 4).
In colon cancer cells miR-7 also suppresses proliferation, increases apoptosis and causes cell-cycle arrest, by targeting YY1 and by influencing downstream p53, caspases and c-jun, as well as wnt signaling (through beta-catenin, survivin and FGF4) [13]. Neil, an endonuclease that inhibits apoptosis and increases cell survival and proliferation was found to be regulated by miR7 [113].
6. MiR-7 Roles in Lung Cancer
Promoter mutation of miR-7 was found to be associated with a poor prognosis of lung cancer [12]. The main targets released from miR-7 downregulation are those of EGFR and PIK3/Akt pathways, apoptosis inhibitors [109], and proteins involved in migration, FAK, PAK2 and NOVA2 [12,72,94,102,103]. PIK3/Akt signaling also connects TLR9 and miR-7 regulation [65]. However, Chou found that miR-7 could act as an oncomir in lung tumorigenesis [88]. EGFR, through the Ras/ERK/Myc pathway, increased the production of miR-7-1, which targets ERF, a transcriptional repressor. Therefore, in carcinoma samples, a positive correlation between EGFR and miR-7 expression was found, and miR-7 increased cell proliferation and tumor volume. Another example of oncogene activity was miR-7 modulation of the MYC pathway, in a positive feedback loop. The miRNA target is AMBRA1, an important regulator of early autophagy and a mediator in MYC dephosphorylation [158].
Hong et al. identified SMARCD1, a chromatin remodeling protein, to be a direct target of miR-7 in lung cancer cells [59]. They concluded that miR-7 influences the coupling of SMARCD1 with p53, which leads to an increased chemoresistance of lung cancer cells. miR-7 also downregulates PARP1, thus influencing DNA homologous recombination repair and survival after Adriamycin treatment of small cell lung cancer cells [121]. Furthermore, miR-7 modulates chemoresistance by targeting the multidrug resistance-associated protein MRP1/ABCC1 [125].
In addition to that, miR-7 was found to target several proteins linked to protein degradation as PA28gamma, a proteasome activator, targeted in non-small cell lung carcinoma [140]. O-GlcNAcyl Transferase (OGT), an enzyme involved in O-linked N-acetylglucosaminylation and contributing to cancer phenotype, is regulated by miR-7 [147]. In nasopharyngeal carcinoma cells, miR-7 was found to regulate the expression of enolase, ENO2, and therefore its downregulation can influence cell glycolysis [149].
7. MiR-7 Roles in Melanoma and Skin Cancer
Similarly to its role in other tissues, in melanoma cells, miR-7 takes part in the suppression of proliferation. However, as melanoma cells are not typically EGFR-driven, Giles et al. found miR-7 to target RelA and thus inhibit NF-κB activity and its downstream genes, such as IL-1β, IL-6 and IL-8 [70]. The analysis of melanoma patient samples revealed a correlation between RelA expression and poor survival.
On the contrary, Meza-Sosa et al. found KLF4 to be a miR-7 direct target in epithelial cells, and miR-7 overexpression in lung and skin epithelial cells enhanced cell proliferation, migration and tumorigenesis [84]. Tumors with an increased miR-7 had a decreased p21 and cyclin D. In thyroid papillary cancer, miR-7 targeted CKS2, a cyclin-dependent kinase regulator, and downstream cyclin B1 and cdk1 [89]. As a target, also PAK1 was detected [95].
It has been shown that in the cancer-associated fibroblasts of head and neck cancers, overexpression of miR-7 downregulates RSSF2, a proapoptotic molecule influencing proliferation and migration, and decreases the secretion of a tumor suppressor PAR-4 (prostate apoptosis response 4) [159]. In the human ocular tissue, miR-7 targets RGS5, a regulator of G protein signaling [139].
8. MiR-7 Roles in Breast, Prostate and Ovarian Cancer
In breast cancer miR-7 inhibits the metastases and influences epithelial-mesenchymal transition by targeting FAK, a kinase that acts as a mediator in ECM-integrin signaling [100]. Overexpression of miR-7 induces an increase in E cadherin and downregulation of mesenchymal proteins, suppresses proliferation, anchorage-independent growth, migration and invasion, as well as anchorage-independent growth in matrigel. The level of miR-7 is associated with the aggressiveness of estrogen receptor-positive breast tumors [160]. It also targets proteasome activator subunit 3 (REGγ) and contributes to the decrease in the cancer stem cell population survival, proliferation and migration [30,91,112,161]. Several miR-7 targets influence chemotherapy resistance, such as members of EGFR/PI3K signaling, BRCA1, LASP1, BCL-2 and MRP1 [104,162,163]. Okuda et al. (2013) found that miR-7 suppresses the ability of breast cancer stem cells to metastasize to the brain [85]. The correlation was found with miR-7 modulation of KLF4 expression, involved in stem cell biology. In addition, in a breast cancer cell line miR-7 was also found to be in regulation circuit with HOXD10, and, together with miR-218, to downregulate HoxB3 [69,120]. These changes were further connected with increased activity of other tumor suppressors, RASSF1A and Claudin-6 through epigenetic regulation, leading to cell cycle inhibition. Seong et al. found miR-7 to target REDD1, a negative regulator of mTOR signaling in the stress conditions [122]. miR-7 was therefore assigned to so-called hypoxamirs, miRNAs involved in hypoxic response. In HeLa cells, hypoxia caused downregulation of miR-7, in order to increase REDD1 level and inhibit mTOR signaling. In prostate cancer, it was found that miR-7 can regulate the expression of AXL, a receptor tyrosine kinase, associated with tumorigenesis, inhibition of apoptosis and EMT, often deregulated in different types of carcinomas [164]. miR-7 also inhibited the stemness of prostate stem cancer cells through repression of KLF4 and PI3K/Akt/p21 downstream pathway [165].
miR-7 overexpression in hamster ovary cell line CHO decreased the cell proliferation, without influencing viability. Transient transfection of CHO led to upregulation of nearly 200 genes and downregulation of around 350 genes. The pathways involved included translation, RNA and DNA processing, secretion and protein folding. miR-7 has been found to target regulators of G1-S transition, Skp2 and Psme, to upregulate p27KIP and arrest the cells in the G1 phase. Furthermore, it was found that miR-7 coordinately changes the levels of many genes in order to maintain homeostasis under the arrest conditions. It regulates Rad54L, a DNA repair protein, and influences the proapoptotic regulator p53 and the antiapoptotic Akt pathway to insure cell survival [82,166].
In testicular germ cell tumors, miR-7 was found to be one of the four hub miRNAs in regulatory networks of nonseminoma tumors [167].
In addition, expression of miR-7 was found increased in renal cell carcinomas in comparison with normal tissue, suggesting its activities as an oncogene [168].
9. MiR-7 Roles in Mesenchymal Tissue and Tumours
In osteosarcoma miR-7 influences IGF1R, and in paediatric rhabdomyosarcomas targets SLC25A37 and TIMM50, two mitochondrial proteins, important for the induction of cell death [79,111]. In osteosarcomas, miR-7 is supposed to be a regulating link between Linc00852 lncRNA, and AXL, a tyrosine kinase involved in tumor growth [169].
10. MiR-7 Roles in Leukaemia
miR-7 had a low expression in haematopoietic cells and in B-chronic lymphocytic leukaemia (Antica et al. unpublished results). In chronic myeloid leukaemia, it was found to interfere with Bcr/Abl signaling [170]. A higher expression of miR-7 was found in acute lymphocytic leukaemia (ALL) patients with CNS relapse compared to those without [171]. In B cell lymphoma it was found to be regulated by c-Myc [172]. In T-cell acute lymphocytic leukaemia (T-ALL), upregulation of long noncoding RNA ANRIL caused miR-7 sponging, binding multiple tandem miRNAs through response elements binding seed sequences, in order to sequester them from their target sequences. Consequently, TCF4, a miR-7 direct target, is upregulated and is involved in the disease progression [173]. In T-ALL, miR-7 was found to bind to TAL1, coding for T-cell acute lymphocytic leukaemia protein. In T-ALL, expression of miR-7 is often attenuated, while TAL1 expression is increased and solicitates cell proliferation [80]. In Non-Hodgkin lymphoma cells, miR-7 regulates migration and chemoresistance through KLF4 and YY1 [108] and miR-7 downregulation can increase the aggressiveness of follicular lymphoma by FasL upregulation in macrophages which modulate immunosuppressive stroma [174].
11. Conclusions
miR-7 is one of the most conserved and oldest miRs, and is engaged in numerous signaling circuits involved in differentiation, regulation of proliferation, apoptosis and migration. It targets numerous mRNAs depending on the intracellular milieu and is also regulated by different transcription factors and molecules involved in its processing and degradation. It was suggested that its role could be to buffer cellular processes under stress conditions and to coordinate cell proliferation with other functions. This could be the reason for its involvement in numerous diseases. In most tumors its expression is downregulated, as its dominant activity is tumor suppression by inhibition of cell proliferation and survival. In some cancer types, it acts as an oncomir, stressing the importance of nuances of signaling circuits in which it is involved. We believe that numerous functions in the maintenance of cell homeostasis and cell fate determination are still to be discovered.
Author Contributions
Conceptualization: P.K., M.A. and M.M.; writing—original draft preparation, M.M. and P.K.; writing—review and editing, M.A., P.K. and M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the University of ZagrebUniversity Support 20285121 for 2020 was given to M. Matulić.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank all the colleagues from the Department of Molecular Biology, Faculty of Science, University of Zagreb.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Treiber, T.; Treiber, N.; Meister, G. Regulation of microRNA biogenesis and its crosstalk with other cellular pathways. Nat. Rev. Mol. Cell Biol. 2019, 20, 5–20. [Google Scholar] [CrossRef]
- Oliveira, A.C.; Bovolenta, L.A.; Alves, L.; Figueiredo, L.; Ribeiro, A.O.; Campos, V.F.; Lemke, N.; Pinhal, D. Understanding the Modus Operandi of MicroRNA Regulatory Clusters. Cells 2019, 8, 1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Zhou, Y.; Guo, M.; Yue, D.; Chen, C.; Liang, G.; Xu, L. MicroRNA-7: Expression and function in brain physiological and pathological processes. Cell Biosci. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Cassidy, J.J.; Reinke, C.A.; Fischboeck, S.; Carthew, R.W. A MicroRNA Imparts Robustness against Environmental Fluctuation during Development. Cell 2009, 137, 273–282. [Google Scholar] [CrossRef] [Green Version]
- De Chevigny, A.; Coré, N.; Follert, P.; Gaudin, M.; Barbry, P.; Beclin, C.; Cremer, H. miR-7a regulation of Pax6 controls spatial origin of forebrain dopaminergic neurons. Nat. Neurosci. 2012, 15, 1120–1126. [Google Scholar] [CrossRef]
- Kredo-Russo, S.; Mandelbaum, A.D.; Ness, A.; Alon, I.; Lennox, K.A.; Behlke, M.A.; Hornstein, E. Pancreas-enriched miRNA refines endocrine cell differentiation. Development 2012, 139, 3021–3031. [Google Scholar] [CrossRef] [Green Version]
- Horsham, J.L.; Ganda, C.; Kalinowski, F.C.; Brown, R.A.; Epis, M.R.; Leedman, P.J. MicroRNA-7: A miRNA with expanding roles in development and disease. Int. J. Biochem. Cell Biol. 2015, 69, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, N.R.; Alves, F.D.L.; De Andrés-Aguayo, L.; Graf, T.; Cáceres, J.F.; Rappsilber, J.; Michlewski, G. Tissue-specific control of brain-enriched miR-7 biogenesis. Genes Dev. 2013, 27, 24–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valluy, J.; Bicker, S.; Aksoy-Aksel, A.; Lackinger, M.; Sumer, S.; Fiore, R.; Wüst, T.; Seffer, D.; Metge, F.; Dieterich, C.; et al. A coding-independent function of an alternative Ube3a transcript during neuronal development. Nat. Neurosci. 2015, 18, 666–673. [Google Scholar] [CrossRef]
- Caygill, E.E.; Brand, A.H. miR-7 Buffers Differentiation in the Developing Drosophila Visual System. Cell Rep. 2017, 20, 1255–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saydam, O.; Senol, O.; Würdinger, T.; Mizrak, A.; Ozdener, G.B.; Stemmer-Rachamimov, A.O.; Yi, M.; Stephens, R.M.; Krichevsky, A.M.; Saydam, N.; et al. miRNA-7 Attenuation in Schwannoma Tumors Stimulates Growth by Upregulating Three Oncogenic Signaling Pathways. Cancer Res. 2011, 71, 852–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Wang, K.; Liao, Z.; Li, Y.; Yang, H.; Chen, C.; Zhou, Y.; Tao, Y.; Guo, M.; Ren, T.; et al. Promoter mutation of tumor suppressor microRNA-7 is associated with poor prognosis of lung cancer. Mol. Clin. Oncol. 2015, 3, 1329–1336. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Li, X.; Wu, C.W.; Dong, Y.; Cai, M.; Mok, M.T.S.; Wang, H.; Chen, J.; Ng, S.S.M.; Chen, M.; et al. microRNA-7 is a novel inhibitor of YY1 contributing to colorectal tumorigenesis. Oncogene 2012, 32, 5078–5088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Midgley, A.; Morris, G.; Phillips, A.O.; Steadman, R. 17β-estradiol ameliorates age-associated loss of fibroblast function by attenuating IFN-γ/STAT1-dependent miR-7 upregulation. Aging Cell 2016, 15, 531–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzioni, M.; Agostinelli, L.; Candelaresi, C.; Saccomanno, S.; De Minicis, S.; Maroni, L.; Mingarelli, E.; Rychlicki, C.; Trozzi, L.; Banales, J.M.; et al. Activation of the developmental pathway neurogenin-3/microRNA-7a regulates cholangiocyte proliferation in response to injury. Hepatology 2014, 60, 1324–1335. [Google Scholar] [CrossRef]
- Chen, H.; Shalom-Feuerstein, R.; Riley, J.; Zhang, S.-D.; Tucci, P.; Agostini, M.; Aberdam, D.; Knight, R.A.; Genchi, G.; Nicotera, P.; et al. miR-7 and miR-214 are specifically expressed during neuroblastoma differentiation, cortical development and embryonic stem cells differentiation, and control neurite outgrowth in vitro. Biochem. Biophys. Res. Commun. 2010, 394, 921–927. [Google Scholar] [CrossRef]
- Xin, L.; Liu, L.; Liu, C.; Zhou, L.; Zhou, Q.; Yuan, Y.; Li, S.; Zhang, H. DNA-methylation-mediated silencing of miR-7-5p promotes gastric cancer stem cell invasion via increasing Smo and Hes1. J. Cell. Physiol. 2020, 235, 2643–2654. [Google Scholar] [CrossRef] [PubMed]
- Masuda, M.; Miki, Y.; Hata, S.; Takagi, K.; Sakurai, M.; Ono, K.; Suzuki, K.; Yang, Y.; Abe, E.; Hirakawa, H.; et al. An induction of microRNA, miR-7 through estrogen treatment in breast carcinoma. J. Transl. Med. 2012, 10, S2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duex, J.E.; Comeau, L.; Sorkin, A.; Purow, B.; Kefas, B. Usp18 Regulates Epidermal Growth Factor (EGF) Receptor Expression and Cancer Cell Survival via MicroRNA-7. J. Biol. Chem. 2011, 286, 25377–25386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-J.; Chien, P.-H.; Chen, W.-S.; Chien, Y.-F.; Hsu, Y.-Y.; Wang, L.-Y.; Chen, J.-Y.; Lin, C.-W.; Huang, T.-C.; Yu, Y.-L.; et al. Hepatitis B Virus-Encoded X Protein Downregulates EGFR Expression via Inducing MicroRNA-7 in Hepatocellular Carcinoma Cells. Evidence-Based Complement. Altern. Med. 2013, 2013, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, D.; Ham, J.; Park, S.; Lee, S.; Lee, H.; Kang, H.-S.; Kim, S.J. MicroRNA-7-5p mediates the signaling of hepatocyte growth factor to suppress oncogenes in the MCF-10A mammary epithelial cell. Sci. Rep. 2017, 7, 15425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, C.; Luo, X.; Duan, S.; Guo, L. Long noncoding RNA LINC00115 promotes breast cancer metastasis by inhibiting miR-7. FEBS Open Bio 2020, 10, 1230–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blume, C.J.; Hotzwagenblatt, A.; Hullein, J.; Sellner, L.; Jethwa, A.; Stolz, T.; Slabicki, M.; Lee, K.; Sharathchandra, A.; Benner, A.; et al. p53-dependent non-coding RNA networks in chronic lymphocytic leukemia. Leukemia 2015, 29, 2015–2023. [Google Scholar] [CrossRef]
- Pollock, A.; Bian, S.; Zhang, C.; Chen, Z.; Sun, T. Growth of the Developing Cerebral Cortex Is Controlled by MicroRNA-7 through the p53 Pathway. Cell Rep. 2014, 7, 1184–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gou, Q.; Wu, K.; Zhou, J.-K.; Xie, Y.; Liu, L.; Peng, Y. Profiling and bioinformatic analysis of circular RNA expression regulated by c-Myc. Oncotarget 2017, 8, 71587–71596. [Google Scholar] [CrossRef] [Green Version]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nat. Cell Biol. 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nat. Cell Biol. 2013, 495, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Rajman, M.; Schratt, G. MicroRNAs in neural development: From master regulators to fine-tuners. Development 2017, 144, 2310–2322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleaveland, B.; Shi, C.Y.; Stefano, J.; Bartel, D.P. A Network of Noncoding Regulatory RNAs Acts in the Mammalian Brain. Cell 2018, 174, 350–362. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Pan, M.; You, C.; Zhao, F.; Wu, D.; Guo, M.; Xu, H.; Shi, F.; Zheng, D.; Dou, J. MiR-7 reduces the BCSC subset by inhibiting XIST to modulate the miR-92b/Slug/ESA axis and inhibit tumor growth. Breast Cancer Res. 2020, 22, 1–14. [Google Scholar] [CrossRef]
- Ku, G.W.; Kang, Y.; Yu, S.-L.; Park, J.; Park, S.; Jeong, I.B.; Kang, M.W.; Son, J.W.; Kang, J. LncRNA LINC00240 suppresses invasion and migration in non-small cell lung cancer by sponging miR-7-5p. BMC Cancer 2021, 21, 1–13. [Google Scholar] [CrossRef]
- Liu, Y.; Du, Y.; Hu, X.; Zhao, L.; Xia, W. Up-regulation of ceRNA TINCR by SP1 contributes to tumorigenesis in breast cancer. BMC Cancer 2018, 18, 367. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Wang, D.; Shao, Y.; Zhang, T.; Xie, H.; Jiang, X.; Deng, Q.; Jiao, Y.; Yang, J.; Cai, C.; et al. SP1-induced lncRNA TINCR overexpression contributes to colorectal cancer progression by sponging miR-7-5p. Aging 2019, 11, 1389–1403. [Google Scholar] [CrossRef] [PubMed]
- Mo, D.; Liu, W.; Li, Y.; Cui, W. Long Non-coding RNA Zinc Finger Antisense 1 (ZFAS1) Regulates Proliferation, Migration, Invasion, and Apoptosis by Targeting MiR-7-5p in Colorectal Cancer. Med. Sci. Monit. 2019, 25, 5150–5158. [Google Scholar] [CrossRef]
- Zhang, X.; Niu, W.; Mu, M.; Hu, S.; Niu, C. Long non-coding RNA LPP-AS2 promotes glioma tumorigenesis via miR-7-5p/EGFR/PI3K/AKT/c-MYC feedback loop. J. Exp. Clin. Cancer Res. 2020, 39, 1–20. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, X.; Li, Y.; Zhou, Y.; Zhang, Z. Long noncoding RNA SOX21-AS1 promotes cervical cancer progression by competitively sponging miR-7/VDAC1. J. Cell. Physiol. 2019, 234, 17494–17504. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Yang, L.; Li, L.; Zeng, C. Long non-coding RNA KCNQ1OT1 modulates oxaliplatin resistance in hepatocellular carcinoma through miR-7-5p/ ABCC1 axis. Biochem. Biophys. Res. Commun. 2018, 503, 2400–2406. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Fu, Q.; Li, S.; Liang, N.; Li, F.; Li, C.; Sui, C.; Dionigi, G.; Sun, H. LncRNA FOXD2-AS1 Functions as a Competing Endogenous RNA to Regulate TERT Expression by Sponging miR-7-5p in Thyroid Cancer. Front. Endocrinol. 2019, 10. [Google Scholar] [CrossRef]
- Yang, Z.; Shi, X.; Li, C.; Wang, X.; Hou, K.; Li, Z.; Zhang, X.; Fan, Y.; Qu, X.; Che, X.; et al. Long non-coding RNA UCA1 upregulation promotes the migration of hypoxia-resistant gastric cancer cells through the miR-7-5p/EGFR axis. Exp. Cell Res. 2018, 368, 194–201. [Google Scholar] [CrossRef]
- Shu, L.; Zhang, W.; Huang, C.; Huang, G.; Su, G.; Xu, J. lncRNA ANRIL protects H9c2 cells against hypoxia-induced injury through targeting the miR-7-5p/SIRT1 axis. J. Cell. Physiol. 2020, 235, 1175–1183. [Google Scholar] [CrossRef]
- Zheng, Y.; Nie, P.; Xu, S. Long noncoding RNA CASC21 exerts an oncogenic role in colorectal cancer through regulating miR-7-5p/YAP1 axis. Biomed. Pharmacother. 2020, 121, 109628. [Google Scholar] [CrossRef]
- Arunkumar, G.; Anand, S.; Raksha, P.; Dhamodharan, S.; Rao, H.P.S.; Subbiah, S.; Murugan, A.K.; Munirajan, A.K. LncRNA OIP5-AS1 is overexpressed in undifferentiated oral tumors and integrated analysis identifies as a downstream effector of stemness-associated transcription factors. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Yang, X.; Li, N.-F.; Lin, L.; Luo, H. Circ_0015756 promotes proliferation, invasion and migration by microRNA-7-dependent inhibition of FAK in hepatocellular carcinoma. Cell Cycle 2019, 18, 2939–2953. [Google Scholar] [CrossRef] [PubMed]
- Zeng, K.; Chen, X.; Xu, M.; Liu, X.; Hu, X.; Xu, T.; Sun, H.; Pan, Y.; He, B.; Wang, S. CircHIPK3 promotes colorectal cancer growth and metastasis by sponging miR-7. Cell Death Dis. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Qi, X.; Zhang, X.; Fang, K.; Guo, Z.; Li, L. hsa_circRNA_0006528 as a competing endogenous RNA promotes human breast cancer progression by sponging miR-7-5p and activating the MAPK/ERK signaling pathway. Mol. Carcinog. 2019, 58, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Huang, M.; Cai, Y.; Yang, Y.; Sun, X.; Ke, Y. Circ-U2AF1 promotes human glioma via derepressing neuro-oncological ventral antigen 2 by sponging hsa-miR-7-5p. J. Cell. Physiol. 2019, 234, 9144–9155. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Z.; Hu, Y.; Zheng, W.; Tang, W.; Zhai, C.; Gu, Z.; Tao, J.; Wang, H. RETRACTED ARTICLE: Circ-TFCP2L1 Promotes the Proliferation and Migration of Triple Negative Breast Cancer through Sponging miR-7 by Inhibiting PAK1. J. Mammary Gland. Biol. Neoplasia 2019, 24, 323–331. [Google Scholar] [CrossRef]
- Wang, C.-J.; Gao, F.; Huang, Y.-J.; Han, D.-X.; Zheng, Y.; Wang, W.-H.; Jiang, H.; Gao, Y.; Yuan, B.; Zhang, J.-B. circAkap17b acts as a miR-7 family molecular sponge to regulate FSH secretion in rat pituitary cells. J. Mol. Endocrinol. 2020, 65, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Sang, Q.; Liu, X.; Wang, L.; Qi, L.; Sun, W.; Wang, W.; Sun, Y.; Zhang, H. CircSNCA downregulation by pramipexole treatment mediates cell apoptosis and autophagy in Parkinson’s disease by targeting miR-7. Aging 2018, 10, 1281–1293. [Google Scholar] [CrossRef]
- Chen, Y.; Guo, H.; Li, L.; Bao, D.; Gao, F.; Li, Q.; Huang, Q.; Duan, X.; Xiang, Z. Long Non-Coding RNA (lncRNA) Small Nucleolar RNA Host Gene 15 (SNHG15) Alleviates Osteoarthritis Progression by Regulation of Extracellular Matrix Homeostasis. Med. Sci. Monit. 2020, 26. [Google Scholar] [CrossRef]
- Wang, Y.; Vogel, G.; Yu, Z.; Richard, S. The QKI-5 and QKI-6 RNA Binding Proteins Regulate the Expression of MicroRNA 7 in Glial Cells. Mol. Cell. Biol. 2013, 33, 1233–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higuchi, T.; Todaka, H.; Sugiyama, Y.; Ono, M.; Tamaki, N.; Hatano, E.; Takezaki, Y.; Hanazaki, K.; Miwa, T.; Lai, S.; et al. Suppression of MicroRNA-7 (miR-7) Biogenesis by Nuclear Factor 90-Nuclear Factor 45 Complex (NF90-NF45) Controls Cell Proliferation in Hepatocellular Carcinoma. J. Biol. Chem. 2016, 291, 21074–21084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.-J.; Wang, C.-H.; Zhou, Y.; Liao, Z.-Y.; Zhu, S.-F.; Hu, Y.; Chen, C.; Luo, J.-M.; Wen, Z.-K.; Xu, L. TLR9 signaling repressed tumor suppressor miR-7 expression through up-regulation of HuR in human lung cancer cells. Cancer Cell Int. 2013, 13, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, A.; Shao, T.-J.; Ros, X.B.-D.; Lian, C.; Villanueva, P.; Dai, L.; Gu, S. AGO-bound mature miRNAs are oligouridylated by TUTs and subsequently degraded by DIS3L2. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Sun, S.; Tu, K.; Gao, Y.; Xie, B.; Krainer, A.R.; Zhu, J. A Splicing-Independent Function of SF2/ASF in MicroRNA Processing. Mol. Cell 2010, 38, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Yu, N.; Huangyang, P.; Yang, X.; Han, X.; Yan, R.; Jia, H.; Shang, Y.; Sun, L. microRNA-7 Suppresses the Invasive Potential of Breast Cancer Cells and Sensitizes Cells to DNA Damages by Targeting Histone Methyltransferase SET8. J. Biol. Chem. 2013, 288, 19633–19642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Cai, K.; Wang, J.; Wang, X.; Cheng, K.; Shi, F.; Jiang, L.; Zhang, Y.; Dou, J. MiR-7, Inhibited Indirectly by LincRNA HOTAIR, Directly Inhibits SETDB1 and Reverses the EMT of Breast Cancer Stem Cells by Downregulating the STAT3 Pathway. Stem Cells 2014, 32, 2858–2868. [Google Scholar] [CrossRef]
- Cheng, J.; Guo, S.; Chen, S.; Mastriano, S.J.; Liu, C.; D’Alessio, A.C.; Hysolli, E.; Guo, Y.; Yao, H.; Megyola, C.M.; et al. An Extensive Network of TET2-Targeting MicroRNAs Regulates Malignant Hematopoiesis. Cell Rep. 2013, 5, 471–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, C.-F.; Lin, S.-Y.; Chou, Y.-T.; Wu, C.-W. MicroRNA-7 Compromises p53 Protein-dependent Apoptosis by Controlling the Expression of the Chromatin Remodeling Factor SMARCD1. J. Biol. Chem. 2016, 291, 1877–1889. [Google Scholar] [CrossRef] [Green Version]
- López-Beas, J.; Capilla-González, V.; Aguilera, Y.; Mellado, N.; Lachaud, C.C.; Martín, F.; Smani, T.; Soria, B.; Hmadcha, A. miR-7 Modulates hESC Differentiation into Insulin-Producing Beta-like Cells and Contributes to Cell Maturation. Mol. Ther. Nucleic Acids 2018, 12, 463–477. [Google Scholar] [CrossRef]
- Titze-De-Almeida, R.; Titze-De-Almeida, S.S. miR-7 Replacement Therapy in Parkinson’s Disease. Curr. Gene Ther. 2018, 18, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Kefas, B.; Godlewski, J.; Comeau, L.; Li, Y.; Abounader, R.; Hawkinson, M.; Lee, J.; Fine, H.; Chiocca, E.A.; Lawler, S.; et al. microRNA-7 Inhibits the Epidermal Growth Factor Receptor and the Akt Pathway and Is Down-regulated in Glioblastoma. Cancer Res. 2008, 68, 3566–3572. [Google Scholar] [CrossRef] [Green Version]
- Webster, R.J.; Giles, K.M.; Price, K.J.; Zhang, P.M.; Mattick, J.S.; Leedman, P.J. Regulation of Epidermal Growth Factor Receptor Signaling in Human Cancer Cells by MicroRNA-7. J. Biol. Chem. 2009, 284, 5731–5741. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Li, J.; Sun, Y.; Zhang, Y.; Dong, L.; Shen, C.; Yang, L.; Yang, M.; Li, Y.; Shen, G.; et al. miR-7 reverses the resistance to BRAFi in melanoma by targeting EGFR/IGF-1R/CRAF and inhibiting the MAPK and PI3K/AKT signaling pathways. Oncotarget 2016, 7, 53558–53570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Wen, Z.; Zhou, Y.; Liu, Z.; Li, Q.; Fei, G.; Luo, J.; Ren, T. MicroRNA-7–regulated TLR9 signaling–enhanced growth and metastatic potential of human lung cancer cells by altering the phosphoinositide-3-kinase, regulatory subunit 3/Akt pathway. Mol. Biol. Cell 2013, 24, 42–55. [Google Scholar] [CrossRef]
- Fang, Y.; Xue, J.-L.; Shen, Q.; Chen, J.; Tian, L. MicroRNA-7 inhibits tumor growth and metastasis by targeting the phosphoinositide 3-kinase/Akt pathway in hepatocellular carcinoma. Hepatology 2012, 55, 1852–1862. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Jiang, Z.; Huang, J.; Huang, S.; Li, Y.; Yu, S.; Yu, S.; Liu, X. miR-7 inhibits glioblastoma growth by simultaneously interfering with the PI3K/ATK and Raf/MEK/ERK pathways. Int. J. Oncol. 2014, 44, 1571–1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Zhang, P.; Chen, Z.; Liu, M.; Li, X.; Tang, H. MicroRNA-7 downregulates XIAP expression to suppress cell growth and promote apoptosis in cervical cancer cells. FEBS Lett. 2013, 587, 2247–2253. [Google Scholar] [CrossRef] [Green Version]
- Reddy, S.D.N.; Ohshiro, K.; Rayala, S.K.; Kumar, R. MicroRNA-7, a Homeobox D10 Target, Inhibits p21-Activated Kinase 1 and Regulates Its Functions. Cancer Res. 2008, 68, 8195–8200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giles, K.M.; Brown, R.A.; Epis, M.R.; Kalinowski, F.C.; Leedman, P.J. miRNA-7-5p inhibits melanoma cell migration and invasion. Biochem. Biophys. Res. Commun. 2013, 430, 706–710. [Google Scholar] [CrossRef]
- Jiang, L.; Liu, X.; Chen, Z.; Jin, Y.; Heidbreder, C.E.; Kolokythas, A.; Wang, A.; Dai, Y.; Zhou, X. MicroRNA-7 targets IGF1R (insulin-like growth factor 1 receptor) in tongue squamous cell carcinoma cells. Biochem. J. 2010, 432, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Rai, K.; Takigawa, N.; Ito, S.; Kashihara, H.; Ichihara, E.; Yasuda, T.; Shimizu, K.; Tanimoto, M.; Kiura, K. Liposomal Delivery of MicroRNA-7–Expressing Plasmid Overcomes Epidermal Growth Factor Receptor Tyrosine Kinase Inhibitor-Resistance in Lung Cancer Cells. Mol. Cancer Ther. 2011, 10, 1720–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalinowski, F.C.; Brown, R.A.; Ganda, C.; Giles, K.M.; Epis, M.R.; Horsham, J.; Leedman, P.J. microRNA-7: A tumor suppressor miRNA with therapeutic potential. Int. J. Biochem. Cell Biol. 2014, 54, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.-D.; Lu, Y.-Y.; Guo, H.; Xie, H.-H.; He, L.-J.; Shen, G.-F.; Zhou, J.-F.; Li, T.; Hu, S.-J.; Zhou, L.; et al. MicroRNA-7/NF-κB signaling regulatory feedback circuit regulates gastric carcinogenesis. J. Cell Biol. 2015, 210, 613–627. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, Q.; Song, J. Inhibition of autophagy potentiates the proliferation inhibition activity of microRNA-7 in human hepatocellular carcinoma cells. Oncol. Lett. 2017, 14, 3566–3572. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Cao, T.; Ma, C.; Shi, Y.; Sun, Y.; Wang, Z.P.; Ma, J. miR-7 Suppresses Tumor Progression by Directly Targeting MAP3K9 in Pancreatic Cancer. Mol. Ther. Nucleic Acids 2018, 13, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Dou, W.; He, L.; Liang, S.; Tie, J.; Liu, C.; Li, T.; Lu, Y.; Mo, P.; Shi, Y.; et al. MicroRNA-7 functions as an anti-metastatic microRNA in gastric cancer by targeting insulin-like growth factor-1 receptor. Oncogene 2013, 32, 1363–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Sun, F.; Dong, N.; Sun, Z.; Diao, Y.; Zheng, C.; Sun, J.; Yang, Y.; Jiang, D. MicroRNA-7 directly targets insulin-like growth factor 1 receptor to inhibit cellular growth and glucose metabolism in gliomas. Diagn. Pathol. 2014, 9, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Zhao, M.; Wang, G. Upregulation of microRNA-7 contributes to inhibition of the growth and metastasis of osteosarcoma cells through the inhibition of IGF1R. J. Cell. Physiol. 2019, 234, 22195–22206. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, Z.; Luo, W.; Liu, J.; Lou, Y.; Xia, S. MiR-7 Functions as a Tumor Suppressor by Targeting the Oncogenes TAL1 in T-Cell Acute Lymphoblastic Leukemia. Technol. Cancer Res. Treat. 2020, 19. [Google Scholar] [CrossRef]
- Giles, K.M.; Brown, R.A.; Ganda, C.; Podgorny, M.J.; Candy, P.A.; Wintle, L.C.; Richardson, K.L.; Kalinowski, F.C.; Stuart, L.M.; Epis, M.R.; et al. microRNA-7-5p inhibits melanoma cell proliferation and metastasis by suppressing RelA/NF-κB. Oncotarget 2016, 7, 31663–31680. [Google Scholar] [CrossRef] [Green Version]
- Meleady, P.; Gallagher, M.; Clarke, C.; Henry, M.; Sanchez, N.; Barron, N.; Clynes, M. Impact of miR-7 over-expression on the proteome of Chinese hamster ovary cells. J. Biotechnol. 2012, 160, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hu, S.; Zhang, X.; Wang, L.; Zhang, X.; Yan, B.; Zhao, J.; Yang, A.; Zhang, R. MicroRNA-7 arrests cell cycle in G1 phase by directly targeting CCNE1 in human hepatocellular carcinoma cells. Biochem. Biophys. Res. Commun. 2014, 443, 1078–1084. [Google Scholar] [CrossRef]
- Meza-Sosa, K.F.; Pérez-García, E.I.; Camacho-Concha, N.; López-Gutiérrez, O.; Pedraza-Alva, G.; Pérez-Martínez, L. MiR-7 Promotes Epithelial Cell Transformation by Targeting the Tumor Suppressor KLF4. PLoS ONE 2014, 9, e103987. [Google Scholar] [CrossRef] [Green Version]
- Okuda, H.; Xing, F.; Pandey, P.R.; Sharma, S.; Watabe, M.; Pai, S.K.; Mo, Y.-Y.; Iiizumi-Gairani, M.; Hirota, S.; Liu, Y.; et al. miR-7 Suppresses Brain Metastasis of Breast Cancer Stem-Like Cells By Modulating KLF4. Cancer Res. 2013, 73, 1434–1444. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.-Z.; Wen, L.; Wei, X.; Wang, Q.-R.; Xu, L.-W.; Zhang, H.-M.; Liu, W.-C. Inhibition of miR-7 promotes angiogenesis in human umbilical vein endothelial cells by upregulating VEGF via KLF4. Oncol. Rep. 2016, 36, 1569–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.M.; Phillips, B.L.; Patel, R.S.; Cohen, D.M.; Jakymiw, A.; Kong, W.W.; Cheng, J.Q.; Chan, E.K.L. Keratinization-associated miR-7 and miR-21 Regulate Tumor Suppressor Reversion-inducing Cysteine-rich Protein with Kazal Motifs (RECK) in Oral Cancer*. J. Biol. Chem. 2012, 287, 29261–29272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, Y.-T.; Lin, H.-H.; Lien, Y.-C.; Wang, Y.-H.; Hong, C.-F.; Kao, Y.-R.; Lin, S.-C.; Chang, Y.-C.; Lin, S.-Y.; Chen, S.-J.; et al. EGFR Promotes Lung Tumorigenesis by Activating miR-7 through a Ras/ERK/Myc Pathway That Targets the Ets2 Transcriptional Repressor ERF. Cancer Res. 2010, 70, 8822–8831. [Google Scholar] [CrossRef] [Green Version]
- Hua, K.; Jin, J.; Zhang, H.; Zhao, B.; Wu, C.; Xu, H.; Fang, L. MicroRNA-7 inhibits proliferation, migration and invasion of thyroid papillary cancer cells via targeting CKS2. Int. J. Oncol. 2016, 49, 1531–1540. [Google Scholar] [CrossRef] [Green Version]
- Ling, Y.; Cao, C.; Li, S.; Qiu, M.; Shen, G.; Chen, Z.; Yao, F.; Chen, W. TRIP6, as a target of miR-7, regulates the proliferation and metastasis of colorectal cancer cells. Biochem. Biophys. Res. Commun. 2019, 514, 231–238. [Google Scholar] [CrossRef]
- Pan, M.; Li, M.; You, C.; Zhao, F.; Guo, M.; Xu, H.; Li, L.; Wang, L.; Dou, J. Inhibition of breast cancer growth via miR-7 suppressing ALDH1A3 activity concomitant with decreasing breast cancer stem cell subpopulation. J. Cell. Physiol. 2019, 235, 1405–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.; Qi, Y.; Shao, L.; Liu, M.; Li, X.; Tang, H. Downregulation of miR-7 upregulates Cullin 5 (CUL5) to facilitate G1/S transition in human hepatocellular carcinoma cells. IUBMB Life 2013, 65, 1026–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, A.; Qian, W. MicroRNA-7 inhibits colorectal cancer cell proliferation, migration and invasion via TYRO3 and phosphoinositide 3-kinase/protein B kinase/mammalian target of rapamycin pathway suppression. Int. J. Mol. Med. 2018, 42, 2503–2514. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wu, X.; Guo, L.; Shi, J.; Li, J. MicroRNA-7-5p induces cell growth inhibition, cell cycle arrest and apoptosis by targeting PAK2 in non-small cell lung cancer. FEBS Open Bio 2019, 9, 1983–1993. [Google Scholar] [CrossRef] [Green Version]
- Yue, K.; Wang, X.; Wu, Y.; Zhou, X.; He, Q.; Duan, Y. microRNA-7 regulates cell growth, migration and invasion via direct targeting of PAK1 in thyroid cancer. Mol. Med. Rep. 2016, 14, 2127–2134. [Google Scholar] [CrossRef]
- Pan, C.-M.; Chan, K.-H.; Chen, C.-H.; Jan, C.-I.; Liu, M.-C.; Lin, C.-M.; Cho, D.-Y.; Tsai, W.-C.; Chu, Y.-T.; Cheng, C.-H.; et al. MicroRNA-7 targets T-Box 2 to inhibit epithelial-mesenchymal transition and invasiveness in glioblastoma multiforme. Cancer Lett. 2020, 493, 133–142. [Google Scholar] [CrossRef]
- Shukla, A.; Gupta, P.; Singh, R.; Mishra, D.P. Glycolytic inhibitor 2-Deoxy-d-Glucose activates migration and invasion in glioblastoma cells through modulation of the miR-7-5p/TFF3 signaling pathway. Biochem. Biophys. Res. Commun. 2018, 499, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Hao, S.; Huang, Z.; Wang, W.; Yan, P.; Zhou, W.; Zhu, Q.; Liu, X. MiR-7 inhibited peripheral nerve injury repair by affecting neural stem cells migration and proliferation through cdc42. Mol. Pain 2018, 14. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.-G.; Wang, Y.-Y.; Fan, L.-G.; Luo, H.; Han, B.; Sun, L.-H.; Wang, X.-F.; Zhang, J.-X.; Cao, L.; Wang, X.-R.; et al. MicroRNA-7 regulates glioblastoma cell invasion via targeting focal adhesion kinase expression. Chin. Med. J. 2011, 124, 2616–2621. [Google Scholar] [PubMed]
- Kong, X.; Li, G.; Yuan, Y.; He, Y.; Wu, X.; Zhang, W.; Wu, Z.; Chen, T.; Wu, W.; Lobie, P.E.; et al. MicroRNA-7 Inhibits Epithelial-to-Mesenchymal Transition and Metastasis of Breast Cancer Cells via Targeting FAK Expression. PLoS ONE 2012, 7, e41523. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.-Y.; Zhan, Y.-S.; Huang, J.; Chen, Y.-X. MicroRNA-7 suppresses human colon cancer invasion and proliferation by targeting the expression of focal adhesion kinase. Mol. Med. Rep. 2015, 13, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Cao, Q.; Mao, Z.-D.; Shi, Y.-J.; Chen, Y.; Sun, Y.; Zhang, Q.; Song, L.; Peng, L.-P. MicroRNA-7 inhibits cell proliferation, migration and invasion in human non-small cell lung cancer cells by targeting FAK through ERK/MAPK signaling pathway. Oncotarget 2016, 7, 77468–77481. [Google Scholar] [CrossRef]
- Xiao, H. MiR-7-5p suppresses tumor metastasis of non-small cell lung cancer by targeting NOVA2. Cell. Mol. Biol. Lett. 2019, 24, 1–13. [Google Scholar] [CrossRef]
- Moazzeni, H.; Najafi, A.; Khani, M. Identification of direct target genes of miR-7, miR-9, miR-96, and miR-182 in the human breast cancer cell lines MCF-7 and MDA-MB-231. Mol. Cell. Probes 2017, 34, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Kong, W.; Jiang, J.; Xu, H.; Zhao, W. miR-7-5p inhibits cell migration and invasion in glioblastoma through targeting SATB1. Oncol. Lett. 2018, 17, 1819–1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, Y.; Shen, W.; Min, M.; Liu, Y. MicroRNA-7 functions as a tumor-suppressor gene by regulating ILF2 in pancreatic carcinoma. Int. J. Mol. Med. 2017, 39, 900–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, S.; Jin, S.; Wang, C.; Tu, P.; Hu, K.; Lu, J. Androgen receptor suppresses vasculogenic mimicry in hepatocellular carcinoma via circRNA7/miRNA7-5p/VE-cadherin/Notch4 signalling. J. Cell. Mol. Med. 2020, 24, 14110–14120. [Google Scholar] [CrossRef]
- Morales-Martinez, M.; Vega, G.G.; Neri, N.; Nambo, M.J.; Alvarado, I.; Cuadra, I.; Duran-Padilla, M.A.; Huerta-Yepez, S.; Vega, M.I. MicroRNA-7 Regulates Migration and Chemoresistance in Non-Hodgkin Lymphoma Cells Through Regulation of KLF4 and YY1. Front. Oncol. 2020, 10, 588893. [Google Scholar] [CrossRef]
- Xiong, S.; Zheng, Y.; Jiang, P.; Liu, R.; Liu, X.; Chu, Y. MicroRNA-7 Inhibits the Growth of Human Non-Small Cell Lung Cancer A549 Cells through Targeting BCL-2. Int. J. Biol. Sci. 2011, 7, 805–814. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, X.; Hu, S.; Zheng, M.; Zhang, J.; Zhao, J.; Zhang, X.; Yan, B.; Jia, L.; Zhao, J.; et al. Identification of miRNA-7 by genome-wide analysis as a critical sensitizer for TRAIL-induced apoptosis in glioblastoma cells. Nucleic Acids Res. 2017, 45, 5930–5944. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Kong, D.; He, M.; Gong, J.; Nie, Y.; Tai, S.; Teng, C.-B. MiR-7 mediates mitochondrial impairment to trigger apoptosis and necroptosis in Rhabdomyosarcoma. Biochim. Biophys. Acta (BBA) Bioenerg. 2020, 1867, 118826. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Luo, X.; Li, P.; Tan, J.; Wang, X.; Xiang, T.; Ren, G. miR-7-5p suppresses cell proliferation and induces apoptosis of breast cancer cells mainly by targeting REGγ. Cancer Lett. 2015, 358, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Liu, Y.; Xin, N.; Miao, J.; Du, J.; Wang, Y.; Shi, H.; Wei, Y.; Zhang, H.; Chen, Y.; et al. Nei Endonuclease VIII-Like1 (NEIL1) Inhibits Apoptosis of Human Colorectal Cancer Cells. BioMed Res. Int. 2020, 2020, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Alexandrov, P.N.; Jaber, V.; Lukiw, W.J. Deficiency in the Ubiquitin Conjugating Enzyme UBE2A in Alzheimer’s Disease (AD) is Linked to Deficits in a Natural Circular miRNA-7 Sponge (circRNA; ciRS-7). Genes 2016, 7, 116. [Google Scholar] [CrossRef] [Green Version]
- Jia, B.; Liu, W.; Gu, J.; Wang, J.; Lv, W.; Zhang, W.; Hao, Q.; Pang, Z.; Mu, N.; Zhang, W.; et al. MiR-7-5p suppresses stemness and enhances temozolomide sensitivity of drug-resistant glioblastoma cells by targeting Yin Yang 1. Exp. Cell Res. 2019, 375, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wang, B. MiR-7-5p enhances cerebral ischemia-reperfusion injury by degrading sirt1 mRNA. J. Cardiovasc. Pharmacol. 2020, 76, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, A.D.; Choi, D.C.; Kabaria, S.; Tran, A.; Junn, E. MicroRNA-7 Regulates the Function of Mitochondrial Permeability Transition Pore by Targeting VDAC1 Expression. J. Biol. Chem. 2016, 291, 6483–6493. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Qiang, Y.; Zhu, L.; Jiang, Y.; Wang, Y.; Shao, X.; Yin, L.; Chen, J.; Chen, Z. MicroRNA-7 downregulates the oncogene VDAC1 to influence hepatocellular carcinoma proliferation and metastasis. Tumor Biol. 2016, 37, 10235–10246. [Google Scholar] [CrossRef] [PubMed]
- Kabaria, S.; Choi, D.C.; Chaudhuri, A.D.; Jain, M.R.; Li, H.; Junn, E. MicroRNA-7 activates Nrf2 pathway by targeting Keap1 expression. Free Radic. Biol. Med. 2015, 89, 548–556. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Zhu, F.; Chen, P. miR-7 and miR-218 epigenetically control tumor suppressor genes RASSF1A and Claudin-6 by targeting HoxB3 in breast cancer. Biochem. Biophys. Res. Commun. 2012, 424, 28–33. [Google Scholar] [CrossRef]
- Lai, J.; Yang, H.; Zhu, Y.; Ruan, M.; Huang, Y.; Zhang, Q. MiR-7-5p-mediated downregulation of PARP1 impacts DNA homologous recombination repair and resistance to doxorubicin in small cell lung cancer. BMC Cancer 2019, 19, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Seong, M.; Lee, J.; Kang, H. Hypoxia-induced regulation of mTOR signaling by miR-7 targeting REDD1. J. Cell. Biochem. 2019, 120, 4523–4532. [Google Scholar] [CrossRef]
- Xu, K.; Song, X.; Chen, Z.; Qin, C. miR-7 inhibits colorectal cancer cell proliferation and induces apoptosis by targeting XRCC2. OncoTargets Ther. 2014, 7, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Downing, S.; Zhang, F.; Chen, Z.; Tzanakakis, E.S. MicroRNA-7 directly targets Reg1 in pancreatic cells. Am. J. Physiol. Physiol. 2019, 317, C366–C374. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wu, X.; Huang, J.; Peng, J.; Guo, L. miR-7 modulates chemoresistance of small cell lung cancer by repressing MRP1/ABCC1. Int. J. Exp. Pathol. 2015, 96, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.D.; Fan, Q.Y.; Qiu, Z.; Chen, S. MiR-7 alleviates secondary inflammatory response of microglia caused by cerebral hem-orrhage through inhibiting TLR4 expression. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5597–5604. [Google Scholar] [CrossRef]
- Chen, H.; Guo, M.; Yue, D.; Zhao, J.; Zhou, Y.; Chen, C.; Liang, G.; Xu, L. MicroRNA-7 negatively regulates Toll-like receptor 4 signaling pathway through FAM177A. Immunology 2021, 162, 44–57. [Google Scholar] [CrossRef]
- Fan, Z.; Lu, M.; Qiao, C.; Zhou, Y.; Ding, J.-H.; Hu, G. MicroRNA-7 Enhances Subventricular Zone Neurogenesis by Inhibiting NLRP3/Caspase-1 Axis in Adult Neural Stem Cells. Mol. Neurobiol. 2016, 53, 7057–7069. [Google Scholar] [CrossRef]
- Luo, J.; Li, H.; Zhang, C. MicroRNA-7 inhibits the malignant phenotypes of non-small cell lung cancer in vitro by targeting Pax6. Mol. Med. Rep. 2015, 12, 5443–5448. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Mubarak, T.; Chen, Y.; Lee, T.; Pollock, A.; Sun, T. Counter-Balance Between Gli3 and miR-7 Is Required for Proper Morphogenesis and Size Control of the Mouse Brain. Front. Cell. Neurosci. 2018, 12, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Qiu, M.; An, Y.; Huang, J.; Gong, C. miR-7-5p acts as a tumor suppressor in bladder cancer by regulating the hedgehog pathway factor Gli3. Biochem. Biophys. Res. Commun. 2018, 503, 2101–2107. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Chen, M.; Wang, B.; Han, Y.; Shang, H.; Chen, J. miR-7-5p Promotes Hepatic Stellate Cell Activation by Targeting Fibroblast Growth Factor Receptor 4. Gastroenterol. Res. Pr. 2020, 2020, 1–9. [Google Scholar] [CrossRef]
- Kim, T.; Mehta, S.L.; Morris-Blanco, K.C.; Chokkalla, A.K.; Chelluboina, B.; Lopez, M.; Sullivan, R.; Kim, H.T.; Cook, T.D.; Kim, J.Y.; et al. The microRNA miR-7a-5p ameliorates ischemic brain damage by repressing α-synuclein. Sci. Signal. 2018, 11, eaat4285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adusumilli, L.; Facchinello, N.; Teh, C.; Busolin, G.; Le, M.T.; Yang, H.; Beffagna, G.; Campanaro, S.; Tam, W.L.; Argenton, F.; et al. miR-7 Controls the Dopaminergic/Oligodendroglial Fate through Wnt/β-catenin Signaling Regulation. Cells 2020, 9, 711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.-Y.; Pang, K.; Kim, J.Y.; Ryu, J.R.; Kang, H.; Liu, Z.; Kim, W.-K.; Sun, W.; Kim, H.; Han, K. Post-transcriptional regulation of SHANK3 expression by microRNAs related to multiple neuropsychiatric disorders. Mol. Brain 2015, 8, 74. [Google Scholar] [CrossRef] [Green Version]
- Da Ros, V.G.; Gutierrez-Perez, I.; Ferres-Marco, L.; Dominguez, M. Dampening the Signals Transduced through Hedgehog via MicroRNA miR-7 Facilitates Notch-Induced Tumourigenesis. PLoS Biol. 2013, 11, e1001554. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Xu, T.; Li, Y.; Fei, W.; Yang, G.; Hong, Y. Inhibition of CRY2 by STAT3/miRNA-7-5p Promotes Osteoblast Differentiation through Upregulation of CLOCK/BMAL1/P300 Expression. Mol. Ther. Nucleic Acids 2020, 19, 865–876. [Google Scholar] [CrossRef]
- Yu, X.; Li, M.; Cui, M.; Sun, B.; Zhou, Z. Silence of yki by miR-7 regulates the Hippo pathway. Biochem. Biophys. Res. Commun. 2020, 532, 446–452. [Google Scholar] [CrossRef]
- Banaei-Esfahani, A.; Moazzeni, H.; Nosar, P.N.; Amin, S.; Arefian, E.; Soleimani, M.; Yazdani, S.; Elahi, E. MicroRNAs that target RGS5. Iran. J. Basic Med. Sci. 2015, 18, 108–114. [Google Scholar]
- Xiong, S.; Zheng, Y.; Jiang, P.; Liu, R.; Liu, X.; Qian, J.; Gu, J.; Chang, L.; Ge, D.; Chu, Y. PA28gamma emerges as a novel functional target of tumour suppressor microRNA-7 in non-small-cell lung cancer. Br. J. Cancer 2013, 110, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Frutos, M.F.-D.; Galán-Chilet, I.; Goedeke, L.; Kim, B.; Pardo-Marqués, V.; Pérez-García, A.; Herrero, J.I.; Fernández-Hernando, C.; Kim, J.; Ramírez, C.M. MicroRNA 7 Impairs Insulin Signaling and Regulates Aβ Levels through Posttranscriptional Regulation of the Insulin Receptor Substrate 2, Insulin Receptor, Insulin-Degrading Enzyme, and Liver X Receptor Pathway. Mol. Cell. Biol. 2019, 39. [Google Scholar] [CrossRef]
- Miyazawa, M.; Bogdan, A.R.; Hashimoto, K.; Tsuji, Y. Regulation of transferrin receptor-1 mRNA by the interplay between IRE-binding proteins and miR-7/miR-141 in the 3′-IRE stem–loops. RNA 2018, 24, 468–479. [Google Scholar] [CrossRef] [Green Version]
- Matarese, A.; Gambardella, J.; Lombardi, A.; Wang, X.; Santulli, G. miR-7 Regulates GLP-1-Mediated Insulin Release by Targeting β-Arrestin 1. Cells 2020, 9, 1621. [Google Scholar] [CrossRef]
- Dewing, A.S.; Rueli, R.H.; Robles, M.J.; Nguyen-Wu, E.D.; Zeyda, T.; Berry, M.J.; Bellinger, F.P. Expression and regulation of mouse selenoprotein P transcript variants differing in non-coding RNA. RNA Biol. 2012, 9, 1361–1369. [Google Scholar] [CrossRef] [Green Version]
- Yuan, B.; Sun, G.; Zhang, G.; Wu, J.; Xu, C.; Dai, L.; Chen, J.; Yu, X.; Zhao, Z.; Zhang, J. Identification of target genes for adenohypophysis-prefer miR-7 and miR-375 in cattle. Genet. Mol. Res. 2015, 14, 9753–9763. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, K.; Lapierre, M.P.; Gasser, E.; Denzler, R.; Yang, Y.; Ruelicke, T.; Kero, J.; Latreille, M.; Stoffel, M. Loss of microRNA-7a2 induces hypogonadotropic hypogonadism and infertility. J. Clin. Investig. 2017, 127, 1061–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, S.Y.; Lee, S.Y.; Yu, S.-L.; Park, S.J.; Kang, D.; Kim, J.S.; Jeong, I.B.; Kwon, S.J.; Hwang, W.J.; Park, C.R.; et al. MicroRNA-7-5p′s role in the O-GlcNAcylation and cancer metabolism. Non-coding RNA Res. 2020, 5, 201–207. [Google Scholar] [CrossRef]
- Ma, C.; Gu, R.; Wang, X.; He, S.; Bai, J.; Zhang, L.; Zhang, J.; Li, Q.; Qu, L.; Xin, W.; et al. circRNA CDR1as Promotes Pulmonary Artery Smooth Muscle Cell Calcification by Upregulating CAMK2D and CNN3 via Sponging miR-7-5p. Mol. Ther. Nucleic Acids 2020, 22, 530–541. [Google Scholar] [CrossRef]
- Peng, J.; Liu, F.; Zheng, H.; Wu, Q.; Liu, S. IncRNA ZFAS1 contributes to the radioresistance of nasopharyngeal carcinoma cells by sponging hsa-miR-7-5p to upregulate ENO2. Cell Cycle 2021, 20, 126–141. [Google Scholar] [CrossRef]
- Jin, H.-F.; Wang, J.-F.; Shao, M.; Zhou, K.; Ma, X.; Lv, X.-P. Down-Regulation of miR-7 in Gastric Cancer Is Associated With Elevated LDH-A Expression and Chemoresistance to Cisplatin. Front. Cell Dev. Biol. 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Lapierre, M.P.; Godbersen, S.; Esteban, M.T.; Schad, A.N.; Treier, M.; Ghoshdastider, U.; Stoffel, M. MicroRNA-7a2 Regulates Prolactin in Developing Lactotrophs and Prolactinoma Cells. Endocrinology 2021, 162. [Google Scholar] [CrossRef]
- Zhang, G.; Chen, F.; Wu, P.; Li, T.; He, M.; Yin, X.; Shi, H.; Duan, Y.; Zhang, T.; Wang, J.; et al. MicroRNA-7 Targets the KLF4 Gene to Regulate the Proliferation and Differentiation of Chicken Primary Myoblasts. Front. Genet. 2020, 11, 842. [Google Scholar] [CrossRef] [PubMed]
- Junn, E.; Lee, K.-W.; Jeong, B.S.; Chan, T.W.; Im, J.-Y.; Mouradian, M.M. Repression of -synuclein expression and toxicity by microRNA-7. Proc. Natl. Acad. Sci. USA 2009, 106, 13052–13057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diana, A.; Gaido, G.; Murtas, D. MicroRNA Signature in Human Normal and Tumoral Neural Stem Cells. Int. J. Mol. Sci. 2019, 20, 4123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Liu, Y.; Li, L.; Xu, Z.; Bi, B.; Wang, Y.; Li, J.Y. MiR-7-5p is frequently downregulated in glioblastoma microvasculature and inhibits vascular endothelial cell proliferation by targeting RAF1. Tumor Biol. 2014, 35, 10177–10184. [Google Scholar] [CrossRef]
- Kolic, D.; Horvat, L.; Setinc, M.; Antica, M.; Matulic, M. miR-7 AND miR-34a sequence cloning and expression in a1235 glioblastoma cell line. Mol. Exp. Boil. Med. 2020, 3, 31–38. [Google Scholar] [CrossRef]
- Chen, Y.A.; Cheng, L.; Zhang, Y.; Peng, L.; Yang, H.G. LncRNA RUSC1-AS1 promotes the proliferation of hepatocellular carcinoma cells through modulating NOTCH signaling. Neoplasma 2021, 67, 1204–1213. [Google Scholar] [CrossRef]
- Capizzi, M.; Strappazzon, F.; Cianfanelli, V.; Papaleo, E.; Cecconi, F. MIR7–3HG, a MYC-dependent modulator of cell proliferation, inhibits autophagy by a regulatory loop involving AMBRA1. Autophagy 2017, 13, 554–566. [Google Scholar] [CrossRef] [Green Version]
- Shen, Z.; Qin, X.; Yan, M.; Li, R.; Chen, G.; Zhang, J.; Chen, W. Cancer-associated fibroblasts promote cancer cell growth through a miR-7-RASSF2-PAR-4 axis in the tumor microenvironment. Oncotarget 2016, 8, 1290–1303. [Google Scholar] [CrossRef] [Green Version]
- Foekens, J.A.; Sieuwerts, A.M.; Smid, M.; Look, M.P.; De Weerd, V.; Boersma, A.W.M.; Klijn, J.G.M.; Wiemer, E.A.C.; Martens, J.W. Four miRNAs associated with aggressiveness of lymph node-negative, estrogen receptor-positive human breast cancer. Proc. Natl. Acad. Sci. USA 2008, 105, 13021–13026. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Pan, M.; Wang, J.; You, C.; Zhao, F.; Zheng, D.; Guo, M.; Xu, H.; Wu, D.; Wang, L.; et al. miR-7 Reduces Breast Cancer Stem Cell Metastasis via Inhibiting RELA to Decrease ESAM Expression. Mol. Ther. Oncolytics 2020, 18, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Wu, Y.-Y.; Xing, S.-J.; Yu, Z.-W. Effect of miR-7 on resistance of breast cancer cells to adriamycin via regulating EGFR/PI3K signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5285–5292. [Google Scholar] [PubMed]
- Hong, T.; Ding, J.; Li, W. miR-7 Reverses Breast Cancer Resistance To Chemotherapy By Targeting MRP1 And BCL2. OncoTargets Ther. 2019, 12, 11097–11105. [Google Scholar] [CrossRef] [Green Version]
- Paccez, J.D.; Duncan, K.; Sekar, D.; Correa, R.G.; Wang, Y.; Gu, X.; Bashin, M.; Chibale, K.; Libermann, T.A.; Zerbini, L.F. Dihydroartemisinin inhibits prostate cancer via JARID2/miR-7/miR-34a-dependent downregulation of Axl. Oncogenesis 2019, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.-L.; Zhou, P.-J.; Wei, L.; Li, W.; Ji, Z.; Fang, Y.-X.; Gao, W.-Q. MicroRNA-7 inhibits the stemness of prostate cancer stem-like cells and tumorigenesis by repressing KLF4/PI3K/Akt/p21 pathway. Oncotarget 2015, 6, 24017–24031. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, N.; Gallagher, M.; Lao, N.; Gallagher, C.; Clarke, C.; Doolan, P.; Aherne, S.; Blanco, A.; Meleady, P.; Clynes, M.; et al. MiR-7 Triggers Cell Cycle Arrest at the G1/S Transition by Targeting Multiple Genes Including Skp2 and Psme3. PLoS ONE 2013, 8, e65671. [Google Scholar] [CrossRef]
- Mallik, S.; Qin, G.; Jia, P.; Zhao, Z. Molecular signatures identified by integrating gene expression and methylation in non-seminoma and seminoma of testicular germ cell tumours. Epigenetics 2021, 16, 162–176. [Google Scholar] [CrossRef]
- Yu, Z.; Ni, L.; Chen, D.; Zhang, Q.; Su, Z.; Wang, Y.; Yu, W.; Wu, X.; Ye, J.; Yang, S.; et al. Identification of miR-7 as an oncogene in renal cell carcinoma. J. Mol. Histol. 2013, 44, 669–677. [Google Scholar] [CrossRef]
- Li, Q.; Wang, X.; Jiang, N.; Xie, X.; Liu, N.; Liu, J.; Shen, J.; Peng, T. Exosome-transmitted linc00852 associated with receptor tyrosine kinase AXL dysregulates the proliferation and invasion of osteosarcoma. Cancer Med. 2020, 9, 6354–6366. [Google Scholar] [CrossRef]
- Jiang, M.-J.; Dai, J.-J.; Gu, D.-N.; Huang, Q.; Tian, L. MicroRNA-7 inhibits cell proliferation of chronic myeloid leukemia and sensitizes it to imatinib in vitro. Biochem. Biophys. Res. Commun. 2017, 494, 372–378. [Google Scholar] [CrossRef]
- Zhang, H.; Luo, X.-Q.; Zhang, P.; Huang, L.-B.; Zheng, Y.-S.; Wu, J.; Zhou, H.; Qu, L.-H.; Xu, L.; Chen, Y.-Q. MicroRNA Patterns Associated with Clinical Prognostic Parameters and CNS Relapse Prediction in Pediatric Acute Leukemia. PLoS ONE 2009, 4, e7826. [Google Scholar] [CrossRef]
- O’Donnell, K.A.; Wentzel, E.A.; Zeller, K.I.; Dang, C.V.; Mendell, J.T. c-Myc-regulated microRNAs modulate E2F1 expression. Nat. Cell Biol. 2005, 435, 839–843. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Gao, L.; Zhao, J.; Liu, D.; Li, H.; Hu, M. LncRNA ANRIL/miR-7-5p/TCF4 axis contributes to the progression of T cell acute lymphoblastic leukemia. Cancer Cell Int. 2020, 20, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lou, X.; Fu, J.; Zhao, X.; Zhuansun, X.; Rong, C.; Sun, M.; Niu, H.; Wu, L.; Zhang, Y.; An, L.; et al. MiR-7e-5p downregulation promotes transformation of low-grade follicular lymphoma to aggressive lymphoma by modulating an immunosuppressive stroma through the upregulation of FasL in M1 macrophages. J. Exp. Clin. Cancer Res. 2020, 39, 1–15. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Biogenesis of miRNA. miRNA are transcribed from their genes regulated by promoters. Primary or pri-miRNA is several hundred base pairs long and has a form of a hairpin. It is processed by a microprocessor, a complex containing Drosha enzyme which removes the stem of the structure. Such pre-miRNA is exported from the nucleus and further cleaved by Dicer. miRNA duplex of 20–22 22 nt is produced. One strand of the duplex becomes the mature miRNA loaded on RNA-induced silencing complex (RISC), containing Argonaute protein. miRNA targets mRNA complementary to its sequence and directs it to degradation or inhibits translation, depending on the level of complementarity. Some of the known signaling molecules regulating miR-7 expression are shown [1,8,18,23,26,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,55].
Figure 1.
Biogenesis of miRNA. miRNA are transcribed from their genes regulated by promoters. Primary or pri-miRNA is several hundred base pairs long and has a form of a hairpin. It is processed by a microprocessor, a complex containing Drosha enzyme which removes the stem of the structure. Such pre-miRNA is exported from the nucleus and further cleaved by Dicer. miRNA duplex of 20–22 22 nt is produced. One strand of the duplex becomes the mature miRNA loaded on RNA-induced silencing complex (RISC), containing Argonaute protein. miRNA targets mRNA complementary to its sequence and directs it to degradation or inhibits translation, depending on the level of complementarity. Some of the known signaling molecules regulating miR-7 expression are shown [1,8,18,23,26,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,55].

Figure 2.
Effects of miR-7 on the process of carcinogenesis in different types of cancer. blue: tumor suppressor’s activities; red: activities as oncomirs.
Figure 2.
Effects of miR-7 on the process of carcinogenesis in different types of cancer. blue: tumor suppressor’s activities; red: activities as oncomirs.


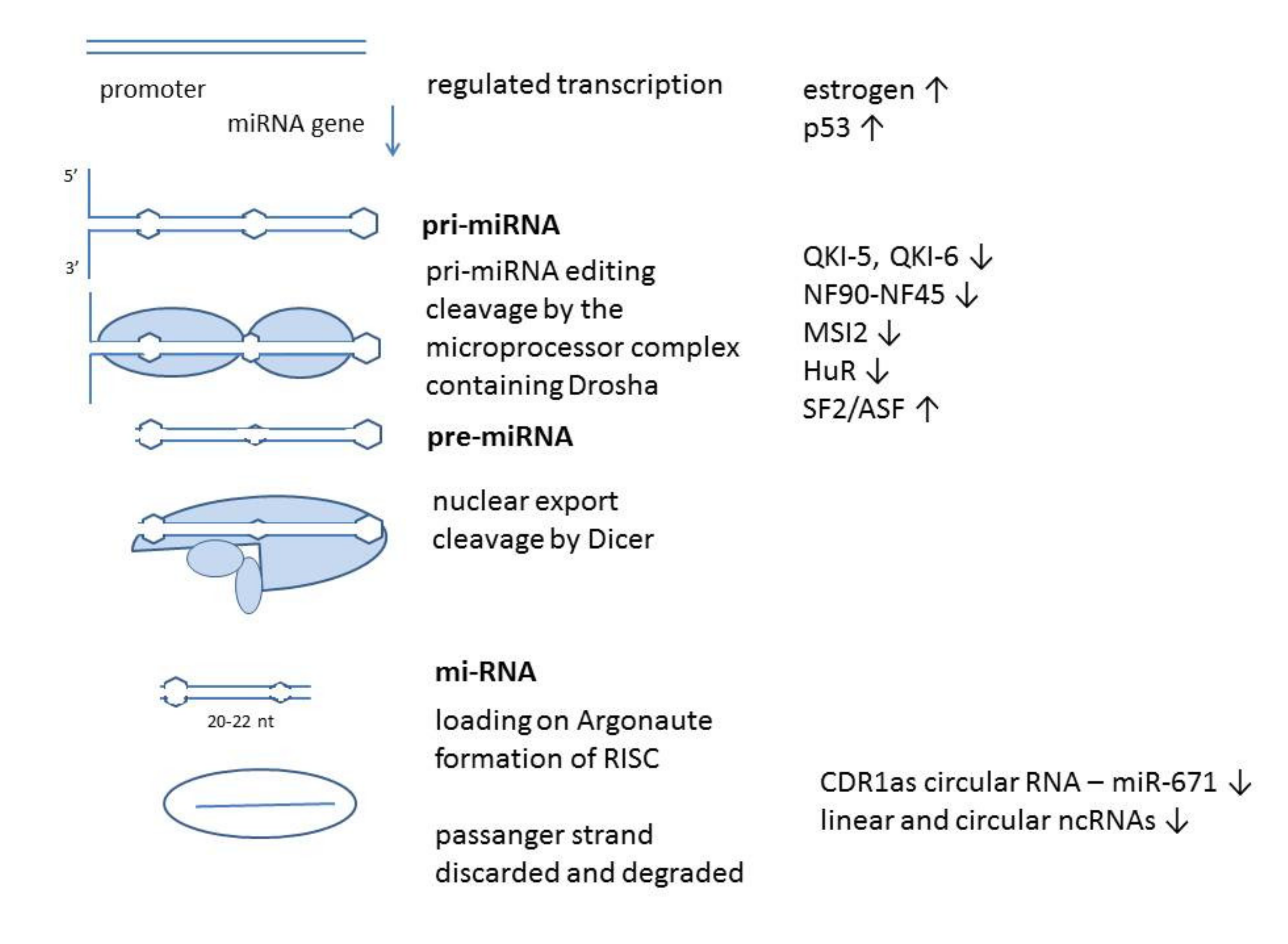{kind=link}
{kind=link}
Table 1.
miR-7 target genes/proteins in proliferation.
| Gene/Protein | Cell Type | Pathway | Function | Reference |
|---|---|---|---|---|
| EGFR | glioblastoma schwanoma lung cancer melanoma | EGFR signaling | inhibition of proliferation | [11,12,62,63,64] |
| PIK3R3 PIK3CD PI3K | glioblastoma lung cancer | PI3K/Akt pathway TLR9 pathway | proliferation inhibition | [65,66,67,68] |
| IRS-1, IRS-2 | glioblastoma tong squamous cell carcinoma melanoma | PI3K/Akt pathway | inhibition of proliferation viability | [62,69,70,71] |
| Raf1 | glioblastoma | EGFR signaling | inhibition of proliferation | [63,67,72,73] |
| FOS | gastric cancer | MAPK signaling | inhibition of proliferation | [74] |
| ARF4 | glioblastoma | MAPK signaling | inhibition of proliferation | [63] |
| mTOR p70S6K eIF4E Mapkap1 Mknk1 Mknk2 | glioblastoma pancreas | PI3K/Akt pathway | proliferation inhibition | [66,75] |
| MAP3K9 | pancreatic cancer | MAPK pathway | inhibition of proliferation and migration | [76] |
| IGF-1R | gastric metastasis glioma tong squamous carcinoma osteosarcoma | PI3K/Akt pathway IGF1-Snail EMT | inhibition of migration and survival | [71,77,78,79] |
| TAL1 | T acute lymphoblastic leukaemia | proliferation | inhibition of proliferation | [80] |
| RELA | gastric cancer melanoma | NFkappa B signaling | inhibition of proliferation | [74,81] |
| IKK eta | gastric cancer | NF kappa B | regulation of proliferation | [74] |
| Skp2 Psme | ovary cells | cell cycle regulation | cell cycle arrest | [82] |
| CCNE | liver hepatocellular carcinoma | cell cycle | inhibition of proliferation | [83] |
| KLF4 | epithelial brain metastasis endothelial cells lung | stem cell regulation | proliferation migration angiogenesis | [84,85,86] |
| RECK reversion inducing cysteine-rich protein with kazal motifs | oral cancer | metalloproteinase inhibitor | increase in proliferation | [87] |
| ERF Ets2 transcriptional repressor | lung cancer | MAPK pathway | increase in proliferation | [88] |
| CKS2 cyclin-dependent kinase regulatory subunit 2 | thyroid cancer | cell cycle | inhibition of proliferation | [89] |
| TRIP6 thyroid receptor interactor protein | colorectal cancer | proliferation | inhibition of proliferation and metastasis | [90] |
| ALDJ1A3 | breast cancer | stem cell survival | decrease in stem cell survival | [91] |
| CUL5 cullin5 | hepatocellular | ubiquitination and protein degradation | inhibition of proliferation cell cycle arrest | [92] |
| TYRO3 | colorectal cancer | PI3K/Akt/mTOR | inhibition of proliferation | [93] |
Table 2.
miR-7 target genes/proteins involved in migration.
| Gene/Protein | Cell Type | Pathway | Function | Reference |
|---|---|---|---|---|
| PAK2 PAK1 | lung carcinoma thyroid cancer gliomas tong squamous cell carcinomaschwanoma | Rho kinase effector | inhibition of proliferation, motility, regulation of cytoskeleton apoptosis | [11,63,67,69,71,94,95] |
| TBX2 T-Box2 | glioblastoma | differentiation, EMT | inhibition of invasiveness | [96] |
| trefoil factor 3 | glioblastoma | PI3K/Akt pathway | inhibition of migration | [97] |
| cdc42 | brain damaged | Rho kinase | inhibition of migration and proliferation | [98] |
| Ack1 associated cdc42 kinase 1 | schwannoma | Rho pathways citoskeleton regulation | inhibition of migration | [11] |
| FAK FAK (PTK2) | glioblastoma breast cancer colon cancer lung cancer | citoskeleton regulation | inhibition of migration and proliferation | [99,100,101,102] |
| NOVA2 | lung carcinoma | inhibition of migration | [103] | |
| LASP1 | breast cancer | [104] | ||
| SATB1 special AT rich sequence binding protein | glioblastoma | inhibition of migration and invasion | [105] | |
| Slug | breast cancer | EMT | decrease in migration | [30] |
| NFAT | pancreas | EMT | inhibition of migration | [106] |
| VE cadherin Notch4 | hepatocellular carcinoma | inhibition of migration | [107] | |
| KLF4 YY1 | Non-Hodgin lymphoma | inhibition of migration and chemosensitivity | [108] |
Table 3.
miR-7 target genes/proteins involved in apoptosis and protection from stress.
| Gene/Protein | Cell Type | Pathway | Function | Reference |
|---|---|---|---|---|
| BCL-2 | lung, liver | apoptosis | apoptosis | [109] |
| XIAP | glioblastoma cervical cancer hepatocellular carcinoma lung | apoptosis | apoptosis | [68,72,110] |
| SLC25A37 TIMM50 | rhabdomyosarcoma | mitochondria | induction of cell death ?? | [111] |
| REGγ proteasome activator subunit | breast cancer | proteasome | inhibition of proliferation increase of apoptosis | [112] |
| NEIL Nei endonuclease VIII-like 1 | colorectal cancer | inhibition of apoptosis, proliferation | inhibition of proliferation and survival | [113] |
| UBE2A | brain | ubiquitination and protein degradation | amyloid peptide proteolysis | [114] |
| YY1 | glioblastoma colon cancer | p53 pathway cell cycle arrest wnt signaling | resistance to alkylation | [13,115] |
| 1BRCA1 | breast cancer | DNA repair | decrease in survival | [104] |
| Sirtuin /Sirt1 | neuroblastoma | regulation of oxygen-glucose deprivation | protection from damage | [116] |
| VDAC | neuroblastoma hepatocellular carcinoma | ion channel on mitochondria; ROS defense | protection from oxidative stress | [117,118] |
| KEAP1 | neuroblastoma | ROS defense | protection from oxidative stress | [119] |
| HOXB3 | breast cancer retinal epithelial cells | glucose metabolism PI3K/Akt/mTOR | reduction of high glucose damage | [120] |
| PARP1 | lung cancer cells | DNA repair | decreased DNA repair and survival | [121] |
| REDD1 regulated in development and DNA damage response 1 | cervical carcinoma cells under hypoxia | DNA damage response | hypoxamir proliferation modulation | [122] |
| SMARCD1 | lung cancer cells | chromatin regulator p53 pathway | increased chemoresistance | [59] |
| XRCC2 | colorectal cancer cells | DNA repair | proliferation inhibition, induction of apoptosis | [123] |
| Rad54L | ovary cells | DNA damage repair | survival under cell cycle arrest conditions | [82] |
| REG1 regenerating islet-derived protein | pancreas | response to glucose starvation | inhibition of proliferation, apoptosis, differentiation | [124] |
| MRP1/ABCC1 | lung carcinoma | multidrug resistance | decreased survival | [125] |
| NF90 | tumor | DNA repair | DNA damage repair inhibition | [52] |
Table 4.
miR-7 target genes/proteins involved in differentiation and metabolic processes.
| Gene/Protein | CELL TYPE | Pathway | Function | Reference |
|---|---|---|---|---|
| TLR4 | brain | inflammation | downregulation of inflammation | [126] |
| FAM177A | macrophages | inflammation | inhibition of cytokine production | [127] |
| NLRP3 Nod like receptor | brain | inflammation | downregulation of inflammation | [128] |
| TET2 | hematopoietic malignancies | chromatin modification | [58] | |
| SETDB1 SETD8 | pancreas | chromatin regulation | [56,57] | |
| PAX6 | brain lung colon pancreas embryonic stem cells | differentiation | [5,129] | |
| Gli3 | brain bladder cancer | hedgehog | differentiation | [130,131] |
| FGFR4 | liver | protection from injury | stem cell proliferation | [132] |
| HoxD family | brain | differentiation | [133] | |
| TCF4 and TCF12 | brain | wnt pathway | differentiation | [134] |
| TCF7L2 | brain | wnt pathway | [134] | |
| SHANK3 | brain | differentiation | [135] | |
| ihog Hedgehog receptor | drosophila eye | hedgehog pathway | differentiation | [136] |
| CRY2 | osteoblast | CLOCK/BMAL/p300 pathway | differentiation | [137] |
| Yorkie | drosophila wings | Hippo pathway | organ size | [138] |
| G protein signalling 5 RGSS | eye | signaling | [139] | |
| PA28 gamma | lung carcinoma | proteasome | inhibition of protein degradation | [140] |
| insulin receptor INSR insulin receptor substrate 2 IRS-2 insulin-degrading enzyme IDE | brain | regulation of glucose metabolism | insulin sensitivity | [141] |
| TfR1 transferrin receptor 1 | pancreatic and colon cells | iron transport and storage | iron transport and storage | [142] |
| beta arrestin 1 | pancreatic beta cells | regulation of insulin secretion | metabolism | [143] |
| Sepp1b selenoprotein P | brain | synaptic function | [144] | |
| Prostaglandin F2 receptor negative regulator PTGFR Golgi glycoprotein 1 | pituitary gland | hormone regulation | gonad development | [145,146] |
| OGT, O-GlcNAcyl Transferase | lung cancer | O-GlcNAcylation | metabolic reprogramming migration | [147] |
| CAMK2D calponin | smooth muscle cell | calcification | vascular calcification in pulmonary hypertension | [148] |
| enolase ENO2 | nasopharyngeal carcinoma | glycolysis | metabolism radioresistance | [149] |
| Lactat dehydrogenase A | gastric cancer | glycolysis | metabolism | [150] |
| Raf1 | pituitary gland | production of prolactin | development | [151] |
| KLF4 | myoblasts | differentiation and proliferation | inhibition of differentiation and proliferation | [152] |
| Follicle stimulating hormone FSH | pituitary gland | metabolism | inhibition of production | [48] |
| alpha-Synuclein | brain; Parkinson disease | neuron function and survival | inhibition of production | [153] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Korać, P.; Antica, M.; Matulić, M. MiR-7 in Cancer Development. Biomedicines 2021, 9, 325. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9030325
AMA Style
Korać P, Antica M, Matulić M. MiR-7 in Cancer Development. Biomedicines. 2021; 9(3):325. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9030325
Chicago/Turabian StyleKorać, Petra, Mariastefania Antica, and Maja Matulić. 2021. "MiR-7 in Cancer Development" Biomedicines 9, no. 3: 325. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9030325
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.