
Increasing Odontoblast-like Differentiation from Dental Pulp Stem Cells through Increase of β-Catenin/p-GSK-3β Expression by Low-Frequency Electromagnetic Field


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation and Culture of hDPSCs
2.2. Odontoblast-Like Differentiation of hDPSCs
2.3. PEMF
2.4. Cell Viability
2.5. Cytotoxicity Assay
2.6. Cell Surface Antigen Analysis by FACS
2.7. Western Blotting
2.8. RT-PCR and Real-Time PCR Analysis
2.9. Immunofluorescence
2.10. Von Kossa Staining
2.11. Statistical Analysis
3. Results
3.1. Non-Cytotoxicity of PEMF Exposure
3.2. hDPSCs Were Differentiated by Media and PEMF
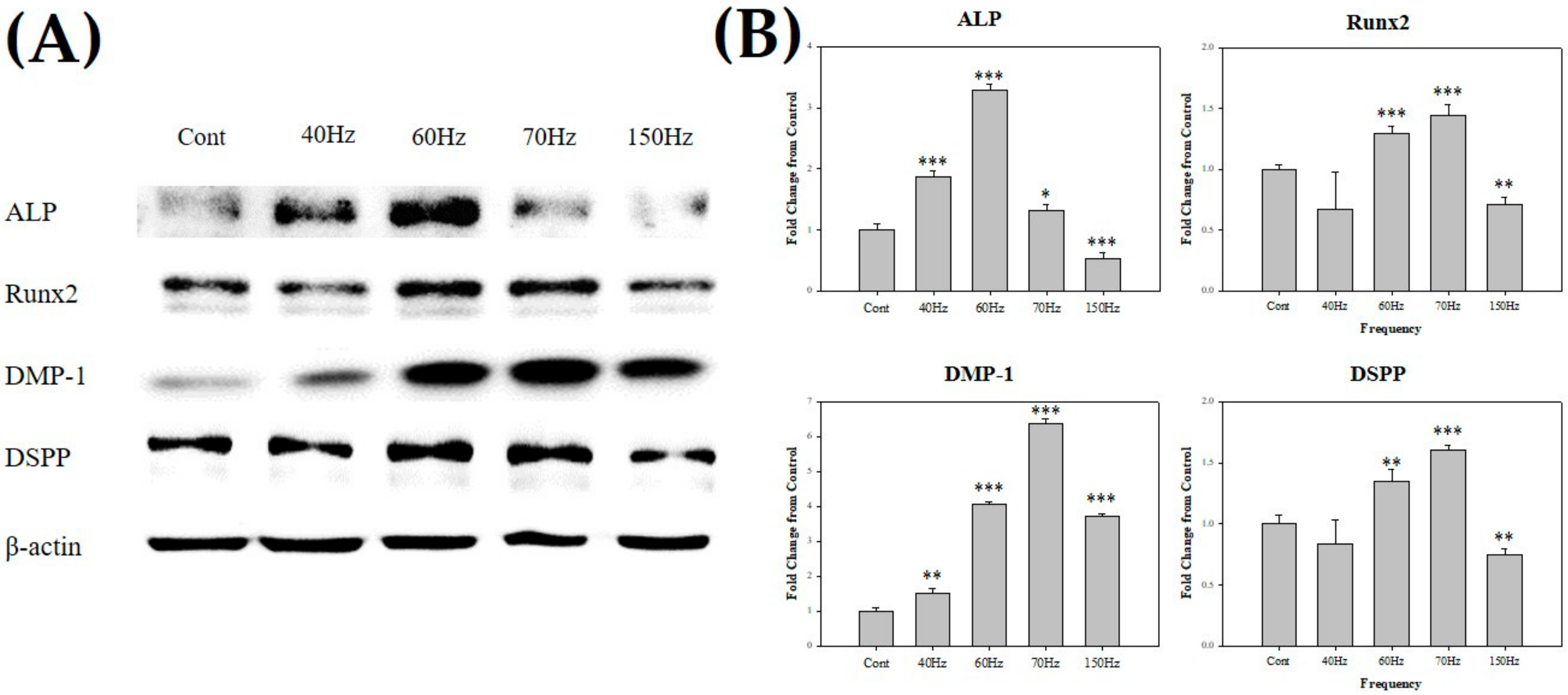
3.3. PEMF Exposure-Induced High Expression of Odontoblast-Related Molecules
3.4. DMP-1 and β-Catenin Increased in Nucleus and Cytoplasm of Cells Exposed to PEMF of Certain Frequency
3.5. PEMF Exposure at Certain Frequencies Promotes Odontoblast-Related Proteins through Phosphorylation of Glycogen Synthase Kinase-3 Beta (GSK-3β)/β-Catenin
3.6. Increased Calcium Deposition by PEMF Exposure
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Grawish, M.E.; Grawish, L.M.; Grawish, H.M.; Grawish, M.M.; El-Negoly, S.A. Challenges of Engineering Biomimetic Dental and Paradental Tissues. Tissue Eng. Regen. Med. 2020, 17, 403–421. [Google Scholar] [CrossRef]
- Han, M.-J.; Seo, Y.-K.; Yoon, H.-H.; Song, K.-Y.; Park, J.-K. Upregulation of bone-like extracellular matrix expression in human dental pulp stem cells by mechanical strain. Biotechnol. Bioprocess Eng. 2010, 15, 572–579. [Google Scholar] [CrossRef]
- Li, Y.; Nan, X.; Zhong, T.-Y.; Li, T.; Li, A.J.T.E.; Medicine, R. Treatment of Periodontal Bone Defects with Stem Cells from Inflammatory Dental Pulp Tissues in Miniature Swine. Tissue Eng. Regen. Med. 2019, 16, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.-L.; Chung, J.-H.; Choung, P.-H. Allogeneic Fibrin Clot for odontogenic/Cementogenic Differentiation of Human Dental Mesenchymal Stem Cells. Tissue Eng. Regen. Med. 2020, 17, 511–524. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Chen, L.; Xue, T.; Yu, J.; Liu, J.; Ji, Y.; Cheng, L. Human umbilical cord and dental pulp-derived mesenchymal stem cells: Biological characteristics and potential roles in vitro and in vivo. Mol. Med. Rep. 2015, 11, 3269–3278. [Google Scholar] [CrossRef]
- Hokmabad, V.R.; Davaran, S.; Aghazadeh, M.; Alizadeh, E.; Salehi, R.; Ramazani, A.J. A comparison of the effects of silica and hydroxyapatite nanoparticles on poly (ε-caprolactone)-poly (ethylene glycol)-poly (ε-caprolactone)/chitosan Nanofibrous scaffolds for bone tissue engineering. Tissue Eng. Regen. Med. 2018, 15, 735–750. [Google Scholar] [CrossRef] [PubMed]
- Tjäderhane, L.; Haapasalo, M.J.E.T. The dentin–pulp border: A dynamic interface between hard and soft tissues. Endod. Top. 2009, 20, 52–84. [Google Scholar] [CrossRef]
- Baldión, P.A.; Velandia-Romero, M.L.; Castellanos, J.E. Odontoblast-like cells differentiated from dental pulp stem cells retain their phenotype after subcultivation. Int. J. Cell Biol. 2018, 2018, 6853189. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, K.; Gajjeraman, S.; Ramachandran, A.; Hao, J.; George, A. Dentin matrix protein 1 regulates dentin sialophosphoprotein gene transcription during early odontoblast differentiation. J. Biol. Chem. 2006, 281, 19064–19071. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.G.; Zheng, Y.; Zhou, J.; Chen, M.; Embree, M.C.; Song, K.; Jiang, N.; Mao, J.J. Dentin and dental pulp regeneration by the patient’s endogenous cells. Endod. Top. 2013, 28, 106–117. [Google Scholar] [CrossRef]
- Angelova, A.V.; Zaugg, L.K.; Neves, V.; Liu, Y.; Sharpe, P.T. Tooth Repair and Regeneration. Curr. Oral Health Rep. 2018, 5, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S.; Haruyama, N.; Nishimura, F.; Kulkarni, A.B. Dentin sialophosphoprotein and dentin matrix protein-1: Two highly phosphorylated proteins in mineralized tissues. Arch. Oral Biol. 2012, 57, 1165–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Li, W.; Wang, H.; Wan, C.; Luo, D.; Deng, S.; Chen, H.; Chen, S. Klf10 regulates odontoblast differentiation and mineralization via promoting expression of dentin matrix protein 1 and dentin sialophosphoprotein genes. Cell Tissue Res. 2016, 363, 385–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, S.; Wang, J.; Dong, F.; Du, N.; Li, W.; Song, P.; Liu, Y. Concentrated Growth Factor Promotes Dental Pulp Cells Proliferation and Mineralization and Facilitates Recovery of Dental Pulp Tissue. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 10016. [Google Scholar] [CrossRef]
- Caiaffa, K.S.; Basso, F.G.; Santos-Filho, N.A.; de Souza-Costa, C.A.; Sakai, V.T.; Cilli, E.M.; Duque, C. Effect of analogues of cationic peptides on dentin mineralization markers in odontoblast-like cells. Arch. Oral Biol. 2019, 103, 19–25. [Google Scholar] [CrossRef]
- Murakami, M.; Hayashi, Y.; Iohara, K.; Osako, Y.; Hirose, Y.; Nakashima, M. Trophic effects and regenerative potential of mobilized mesenchymal stem cells from bone marrow and adipose tissue as alternative cell sources for pulp/dentin regeneration. Cell Transplant. 2015, 24, 1753–1765. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.B.; Tian, W.D.; Liu, L.K.; Xu, Y. In vitro odontoblast-like cell differentiation of cranial neural crest cells induced by fibroblast growth factor 8 and dentin non-collagen proteins. Cell Biol. Int. 2008, 32, 671–678. [Google Scholar] [CrossRef]
- Miyashita, S.; Ahmed, N.E.M.B.; Murakami, M.; Iohara, K.; Yamamoto, T.; Horibe, H.; Kurita, K.; Takano-Yamamoto, T.; Nakashima, M. Mechanical forces induce odontoblastic differentiation of mesenchymal stem cells on three-dimensional biomimetic scaffolds. J. Tissue Eng. Regen. Med. 2017, 11, 434–446. [Google Scholar] [CrossRef]
- Li, Z.-Y.; Chen, L.; Liu, L.; Lin, Y.-F.; Li, S.-W.; Tian, W.-D. Odontogenic potential of bone marrow mesenchymal stem cells. J. Oral Maxillofac. Surg. 2007, 65, 494–500. [Google Scholar] [CrossRef]
- Liao, C.; Ou, Y.; Wu, Y.; Zhou, Y.; Liang, S.; Wang, Y. Sclerostin inhibits odontogenic differentiation of human pulp-derived odontoblast-like cells under mechanical stress. J. Cell. Physiol. 2019, 234, 20779–20789. [Google Scholar] [CrossRef]
- Kim, Y.-M.; Cho, S.-E.; Kim, S.-C.; Jang, H.-J.; Seo, Y.-K. Effects of extremely low frequency electromagnetic fields on melanogenesis through p-ERK and p-SAPK/JNK pathways in human melanocytes. Int. J. Mol. Sci. 2017, 18, 2120. [Google Scholar] [CrossRef] [Green Version]
- D’Angelo, C.; Costantini, E.; Kamal, M.; Reale, M. Experimental model for ELF-EMF exposure: Concern for human health. Saudi J. Biol. Sci. 2015, 22, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Hei, W.H.; Kim, S.; Park, J.C.; Seo, Y.K.; Kim, S.M.; Jahng, J.W.; Lee, J.H. Schwann-like cells differentiated from human dental pulp stem cells combined with a pulsed electromagnetic field can improve peripheral nerve regeneration. Bioelectromagnetics 2016, 37, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Bao, Z.; Fan, M.; Ma, L.; Duan, Q.; Jiang, W. The effects of pulsed electromagnetic fields combined with a static magnetic intramedullary implant on the repair of bone defects: A preliminary study. Electromagn. Biol. Med. 2019, 38, 210–217. [Google Scholar] [CrossRef]
- Parate, D.; Franco-Obregón, A.; Fröhlich, J.; Beyer, C.; Abbas, A.A.; Kamarul, T.; Hui, J.H.; Yang, Z. Enhancement of mesenchymal stem cell chondrogenesis with short-term low intensity pulsed electromagnetic fields. Sci. Rep. 2017, 7, 9421. [Google Scholar] [CrossRef] [PubMed]
- Andrew, C.; Bassett, L.; Pawluk, R.J.; Pilla, A.A. Augmentation of bone repair by inductively coupled electromagnetic fields. Science 1974, 184, 575–577. [Google Scholar] [CrossRef] [PubMed]
- Martino, C.F.; Belchenko, D.; Ferguson, V.; Nielsen-Preiss, S.; Qi, H.J. The effects of pulsed electromagnetic fields on the cellular activity of SaOS-2 cells. Eur. Bioelectromagn. Assoc. 2008, 29, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Sun, L.; Zhu, B.; Fan, Y.; Ma, X.; Yu, L.; Zhang, J. Pulsed electromagnetic fields promote the proliferation and differentiation of osteoblasts by reinforcing intracellular calcium transients. Bioelectromagnetics 2017, 38, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-O.; Jung, H.; Kim, S.-C.; Park, J.-K.; Seo, Y.-K. Electromagnetic fields and nanomagnetic particles increase the osteogenic differentiation of human bone marrow-derived mesenchymal stem cells. Int. J. Mol. Med. 2015, 35, 153–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jazayeri, M.; Shokrgozar, M.A.; Haghighipour, N.; Bolouri, B.; Mirahmadi, F.; Farokhi, M.J.C.J. Effects of electromagnetic stimulation on gene expression of mesenchymal stem cells and repair of bone lesions. Cell J. 2017, 19, 34. [Google Scholar]
- Lim, K.; Hexiu, J.; Kim, J.; Seonwoo, H.; Cho, W.J.; Choung, P.-H.; Chung, J.H.J.B.r.i. Effects of electromagnetic fields on osteogenesis of human alveolar bone-derived mesenchymal stem cells. BioMed Res. Int. 2013, 2013, 296019. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.H.S.; Chen, L.T.; Sun, J.S.; Lin, F.H. Effect of pulse-burst electromagnetic field stimulation on osteoblast cell activities. Eur. Bioelectromagn. Assoc. 2004, 25, 457–465. [Google Scholar] [CrossRef]
- Hyun, S.-Y.; Lee, J.-H.; Kang, K.-J.; Jang, Y.-J.J.M. Effect of FGF-2, TGF-β-1, and BMPs on teno/ligamentogenesis and osteo/cementogenesis of human periodontal ligament stem cells. Mol. Cells 2017, 40, 550. [Google Scholar] [CrossRef] [Green Version]
- Kwon, S.J.; Kim, Y.M.; Jang, H.J.; Seo, Y.K. Synergistic effect of rice bran extract and extremely low-frequency electromagnetic fields on dermal papilla/melanocytes in melanogenesis. Bioelectromagnetics 2018, 39, 595–603. [Google Scholar] [CrossRef]
- Song, M.; Lee, J.-H.; Bae, J.; Bu, Y.; Kim, E.-C. Human dental pulp stem cells are more effective than human bone marrow-derived mesenchymal stem cells in cerebral ischemic injury. Cell Transplant. 2017, 26, 1001–1016. [Google Scholar] [CrossRef]
- Ledesma-Martínez, E.; Mendoza-Núñez, V.M.; Santiago-Osorio, E. Mesenchymal stem cells derived from dental pulp: A review. Stem Cells Int. 2016, 2016, 4709572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costantini, E.; Sinjari, B.; D’Angelo, C.; Murmura, G.; Reale, M.; Caputi, S. Human Gingival Fibroblasts Exposed to Extremely Low-Frequency Electromagnetic Fields: In Vitro Model of Wound-Healing Improvement. Int. J. Mol. Sci. 2019, 20, 2108. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Chen, P.; Yu, Q.; Peng, Y.; Zhu, Z.; Tian, J. The effects of a pulsed electromagnetic field on the proliferation and osteogenic differentiation of human adipose-derived stem cells. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 3274. [Google Scholar] [CrossRef]
- Leone, L.; Podda, M.V.; Grassi, C. Impact of electromagnetic fields on stem cells: Common mechanisms at the crossroad between adult neurogenesis and osteogenesis. Front. Cell. Neurosci. 2015, 9, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehnert, S.; Schröter, S.; Aspera-Werz, R.H.; Eisler, W.; Falldorf, K.; Ronniger, M.; Nussler, A.K. Translational Insights into Extremely Low Frequency Pulsed Electromagnetic Fields (ELF-PEMFs) for Bone Regeneration after Trauma and Orthopedic Surgery. J. Clin. Med. 2019, 8, 2028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santini, S.J.; Cordone, V.; Falone, S.; Mijit, M.; Tatone, C.; Amicarelli, F.; Di Emidio, G.J. Role of mitochondria in the oxidative stress induced by electromagnetic fields: Focus on reproductive systems. Oxidative Med. Cell. Longev. 2018, 2018, 5076271. [Google Scholar] [CrossRef] [Green Version]
- Khaki, A.A.; Khaki, A.; Gharachour, L.S.; Khourshidi, F.; Taj, A.N.; Madinei, N. Pre and post natal exposure of 50 Hz electromagnetic fields on prostate glands of rats: An electron microscopy study. Iran. J. Reprod. Med. 2008, 6, 77–82. [Google Scholar]
- Korpinar, M.; Kalkan, M.; Tuncel, H.J.B.L.L. The 50 Hz (10 mT) sinusoidal magnetic field: Effects on stress-related behavior of rats. Bratisl. Lek. List. 2012, 113, 521–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, Z.; Simon, B.; Duran, M.; Barabino, G.; Chaudhri, R.; Boyan, B.J. Pulsed electromagnetic fields enhance BMP-2 dependent osteoblastic differentiation of human mesenchymal stem cells. J. Orthop. Res. 2008, 26, 1250–1255. [Google Scholar] [CrossRef]
- Ross, C.L.; Siriwardane, M.; Almeida-Porada, G.; Porada, C.D.; Brink, P.; Christ, G.J.; Harrison, B.S. The effect of low-frequency electromagnetic field on human bone marrow stem/progenitor cell differentiation. Stem Cell Res. 2015, 15, 96–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, K.-J.; Ryu, C.-J.; Jang, Y.-J. Identification of dentinogenic cell-specific surface antigens in odontoblast-like cells derived from adult dental pulp. Stem Cell Res. Ther. 2019, 10, 128. [Google Scholar] [CrossRef] [PubMed]
- Matsui, M.; Kobayashi, T.; Tsutsui, T.W. CD146 positive human dental pulp stem cells promote regeneration of dentin/pulp-like structures. Hum. Cell 2018, 31, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Noda, S.; Kawashima, N.; Yamamoto, M.; Hashimoto, K.; Nara, K.; Sekiya, I.; Okiji, T. Effect of cell culture density on dental pulp-derived mesenchymal stem cells with reference to osteogenic differentiation. Sci. Rep. 2019, 9, 5430. [Google Scholar] [CrossRef]
- Potdar, P.D.; Jethmalani, Y.D. Human dental pulp stem cells: Applications in future regenerative medicine. World J. Stem Cells 2015, 7, 839. [Google Scholar] [CrossRef]
- Semeghini, M.S.; Fernandes, R.R.; Chimello, D.T.; Oliveira, F.S.d.; Bombonato-Prado, K.F. In vitro evaluation of the odontogenic potential of mouse undifferentiated pulp cells. Braz. Dent. J. 2012, 23, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Hanawa, M.; Takano, Y.; Wakita, M. An autoradiographic study of calcium movement in the enamel organ of rat molar tooth germs. Arch. Oral Biol. 1990, 35, 899–906. [Google Scholar] [CrossRef]
- Yu, D.; Zhao, X.; Cheng, J.Z.; Wang, D.; Zhang, H.H.; Han, G.H. Downregulated microRNA-488 enhances odontoblast differentiation of human dental pulp stem cells via activation of the p38 MAPK signaling pathway. J. Cell. Physiol. 2019, 234, 1442–1451. [Google Scholar] [CrossRef]
- Han, N.; Zheng, Y.; Li, R.; Li, X.; Zhou, M.; Niu, Y.; Zhang, Q. β-catenin enhances odontoblastic differentiation of dental pulp cells through activation of Runx2. PLoS ONE 2014, 9, e88890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, D.; Xiao, J.; Zhou, Y.; Zhou, X.; Liu, Y.; Peng, Y.; Yu, Y.; Li, H.; Zhou, X.; Yuan, Q. Epiregulin enhances odontoblastic differentiation of dental pulp stem cells via activating MAPK signalling pathway. Cell Prolif. 2019, 52, e12680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajay Sharma, L.; Ali, M.; Love, R.; Wilson, M.; Dias, G. Novel keratin preparation supports growth and differentiation of odontoblast-like cells. Int. Endod. J. 2016, 49, 471–482. [Google Scholar] [CrossRef]
- Umemura, N.; Ohkoshi, E.; Tajima, M.; Kikuchi, H.; Katayama, T.; Sakagami, H. Hyaluronan induces odontoblastic differentiation of dental pulp stem cells via CD44. Stem Cell Res. Ther. 2016, 7, 135. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.H.; Kim, I.-R.; Kim, Y.; Kim, D.-H.; Park, S.-B.; Park, B.-S.; Bae, M.-K.; Kim, Y.-I. The Effect of Mesoporous Bioactive Glass Nanoparticles/Graphene Oxide Composites on the Differentiation and Mineralization of Human Dental Pulp Stem Cells. Nanomaterials 2020, 10, 620. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Hu, J.; Zhang, G.; Qi, W.; Zhang, P.; Li, P.; Zeng, Y.; Zhao, W.; Tan, Y. Extracellular Ca2+ promotes odontoblastic differentiation of dental pulp stem cells via BMP2-mediated Smad1/5/8 and Erk1/2 pathways. J. Cell. Physiol. 2015, 230, 2164–2173. [Google Scholar] [CrossRef]
- Saito, T.; Ogawa, M.; Hata, Y.; Bessho, K. Acceleration effect of human recombinant bone morphogenetic protein-2 on differentiation of human pulp cells into odontoblasts. J. Endod. 2004, 30, 205–208. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Harris, M.; Cui, Y.; Mishina, Y.; Harris, S.E.; Gluhak-Heinrich, J. Bmp2 is required for odontoblast differentiation and pulp vasculogenesis. J. Dent. Res. 2012, 91, 58–64. [Google Scholar] [CrossRef]
- Lin, W.; Gao, L.; Jiang, W.; Niu, C.; Yuan, K.; Hu, X.; Ma, R.; Huang, Z. The role of osteomodulin on osteo/odontogenic differentiation in human dental pulp stem cells. BMC Oral Health 2019, 19, 22. [Google Scholar] [CrossRef] [Green Version]
- Duan, P.; Bonewald, L. The role of the wnt/β-catenin signaling pathway in formation and maintenance of bone and teeth. Int. J. Biochem. Cell Biol. 2016, 77, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.-M.; Kwon, S.-J.; Jang, H.-j.; Seo, Y.-K. Rice bran mineral extract increases the expression of anagen-related molecules in human dermal papilla through wnt/catenin pathway. Food Nutr. Res. 2017, 61, 1412792. [Google Scholar] [CrossRef] [Green Version]
- Da Rosa, W.; Piva, E.; da Silva, A. Disclosing the physiology of pulp tissue for vital pulp therapy. Int. Endod. J. 2018, 51, 829–846. [Google Scholar] [CrossRef] [Green Version]
- Amirhosseini, M.; Madsen, R.V.; Escott, K.J.; Bostrom, M.P.; Ross, F.P.; Fahlgren, A. GSK-3β inhibition suppresses instability-induced osteolysis by a dual action on osteoblast and osteoclast differentiation. J. Cell. Physiol. 2018, 233, 2398–2408. [Google Scholar] [CrossRef] [PubMed]
- Moon, R.T.; Bowerman, B.; Boutros, M.; Perrimon, N. The promise and perils of Wnt signaling through β-catenin. Science 2002, 296, 1644–1646. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ye, L.; Hui, T.-Q.; Yang, D.-M.; Huang, D.-M.; Zhou, X.-D.; Mao, J.J.; Wang, C.-L. Bone morphogenetic protein 2-induced human dental pulp cell differentiation involves p38 mitogen-activated protein kinase-activated canonical WNT pathway. Int. J. Oral Sci. 2015, 7, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Song, Z.; Huang, S.; Wang, R.; Qin, W.; Guo, J.; Lin, Z. lncRNA DANCR suppresses odontoblast-like differentiation of human dental pulp cells by inhibiting wnt/β-catenin pathway. Cell Tissue Res. 2016, 364, 309–318. [Google Scholar] [CrossRef]
- Rodríguez-Carballo, E.; Gámez, B.; Ventura, F. p38 MAPK signaling in osteoblast differentiation. Front. Cell Dev. Biol. 2016, 4, 40. [Google Scholar] [CrossRef] [Green Version]
- Yun, H.-M.; Kang, S.-K.; Singh, R.K.; Lee, J.-H.; Lee, H.-H.; Park, K.-R.; Yi, J.-K.; Lee, D.-W.; Kim, H.-W.; Kim, E.-C. Magnetic nanofiber scaffold-induced stimulation of odontogenesis and pro-angiogenesis of human dental pulp cells through Wnt/MAPK/NF-κB pathways. Dent. Mater. 2016, 32, 1301–1311. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′-3′) | Reverse(5′-3′) | Temp (°C) | Cycles | Base Pair (bp) |
|---|---|---|---|---|---|
| β-actin | GTGATGGTGGGCATGGGTCA | GCCGGACTCGTCATACTCCT | 58.0 | 37 | 972 |
| BMP2 | GTCCAGCTGTAAGAGACACC | GTACTAGCGACACCCACAAC | 58.5 | 28 | 316 |
| ALP | ATCTCGTTGTCTGAGTACCAGTCC | TGGAGCTTCAGAAGCTCAACACCA | 59.0 | 32 | 454 |
| Runx2 | ACAGTAGATGGACCTCGGGA | ATACTGGGATGAGGAATGCG | 55.0 | 30 | 113 |
| OMD | GCTATGGATGGGCTAGTAAAC | GGGATGTCTTGTAGTTTGTTGTG | 58.5 | 36 | 278 |
| DMP-1 | GAGTGGCTTCATTGGGCATAG | GACTCACTGCTCTCCAAGGG | 60.0 | 37 | 260 |
| DSPP | GGAATGGCTCTAAGTGGGCA | CTCATTGTGACCTGCATCGC | 60.0 | 37 | 284 |
| Markers% | Cont | 40 Hz | 60 Hz | 70 Hz | 150 Hz | |
|---|---|---|---|---|---|---|
| CD73 | 5 D | 77.21 ± 1.47 | 77.50 ± 4.72 | 61.55 ± 1.57 *** | 61.29 ± 0.32 *** | 69.25 ± 2.71 * |
| 10 D | 7.95 ± 0.34 | 7.50 ± 2.14 | 7.27 ± 3.29 | 5.50 ± 1.97 | 4.56 ± 2.12 | |
| CD105 | 5 D | 43.46 ± 1.09 | 42.33 ± 2.87 | 29.24 ± 6.51 * | 29.33 ± 4.34 * | 36.14 ± 3.76 * |
| 10 D | 4.19 ± 0.49 | 3.66 ± 1.22 | 2.82 ± 1.07 | 3.38 ± 0.92 | 2.23 ± 1.04 * | |
| CD146 | 5 D | 40.38 ± 0.53 | 41.34 ± 1.29 | 32.41 ± 2.16 ** | 30.19 ± 3.24 ** | 36.59 ± 4.24 |
| 10 D | 5.54 ± 2.31 | 3.59 ± 1.25 | 1.20 ± 0.46 * | 0.77 ± 0.13 * | 0.99 ± 0.40 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, H.-M.; Nam, M.-H.; Kim, Y.-M.; Seo, Y.-K. Increasing Odontoblast-like Differentiation from Dental Pulp Stem Cells through Increase of β-Catenin/p-GSK-3β Expression by Low-Frequency Electromagnetic Field. Biomedicines 2021, 9, 1049. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9081049
Lim H-M, Nam M-H, Kim Y-M, Seo Y-K. Increasing Odontoblast-like Differentiation from Dental Pulp Stem Cells through Increase of β-Catenin/p-GSK-3β Expression by Low-Frequency Electromagnetic Field. Biomedicines. 2021; 9(8):1049. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9081049
Chicago/Turabian StyleLim, Han-Moi, Myeong-Hyun Nam, Yu-Mi Kim, and Young-Kwon Seo. 2021. "Increasing Odontoblast-like Differentiation from Dental Pulp Stem Cells through Increase of β-Catenin/p-GSK-3β Expression by Low-Frequency Electromagnetic Field" Biomedicines 9, no. 8: 1049. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9081049