
Artificially Induced Pluripotent Stem Cell-Derived Whole-Brain Organoid for Modelling the Pathophysiology of Metachromatic Leukodystrophy and Drug Repurposing


Abstract
:1. Introduction
Current and Future Potential for MLD Modeling
2. Methods
2.1. The Whole-Brain Organoid Simulations, Using NEUBOrg (DeepNEU v6.2)
2.2. The NEUBOrg, aiWBO Simulations Applied to MLD
2.3. NEUBOrg Platform Statistical Analysis
2.4. Drug Target Discovery and Drug Repurposing for Treating MLD
3. Results
3.1. The NEUBOrg Platform Specification
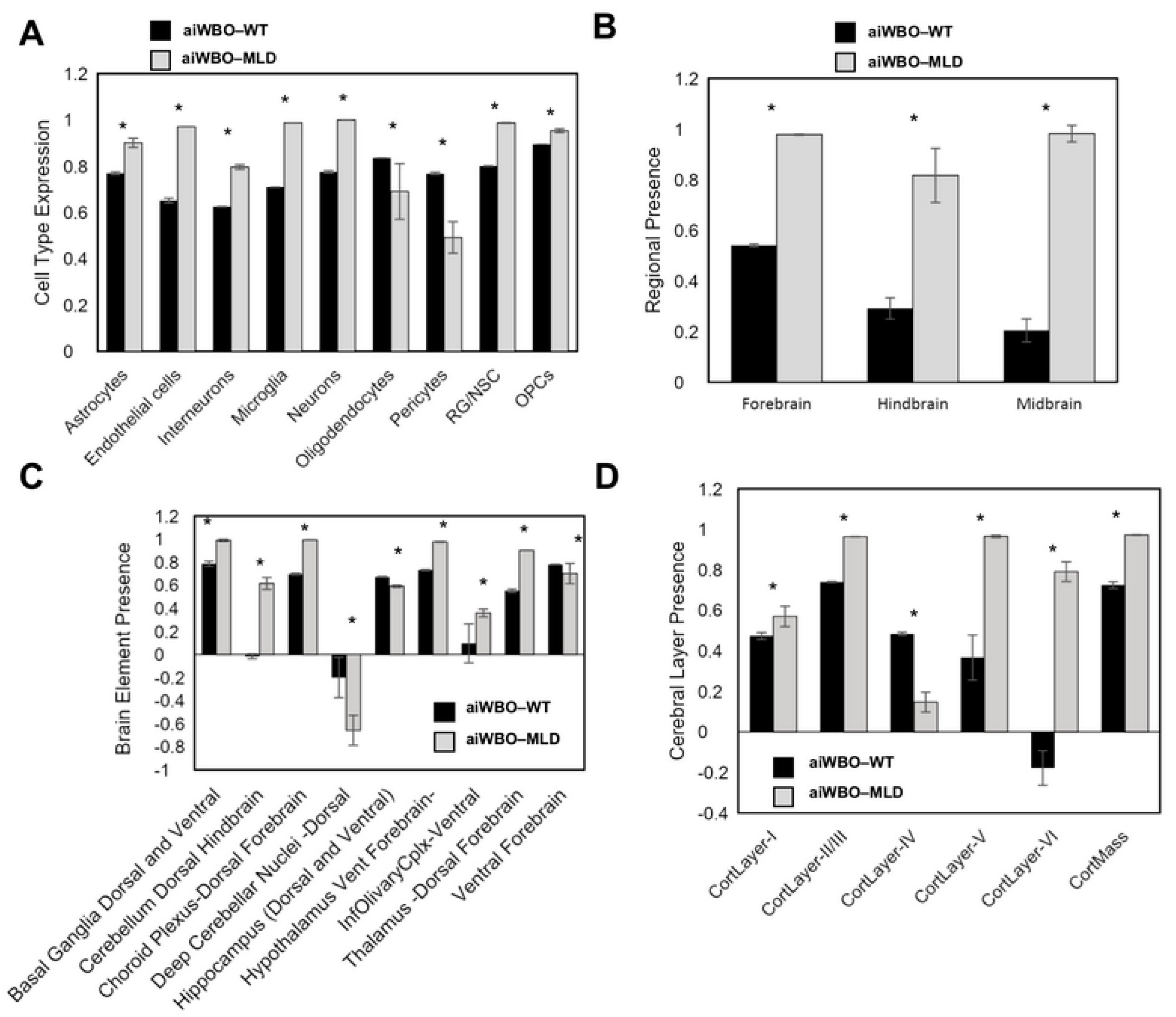
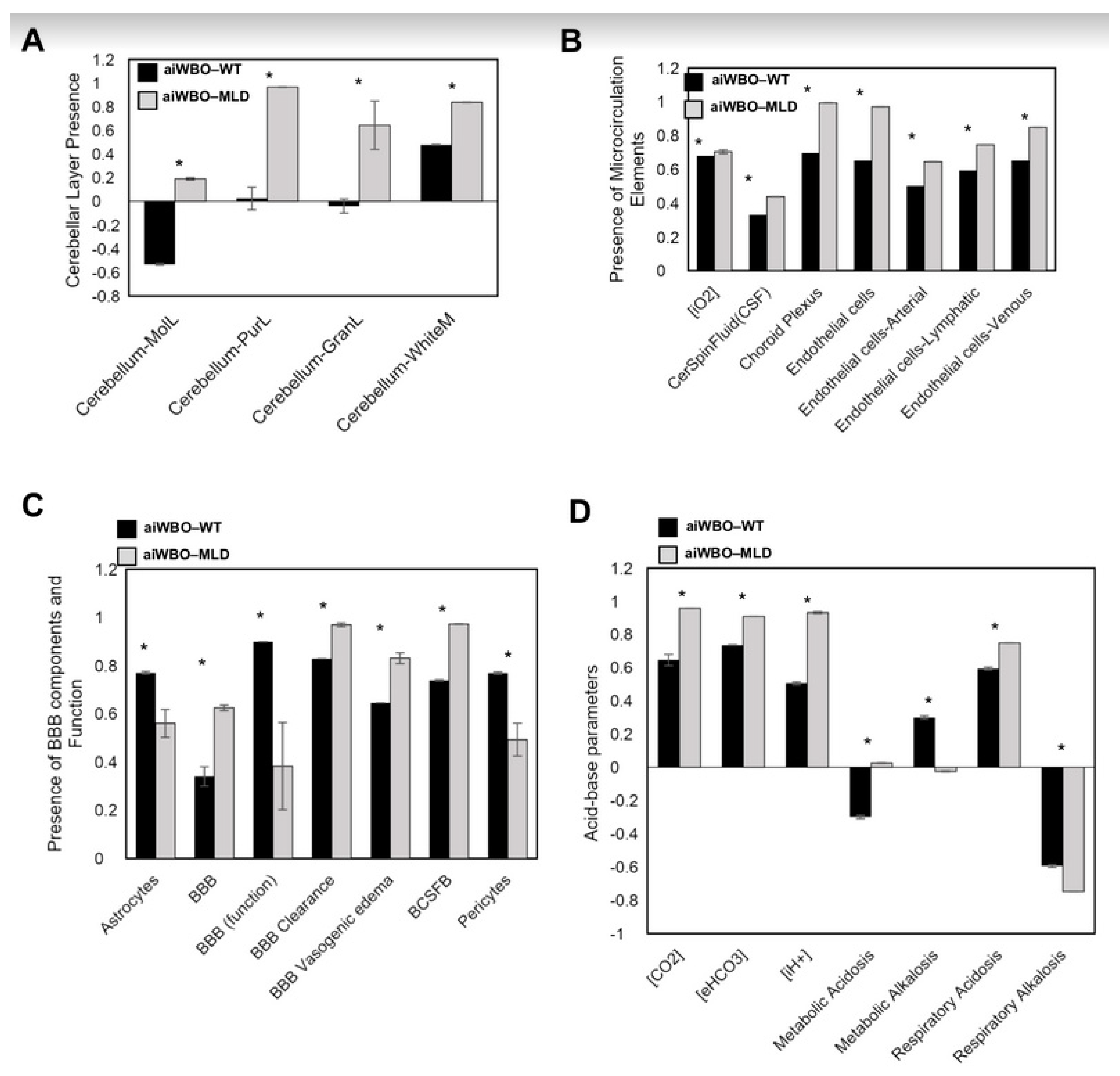
3.2. The aiWBO Wild Type Simulations
4. Summary of aiWBO–WT Simulations Profile
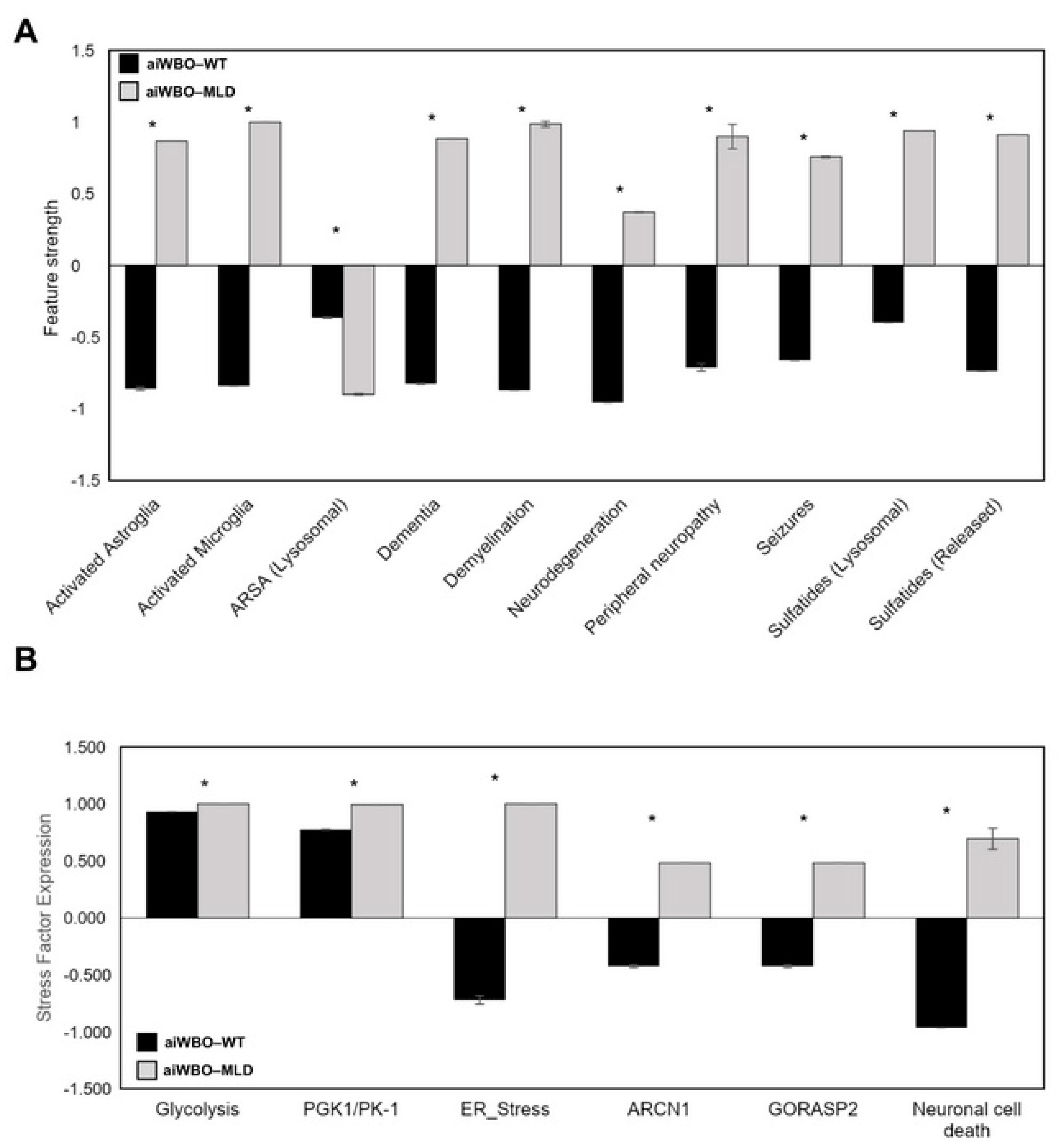
5. Summary of aiWBO–MLD Disease Profile Simulations
Results of Drug Target Evaluation and Drug Repurposing Efforts
6. Discussion
6.1. The Issues of Cellular Stress and Neuron Cell Death
6.2. Drug Repurposing for MLD
6.3. Promises and Pitfalls of the Current aiWBO Simulations
7. Conclusions/Future Directions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Liaw, H.-R.; Lee, H.-F.; Chi, C.-S.; Tsai, C.-R. Late infantile metachromatic leukodystrophy: Clinical manifestations of five Taiwanese patients and Genetic features in Asia. Orphanet J. Rare Dis. 2015, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Maegawa, G.H. Lysosomal leukodystrophies lysosomal storage diseases associated with white matter abnormalities. J. Child Neurol. 2019, 34, 339–358. [Google Scholar] [CrossRef]
- Eichler, F.S. Metachromatic leukodystrophy and multiple sulfatase deficiency. In Rosenberg’s Molecular and Genetic Basis of Neurological and Psychiatric Disease; Elsevier: Amsterdam, The Netherlands, 2020; pp. 473–479. [Google Scholar]
- Meneghini, V.; Frati, G.; Sala, D.; De Cicco, S.; Luciani, M.; Cavazzin, C.; Paulis, M.; Mentzen, W.; Morena, F.; Giannelli, S. Generation of Human Induced Pluripotent Stem Cell-Derived Bona Fide Neural Stem Cells for Ex Vivo Gene Therapy of Metachromatic Leukodystrophy. Stem Cells Transl. Med. 2017, 6, 352–368. [Google Scholar] [CrossRef]
- Frati, G.; Luciani, M.; Meneghini, V.; De Cicco, S.; Ståhlman, M.; Blomqvist, M.; Grossi, S.; Filocamo, M.; Morena, F.; Menegon, A. Human iPSC-based models highlight defective glial and neuronal differentiation from neural progenitor cells in metachromatic leukodystrophy. Cell Death Dis. 2018, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Danter, W.R. DeepNEU: Cellular reprogramming comes of age—A machine learning platform with application to rare diseases research. Orphanet J. Rare Dis. 2019, 14, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Esmail, S.; Danter, W.R. NEUBOrg: Artificially Induced Pluripotent Stem Cell-Derived Brain Organoid to Model and Study Genetics of Alzheimer’s Disease Progression. Front. Aging Neurosci. 2021, 13, 643889. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hattori, N. Cerebral organoids model human brain development and microcephaly. Mov. Disord. 2014, 2, 185. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A.; Knoblich, J.A. Generation of cerebral organoids from human pluripotent stem cells. Nat. Protoc. 2014, 9, 2329–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lancaster, M.A.; Huch, M. Disease modelling in human organoids. Dis. Models Mech. 2019, 12, dmm039347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Koo, B.-K.; Knoblich, J.A. Human organoids: Model systems for human biology and medicine. Nat. Rev. Mol. Cell Biol. 2020, 21, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Logan, S.; Arzua, T.; Canfield, S.G.; Seminary, E.R.; Sison, S.L.; Ebert, A.D.; Bai, X. Studying human neurological disorders using induced pluripotent stem cells: From 2D monolayer to 3D organoid and blood brain barrier models. Compr. Physiol. 2011, 9, 565–611. [Google Scholar]
- Chen, H.I.; Song, H.; Ming, G.l. Applications of human brain organoids to clinical problems. Dev. Dyn. 2019, 248, 53–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, X.; Song, H.; Ming, G.-l. Brain organoids: Advances, applications and challenges. Development 2019, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Nelson, A.R.; Betsholtz, C.; Zlokovic, B.V. Establishment and dysfunction of the blood-brain barrier. Cell 2015, 163, 1064–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H. Modeling neurological diseases with human brain organoids. Front. Synaptic Neurosci. 2018, 10, 15. [Google Scholar] [CrossRef]
- Chen, A.; Guo, Z.; Fang, L.; Bian, S. Application of Fused Organoid Models to Study Human Brain Development and Neural Disorders. Front. Cell. Neurosci. 2020, 14, 133. [Google Scholar] [CrossRef]
- Qian, X.; Jacob, F.; Song, M.M.; Nguyen, H.N.; Song, H.; Ming, G.-L. Generation of human brain region–specific organoids using a miniaturized spinning bioreactor. Nat. Protoc. 2018, 13, 565. [Google Scholar] [CrossRef]
- Bhaduri, A.; Andrews, M.G.; Leon, W.M.; Jung, D.; Shin, D.; Allen, D.; Jung, D.; Schmunk, G.; Haeussler, M.; Salma, J. Cell stress in cortical organoids impairs molecular subtype specification. Nature 2020, 578, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.R.; Cardoso, I.; Gonçalves, I. Key enzymes and proteins in amyloid-β production and clearance. In Alzheimer’s Disease Pathogenesis—Core Concepts, Shifting Paradigms and Therapeutic Targets; de la Monte, S., Ed.; IntechOpen: London, UK, 2011; pp. 53–86. [Google Scholar]
- Lippmann, E.S.; Williams, C.E.; Ruhl, D.A.; Estevez-Silva, M.C.; Chapman, E.R.; Coon, J.J.; Ashton, R.S. Deterministic HOX patterning in human pluripotent stem cell-derived neuroectoderm. Stem Cell Rep. 2015, 4, 632–644. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.-I.; Lee, K.-H.; Shin, D.-M.; Shim, J.-W.; Lee, J.-S.; Chang, S.Y.; Lee, Y.-S.; Brownstein, M.J.; Lee, S.-H.; Lee, Y.-S. Comprehensive transcriptome analysis of differentiation of embryonic stem cells into midbrain and hindbrain neurons. Dev. Biol. 2004, 265, 491–501. [Google Scholar] [CrossRef] [Green Version]
- Kirsch, L.; Liscovitch, N.; Chechik, G. Localizing genes to cerebellar layers by classifying ISH images. PLoS Comput. Biol. 2012, 8, e1002790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logan, S.; Arzua, T.; Yan, Y.; Jiang, C.; Liu, X.; Yu, L.-K.; Liu, Q.-S.; Bai, X. Dynamic Characterization of Structural, Molecular, and Electrophysiological Phenotypes of Human-Induced Pluripotent Stem Cell-Derived Cerebral Organoids, and Comparison with Fetal and Adult Gene Profiles. Cells 2020, 9, 1301. [Google Scholar] [CrossRef] [PubMed]
- Papaspyropoulos, A.; Tsolaki, M.; Foroglou, N.; Pantazaki, A.A. Modeling and Targeting Alzheimer’s Disease With Organoids. Front. Pharmacol. 2020, 11, 396. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Xu, R.; Padmashri, R.; Dunaevsky, A.; Liu, Y.; Dreyfus, C.F.; Jiang, P. Pluripotent stem cell-derived cerebral organoids reveal human oligodendrogenesis with dorsal and ventral origins. Stem Cell Rep. 2019, 12, 890–905. [Google Scholar] [CrossRef] [Green Version]
- Gerakis, Y.; Hetz, C. Brain organoids: A next step for humanized Alzheimer’s disease models? Mol. Psychiatry 2019, 24, 474–478. [Google Scholar] [CrossRef] [Green Version]
- Espuny-Camacho, I.; Michelsen, K.A.; Linaro, D.; Bilheu, A.; Acosta-Verdugo, S.; Herpoel, A.; Giugliano, M.; Gaillard, A.; Vanderhaeghen, P. Human pluripotent stem-cell-derived cortical neurons integrate functionally into the lesioned adult murine visual cortex in an area-specific way. Cell Rep. 2018, 23, 2732–2743. [Google Scholar] [CrossRef] [Green Version]
- Zemke, M.; Draganova, K.; Klug, A.; Schöler, A.; Zurkirchen, L.; Gay, M.H.-P.; Cheng, P.; Koseki, H.; Valenta, T.; Schübeler, D. Loss of Ezh2 promotes a midbrain-to-forebrain identity switch by direct gene derepression and Wnt-dependent regulation. BMC Biol. 2015, 13, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Krefft, O.; Jabali, A.; Iefremova, V.; Koch, P.; Ladewig, J. Generation of standardized and reproducible forebrain-type cerebral organoids from human induced pluripotent stem cells. JoVE 2018, 131, e56768. [Google Scholar] [CrossRef] [Green Version]
- Do, J.T.; Hong, Y.J. Neural lineage differentiation from pluripotent stem cells to mimic human brain tissues. Front. Bioeng. Biotechnol. 2019, 7, 400. [Google Scholar]
- Tkachenko, L.A.; Zykin, P.A.; Nasyrov, R.A.; Krasnoshchekova, E.I. Distinctive features of the human marginal zone and Cajal–Retzius cells: Comparison of morphological and immunocytochemical features at midgestation. Front. Neuroanat. 2016, 10, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velasco, S.; Kedaigle, A.J.; Simmons, S.K.; Nash, A.; Rocha, M.; Quadrato, G.; Paulsen, B.; Nguyen, L.; Adiconis, X.; Regev, A. Individual brain organoids reproducibly form cell diversity of the human cerebral cortex. Nature 2019, 570, 523–527. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, A.T.; Wang, M.; Hauberg, M.E.; Fullard, J.F.; Kozlenkov, A.; Keenan, A.; Hurd, Y.L.; Dracheva, S.; Casaccia, P.; Roussos, P. Brain cell type specific gene expression and co-expression network architectures. Sci. Rep. 2018, 8, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichler, F.; Grodd, W.; Grant, E.; Sessa, M.; Biffi, A.; Bley, A.; Kohlschuetter, A.; Loes, D.; Kraegeloh-Mann, I. Metachromatic leukodystrophy: A scoring system for brain MR imaging observations. Am. J. Neuroradiol. 2009, 30, 1893–1897. [Google Scholar] [CrossRef] [Green Version]
- Harrington, M.; Whalley, D.; Twiss, J.; Rushton, R.; Martin, S.; Huynh, L.; Yang, H. Insights into the natural history of metachromatic leukodystrophy from interviews with caregivers. Orphanet J. Rare Dis. 2019, 14, 1–10. [Google Scholar] [CrossRef]
- Rosenberg, J.B.; Kaminsky, S.M.; Aubourg, P.; Crystal, R.G.; Sondhi, D. Gene therapy for metachromatic leukodystrophy. J. Neurosci. Res. 2016, 94, 1169–1179. [Google Scholar] [CrossRef] [Green Version]
- Hettiarachchi, D.; Dissanayake, V. Three novel variants in the arylsulfatase A (ARSA) gene in patients with metachromatic leukodystrophy (MLD). BMC Res. Notes 2019, 12, 1–4. [Google Scholar] [CrossRef]
- Pekgül, F.; Eroğlu-Ertuğrul, N.G.; Bekircan-Kurt, C.E.; Erdem-Ozdamar, S.; Çetinkaya, A.; Tan, E.; Konuşkan, B.; Karaağaoğlu, E.; Topçu, M.; Akarsu, N.A. Comprehensive clinical, biochemical, radiological and genetic analysis of 28 Turkish cases with suspected metachromatic leukodystrophy and their relatives. Mol. Genet. Metab. Rep. 2020, 25, 100688. [Google Scholar] [CrossRef]
- Narayanan, D.L.; Matta, D.; Gupta, N.; Kabra, M.; Ranganath, P.; Aggarwal, S.; Phadke, S.R.; Datar, C.; Gowrishankar, K.; Kamate, M. Spectrum of ARSA variations in Asian Indian patients with Arylsulfatase A deficient metachromatic leukodystrophy. J. Hum. Genet. 2019, 64, 323–331. [Google Scholar] [CrossRef]
- Mahdieh, N.; Soveizi, M.; Tavasoli, A.R.; Rabbani, A.; Ashrafi, M.R.; Kohlschütter, A.; Rabbani, B. Genetic testing of leukodystrophies unraveling extensive heterogeneity in a large cohort and report of five common diseases and 38 novel variants. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Addinsoft. XLSTAT Statistical and Data Analysis Solution; Addinsoft Long Island: New York, NY, USA, 2019. [Google Scholar]
- Beerepoot, S.; Nierkens, S.; Boelens, J.J.; Lindemans, C.; Bugiani, M.; Wolf, N.I. Peripheral neuropathy in metachromatic leukodystrophy: Current status and future perspective. Orphanet J. Rare Dis. 2019, 14, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groumpos, P.P. Deep learning vs. wise learning: A critical and challenging overview. IFAC Pap. OnLine 2016, 49, 180. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Simulation Summary | Components (Simulated) |
|---|---|
| Yamanaka (2007) transcription factors | OCT4, cMYC, KLF4 and SOX2 turned ON |
| B27 neural media | biotin, amino acids, ascorbate, catalase, cortisol, basic fibroblast growth factor (FGF2/bFGF), glutathione, albumin, insulin, SOD1(Cu/Zn), MnSOD/SOD2, progesterone, retinol/vitamin A, thyroid hormones (T3/T4), transferrin, VitE/Tocopherol, l-carnitine locked ON |
| Supplements | zinc and doxycycline locked ON |
| Rotating bioreactor (optimized) | B27 media + [CO2] = 5%, [O2] = 21%, glucose, temperature = 37 degrees C locked ON and high shear forces locked OFF |
| Age—evolved by the algorithm | Fetal/neonatal |
| MLD Features (N = 10) | Genotypic/Phenotypic Feature Inputs |
|---|---|
| ARSA gene ** | N = 4 (0 negatives and 4 positives) |
| Astrocyte/astroglial cell activation | N = 23 (5 negatives and 18 positives) |
| Microglial cell activation | N = 48 (17 negatives and 31 positives) |
| Dementia | N = 18 (5 negatives and 13 positives) |
| Demyelination | N = 22 (8 negatives and 14 positives) |
| Neurodegeneration | N = 39 (8 negatives and 31 positives) |
| Sulfatides (lysosomal) | N = 4 (1 negative and 3 positives) |
| Sulfatides (released) | N = 3 (0 negatives and 3 positives) |
| Seizures | N = 14 (5 negatives and 9 positives) |
| Peripheral neuropathy | N = 5 (0 negative and 5 positives) |
| Therapeutic Option | Activated Astroglia | Activated Microglia | Dementia | Demyelination | Neurodegeneration | Peripheral Neuropathy | Seizures | Sulfatides (Lysosomal) | Sulfatides (Released) | CS | ACD | t–Test p Value |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Placebo | 0.9 | 0.8 | 0.7 | 0.9 | 0.4 | 0.9 | 0.7 | 0.9 | 0.9 | 1.0 | 0.0 | 1.0 |
| Regorafenib + Olaparib | −0.9 | −0.9 | −0.6 | −1.0 | −0.4 | −0.8 | −0.7 | −0.8 | −0.9 | −1.0 | 1.0 | <0.00001 |
| Pembrolizumab + Lenvatinib | −0.9 | −0.9 | −0.7 | −0.9 | −0.3 | −0.8 | −0.7 | −0.8 | −0.9 | −1.0 | 1.0 | <0.00001 |
| Sunitinib+Lenvatinib | −0.8 | −0.8 | −0.6 | −0.9 | −0.3 | −0.8 | −0.7 | −0.9 | −0.9 | −1.0 | 1.0 | <0.00001 |
| Lenvatinib + Capmatinib | −0.8 | −0.9 | −0.7 | −1.0 | −0.3 | −0.8 | −0.7 | −0.9 | −0.9 | −1.0 | 1.0 | <0.00001 |
| Rapamycin + Lenvatinib | −0.8 | −0.9 | −0.6 | −1.0 | −0.3 | −0.8 | −0.7 | −0.9 | −0.9 | −1.0 | 1.0 | <0.00001 |
| Regorafenib + Lenvatinib | −0.9 | −0.9 | −0.7 | −1.0 | −0.3 | −0.8 | −0.7 | −0.9 | −0.9 | −1.0 | 1.0 | <0.00001 |
| Regorafenib + Calpain Inhibitor | −0.9 | −0.9 | −0.6 | −1.0 | −0.3 | −0.8 | −0.7 | −0.8 | −0.9 | −1.0 | 1.0 | <0.00001 |
| Sunitinib + Nutlin3 | −0.8 | −0.8 | −0.6 | −0.9 | −0.3 | −0.8 | −0.6 | −0.8 | −0.8 | −1.0 | 1.0 | <0.00001 |
| Sunitinib + Olaparib | −0.8 | −0.9 | −0.7 | −1.0 | −0.4 | −0.8 | −0.7 | −0.9 | −0.9 | −1.0 | 1.0 | <0.00001 |
| Olaparib + Abemaciclib | −0.8 | −0.9 | −0.6 | −1.0 | −0.4 | −0.8 | −0.6 | −0.8 | −0.8 | −1.0 | 1.0 | <0.00001 |
| Palbociclib + Olaparib | −0.8 | −0.9 | −0.6 | −1.0 | −0.4 | −0.8 | −0.6 | −0.8 | −0.8 | −1.0 | 1.0 | <0.00001 |
| Ribociclib + Olaparib | −0.8 | −0.9 | −0.6 | −1.0 | −0.4 | −0.8 | −0.6 | −0.8 | −0.8 | −1.0 | 1.0 | <0.00001 |
| Drug Class | Class Examples |
|---|---|
| Ca++ Homeostasis is Agents, N = 3 | Calcium Channel Blocker (VDCC), Calpain Inhibitor, Calpastatin Agonist |
| Check point Inhibitor, N = 2 | Ipilimumab, Pembrolizumab |
| Chemotherapy agents, N = 5 | Cisplatin, Dichloroacetate, Doxorubicin, Gemcitabine, Taxol |
| Multitargeted agents, N = 14 | Afatinib, Alectinib, Brigatinib, Cabozantinib, Capmatinib, Crizotinib, Imatinib |
| Targeted agents, N = 16 | Bortezomib, Nutlin3, Olaparib, Rapamycin, Tamoxifen, Vitamin D3, Anakinra. Jak2 inhibitor, Rituximab, Tocilizumab, Abemaciclib, Bevacizumab, Cetuximab, Palbociclib, Enhertu, Ribociclib |
| Other, N = 1 | Placebo |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esmail, S.; Danter, W.R. Artificially Induced Pluripotent Stem Cell-Derived Whole-Brain Organoid for Modelling the Pathophysiology of Metachromatic Leukodystrophy and Drug Repurposing. Biomedicines 2021, 9, 440. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9040440
Esmail S, Danter WR. Artificially Induced Pluripotent Stem Cell-Derived Whole-Brain Organoid for Modelling the Pathophysiology of Metachromatic Leukodystrophy and Drug Repurposing. Biomedicines. 2021; 9(4):440. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9040440
Chicago/Turabian StyleEsmail, Sally, and Wayne R. Danter. 2021. "Artificially Induced Pluripotent Stem Cell-Derived Whole-Brain Organoid for Modelling the Pathophysiology of Metachromatic Leukodystrophy and Drug Repurposing" Biomedicines 9, no. 4: 440. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9040440