
Childhood Obesity-Related Mechanisms: MicroRNome and Transcriptome Changes in a Nested Case-Control Study

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants and Blood Sampling
2.2. Total RNA Extraction and Quality Assurance
2.3. MicroRNome Level Measurements and Data Filtering
2.4. Transcriptome Level Measurements and Data Filtering
2.5. Measurement of Adiponectin and Leptin Levels in Blood
2.6. Statistical Analyses
2.7. Target Gene Prediction of Obesity-Associated miRNAs and Comparison with Obesity-Related mRNAs
2.8. Exploring Networking among miRNAs, mRNAs, Obesity-Related Proteins, and Obesity using Cytoscape
2.9. Exploring Functional Pathways and Diseases Classes
3. Results
3.1. Study Population
3.2. Relations among miRNAs, mRNAs, and Obesity Indicators
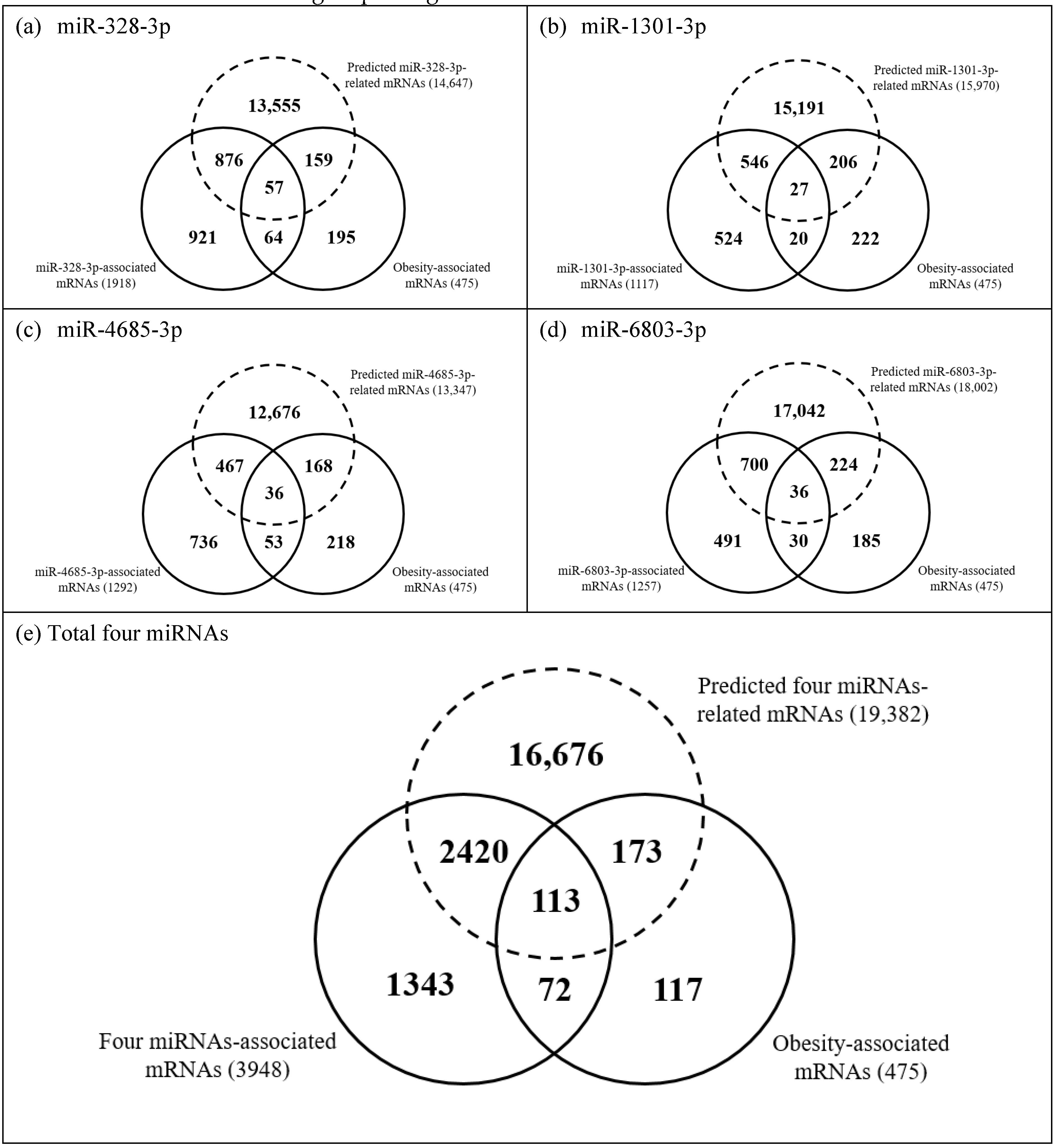
3.3. Prediction for Target Genes of the Four Obesity-Associated miRNAs and Comparison with Four miRNAs-Associated mRNAs or Obesity-Associated mRNAs
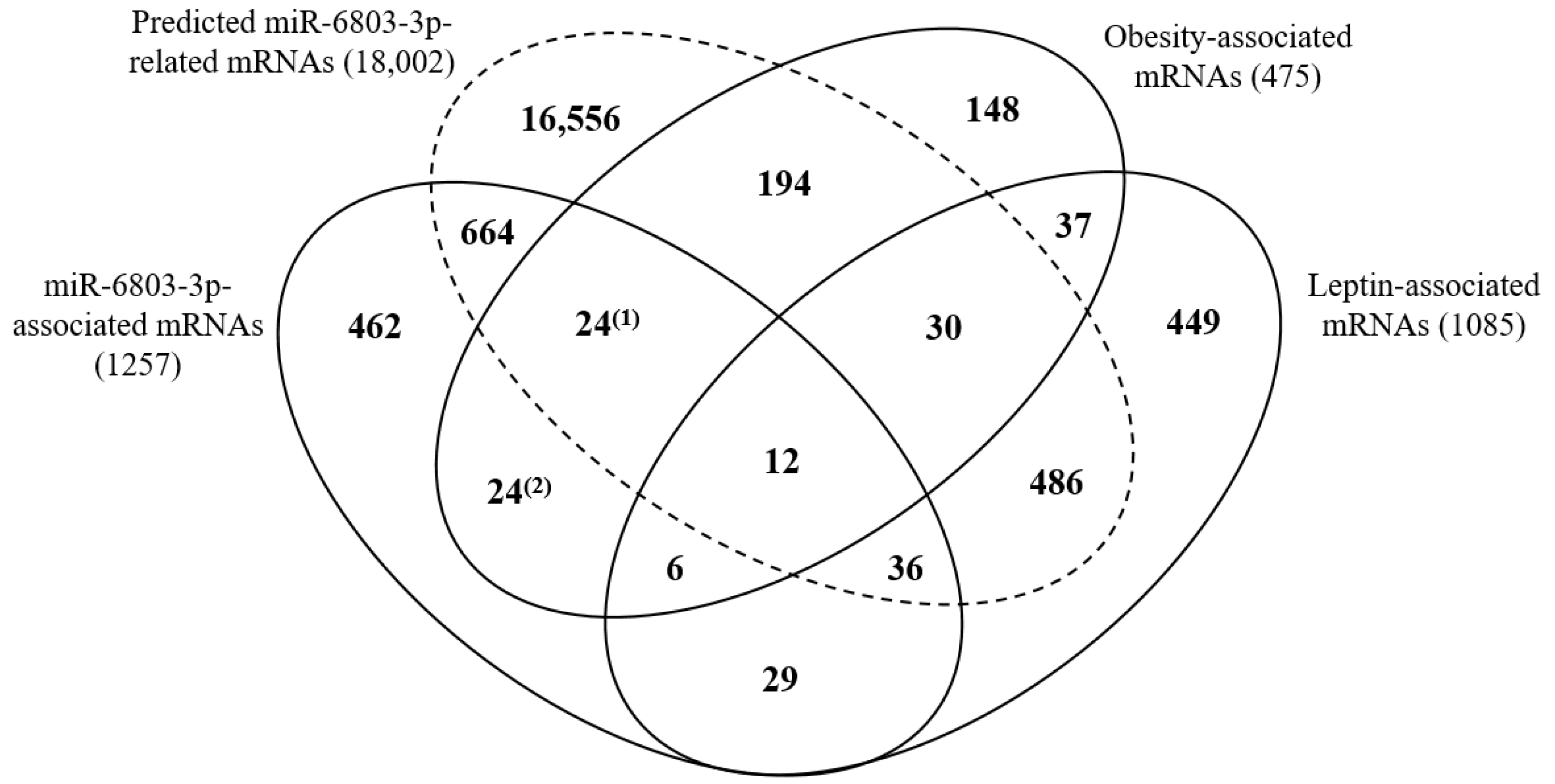
3.4. Relations among miRNAs, mRNAs, and Obesity with Leptin
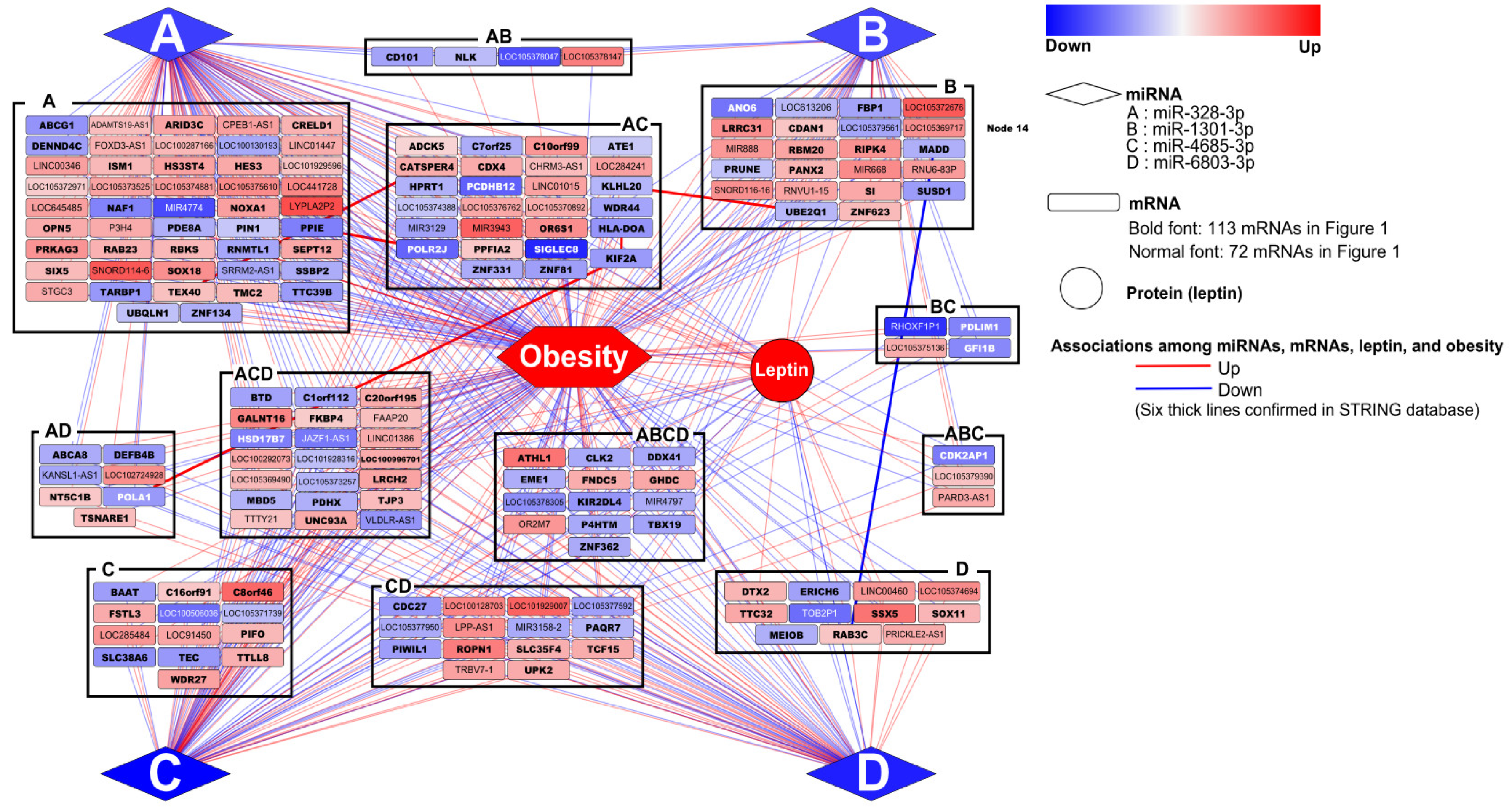
3.5. Networking among miRNAs, mRNAs, and Obesity with Leptin
3.6. KEGG Pathways and GAD Disease Classes Related with Obesity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Despres, J.P.; Lemieux, I. Abdominal obesity and metabolic syndrome. Nature 2006, 444, 881–887. [Google Scholar] [CrossRef]
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef]
- World Health Organization. Health Topics. Obesity. Available online: https://www.who.int/topics/obesity/en/ (accessed on 1 July 2019).
- Kim, K.Y.; Lee, E.; Kim, Y. The association between bisphenol A exposure and obesity in children-a systematic review with meta-analysis. Int. J. Environ. Res. Public Health 2019, 16, 2521. [Google Scholar] [CrossRef] [Green Version]
- Avissar-Whiting, M.; Veiga, K.R.; Uhl, K.M.; Maccani, M.A.; Gagne, L.A.; Moen, E.L.; Marsit, C.J. Bisphenol A exposure leads to specific microRNA alterations in placental cells. Reprod. Toxicol. 2010, 29, 401–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, D.J. The fetal and infant origins of adult disease. Br. Med. J. 1990, 301, 1111. [Google Scholar] [CrossRef] [Green Version]
- Gillman, M.W.; Barker, D.; Bier, D.; Cagampang, F.; Challis, J.; Fall, C.; Godfrey, K.; Gluckman, P.; Hanson, M.; Kuh, D.; et al. Meeting report on the 3rd International Congress on Developmental Origins of Health and Disease (DOHaD). Pediatr. Res. 2007, 61, 625–629. [Google Scholar] [CrossRef] [Green Version]
- Moss, E.G. MicroRNAs: Hidden in the genome. Curr. Biol. 2002, 12, R138–R140. [Google Scholar] [CrossRef] [Green Version]
- Castaño, C.; Kalko, S.; Novials, A.; Párrizas, M. Obesity-associated exosomal miRNAs modulate glucose and lipid metabolism in mice. Proc. Natl. Acad. Sci. USA 2018, 115, 12158–12163. [Google Scholar] [CrossRef] [Green Version]
- Piletic, K.; Kunej, T. MicroRNA epigenetic signatures in human disease. Arch. Toxicol. 2016, 90, 2405–2419. [Google Scholar] [CrossRef] [PubMed]
- Arner, P.; Kulyté, A. MicroRNA regulatory networks in human adipose tissue and obesity. Nat. Rev. Endocrinol. 2015, 11, 276–288. [Google Scholar] [CrossRef] [PubMed]
- Oses, M.; Sanchez, J.M.; Portillo, M.P.; Aguilera, C.M.; Labayen, I. Circulating miRNAs as biomarkers of obesity and obesity-associated comorbidities in children and adolescents: A systematic review. Nutrients 2019, 11, 2890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider-Poetsch, T.; Yoshida, M. Along the central dogma-controlling gene expression with small molecules. Annu. Rev. Biochem. 2018, 87, 391–420. [Google Scholar] [CrossRef] [PubMed]
- Arambula, S.E.; Jima, D.; Patisaul, H.B. Prenatal bisphenol A (BPA) exposure alters the transcriptome of the neonate rat amygdala in a sex-specific manner: A CLARITY-BPA consortium study. Neurotoxicology 2018, 65, 207–220. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Ren, X.M.; Li, Y.Y.; Yao, X.F.; Li, C.H.; Qin, Z.F.; Guo, L.H. Bisphenol A alternatives bisphenol S and bisphenol F interfere with thyroid hormone signaling pathway In Vitro and In Vivo. Environ. Pollut. 2018, 237, 1072–1079. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ji, Y.; Li, M.; Wang, M.; Yi, X.; Yin, C.; Wang, S.; Zhang, M.; Zhao, Z.; Xiao, Y. Integrated analysis of long noncoding RNA and mRNA expression profile in children with obesity by microarray analysis. Sci. Rep. 2018, 8, 8750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.N.; Lim, Y.H.; Shin, C.H.; Lee, Y.A.; Kim, B.N.; Kim, J.I.; Hwang, I.G.; Hwang, M.S.; Suh, J.H.; Hong, Y.C. Cohort profile: The Environment and Development of Children (EDC) study: A prospective children’s cohort. Int. J. Epidemiol. 2018, 47, 1049–1050f. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.S.; Lee, S.Y.; Nam, C.M.; Choi, J.M.; Choe, B.K.; Seo, J.W.; Oh, K.; Jang, M.J.; Hwang, S.S.; Yoo, M.H.; et al. 2007 Korean National Growth Charts: Review of developmental process and an outlook. Korean J. Pediatr. 2008, 51, 1–25. [Google Scholar] [CrossRef]
- Triantafyllou, G.A.; Paschou, S.A.; Mantzoros, C.S. Leptin and hormones: Energy homeostasis. Endocrinol. Metab. Clin. N. Am. 2016, 45, 633–645. [Google Scholar] [CrossRef]
- Salvador, F.; Gomis, R.R. CLK2 blockade modulates alternative splicing compromising MYC-driven breast tumors. EMBO Mol. Med. 2018, 10, e9213. [Google Scholar] [CrossRef]
- Hatting, M.; Rines, A.K.; Luo, C.; Tabata, M.; Sharabi, K.; Hall, J.A.; Verdeguer, F.; Trautwein, C.; Puigserver, P. Adipose tissue CLK2 promotes energy expenditure during high-fat diet intermittent fasting. Cell Metab. 2017, 25, 428–437. [Google Scholar] [CrossRef] [Green Version]
- Quaresma, P.G.F.; Weissmann, L.; Zanotto, T.M.; Santos, A.C.; Matos, A.H.B.; Furigo, I.C.; Simabuco, F.M.; Donato, J., Jr.; Bittencourt, J.C.; Lopes-Cendes, I.; et al. Cdc2-like kinase 2 in the hypothalamus is necessary to maintain energy homeostasis. Int. J. Obes. 2017, 41, 268–278. [Google Scholar] [CrossRef]
- Qin, K.; Jian, D.; Xue, Y.; Cheng, Y.; Zhang, P.; Wei, Y.; Zhang, J.; Xiong, H.; Zhang, Y.; Yuan, X. DDX41 regulates the expression and alternative splicing of genes involved in tumorigenesis and immune response. Oncol. Rep. 2021, 45, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Riviere, T.; Bader, A.; Pogoda, K.; Walzog, B.; Maier-Begandt, D. Structure and emerging functions of LRCH proteins in leukocyte biology. Front. Cell Dev. Biol. 2020, 8, 584134. [Google Scholar] [CrossRef]
- Tajir, M.; Arnoux, J.B.; Boutron, A.; Elalaoui, S.C.; Lonlay, P.D.; Sefiani, A.; Brivet, M. Pyruvate dehydrogenase deficiency caused by a new mutation of PDHX gene in two Moroccan patients. Eur. J. Med. Genet. 2012, 55, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, I.S.; Azmanov, D.N.; Ivanova, M.B.; Chamova, T.; Pacheva, I.H.; Panova, M.V.; Song, S.; Morar, B.; Yordanova, R.V.; Galabova, F.K.; et al. Founder p.Arg 446∗ mutation in the PDHX gene explains over half of cases with congenital lactic acidosis in Roma children. Mol. Genet. Metab. 2014, 113, 76–83. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Yu, X.; Han, X.; Hao, J.; Zhao, J.; Bebek, G.; Bao, S.; Prayson, R.A.; Khalil, A.M.; Jankowsky, E.; et al. Piwil1 regulates glioma stem cell maintenance and glioblastoma progression. Cell Rep. 2021, 34, 108522. [Google Scholar] [CrossRef]
- Koizumi, K.; Oku, M.; Hayashi, S.; Inujima, A.; Shibahara, N.; Chen, L.; Igarashi, Y.; Tobe, K.; Saito, S.; Kadowaki, M.; et al. Identifying pre-disease signals before metabolic syndrome in mice by dynamical network biomarkers. Sci. Rep. 2019, 9, 8767. [Google Scholar] [CrossRef]
- Uphues, I.; Chern, Y.; Eckel, J. Insulin-dependent translocation of the small GTP-binding protein rab3C in cardiac muscle: Studies on insulin-resistant Zucker rats. FEBS Lett. 1995, 377, 109–112. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.A.; Cronk, R.J.; Lang, J.M.; McNeel, D.G. Expression and immunotherapeutic targeting of the SSX family of cancer-testis antigens in prostate cancer. Cancer Res. 2011, 71, 6785–6795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lother, A.; Bergemann, S.; Deng, L.; Moser, M.; Bode, C.; Hein, L. Cardiac endothelial cell transcriptome. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 566–574. [Google Scholar] [CrossRef] [Green Version]
- Coppiello, G.; Collantes, M.; Sirerol-Piquer, M.S.; Vandenwijngaert, S.; Schoors, S.; Swinnen, M.; Vandersmissen, I.; Herijgers, P.; Topal, B.; Loon, J.; et al. Meox2/Tcf15 heterodimers program the heart capillary endothelium for cardiac fatty acid uptake. Circulation 2015, 131, 815–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, Y.M. CD36, a scavenger receptor implicated in atherosclerosis. Exp. Mol. Med. 2014, 46, e99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagerberg, L.; Hallström, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics. Mol. Cell Proteom. 2014, 13, 397–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurada, B.R.V.; Li, L.C.; Mulherkar, N.; Subramanian, M.; Prasad, K.V.; Prabhakar, B.S. MADD, a splice variant of IG20, is indispensable for MAPK activation and protection against apoptosis upon tumor necrosis factor-alpha treatment. J. Biol. Chem. 2009, 284, 13533–13541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mollard, G.F.; Stahl, B.; Khokhlatchev, A.; Südhof, T.C.; Jahn, R. Rab3C is a synaptic vesicle protein that dissociates from synaptic vesicles after stimulation of exocytosis. J. Biol. Chem. 1994, 269, 10971–10974. [Google Scholar] [CrossRef]
- Liu, C.C.; Lin, Y.C.; Chen, Y.H.; Chen, C.M.; Pang, L.Y.; Chen, H.A.; Wu, P.R.; Lin, M.Y.; Jiang, S.T.; Tsai, T.F.; et al. Cul3-KLHL20 ubiquitin ligase governs the turnover of ULK1 and VPS34 complexes to control autophagy termination. Mol. Cell 2016, 61, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Mokarram, P.; Shakiba-Jam, F.; Kavousipour, S.; Sarabi, M.M.; Seghatoleslam, A. Promoter methylation status of two novel human genes, UBE2Q1 and UBE2Q2, in colorectal cancer: A new finding in Iranian patients. Asian Pac. J. Cancer Prev. 2015, 16, 8247–8252. [Google Scholar] [CrossRef] [Green Version]
- Seghatoleslam, A.; Bozorg-Ghalati, F.; Monabati, A.; Nikseresht, M.; Owji, A.A. UBE2Q1, as a down regulated gene in pediatric acute lymphoblastic leukemia. Int. J. Mol. Cell Med. 2014, 3, 95–101. [Google Scholar] [PubMed]
- Uehara, R.; Tsukada, Y.; Kamasaki, T.; Poser, I.; Yoda, K.; Gerlich, D.W.; Goshima, G. Aurora B and Kif2A control microtubule length for assembly of a functional central spindle during anaphase. J. Cell Biol. 2013, 202, 623–636. [Google Scholar] [CrossRef] [Green Version]
- Esch, H.V.; Colnaghi, R.; Freson, K.; Starokadomskyy, P.; Zankl, A.; Backx, L.; Abramowicz, I.; Outwin, E.; Rohena, L.; Faulkner, C.; et al. Defective DNA polymerase alpha-primase leads to X-linked intellectual disability associated with severe growth retardation, microcephaly, and hypogonadism. Am. J. Hum. Genet. 2019, 104, 957–967. [Google Scholar] [CrossRef] [Green Version]
- Lith, M.; Ham, M.; Neefjes, J. Novel polymorphisms in HLA-DOA and HLA-DOB in B-cell malignancies. Immunogenetics 2002, 54, 591–595. [Google Scholar] [CrossRef]
- Belabed, M.; Mauvais, F.X.; Maschalidi, S.; Kurowska, M.; Goudin, N.; Huang, J.D.; Fischer, A.; Basile, G.S.; Endert, P.; Sepulveda, F.E.; et al. Kinesin-1 regulates antigen cross-presentation through the scission of tubulations from early endosomes in dendritic cells. Nat. Commun. 2020, 11, 1817. [Google Scholar] [CrossRef]
- Kim, S.; Gwon, D.; Kim, J.A.; Choi, H.; Jang, C.Y. Bisphenol A disrupts mitotic progression via disturbing spindle attachment to kinetochore and centriole duplication in cancer cell lines. Toxicol. In Vitro 2019, 59, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Proshkin, S.A.; Shematorova, E.K.; Souslova, E.A.; Proshkina, G.M.; Shpakovski, G.V. A minor isoform of the human RNA polymerase II subunit hRPB11 (POLR2J) interacts with several components of the translation initiation factor eIF3. Biochemistry 2011, 76, 976–980. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Han, R.; Zhang, W.; Yuan, Y.; Zhang, X.; Long, Y.; MiHuman, H. CyP33 binds specifically to mRNA and binding stimulates PPIase activity of hCyP33. FEBS Lett. 2008, 582, 835–839. [Google Scholar] [CrossRef] [Green Version]
- Chung, J.J.; Miki, K.; Kim, D.; Shim, S.H.; Shi, H.F.; Hwang, J.Y.; Cai, X.; Iseri, Y.; Zhuang, X.; Clapham, D.E. CatSperz regulates the structural continuity of sperm Ca2+ signaling domains and is required for normal fertility. eLife 2017, 6, e23082. [Google Scholar] [CrossRef]
- Jin, J.L.; O’Doherty, A.M.; Wang, S.; Zheng, H.; Sanders, K.M.; Yan, W. Catsper3 and catsper4 encode two cation channel-like proteins exclusively expressed in the testis. Biol. Reprod. 2005, 73, 1235–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, S.; Ren, F.; Li, L.; Liu, D.; Li, Y.; Wang, A.; Li, W.; Dong, Y.; Guo, W. MiR-328-3p inhibits cell proliferation and metastasis in colorectal cancer by targeting Girdin and inhibiting the PI3K/Akt signaling pathway. Exp. Cell Res. 2020, 390, 111939. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Liu, G.; Guo, J.; Su, Z. The PI3K/AKT pathway in obesity and type 2 diabetes. Int. J. Biol. Sci. 2018, 14, 1483–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, Y.; Zheng, X.; Fu, Y.; Qi, J.; Li, M.; Ma, M.; Wang, S.; Li, S.; Zhu, D. Long noncoding RNA-maternally expressed gene 3 contributes to hypoxic pulmonary hypertension. Mol. Ther. 2019, 27, 2166–2181. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Wang, H.; Dong, T.; Gan, P.; Fang, H.; Wu, S.; Li, J.; Zhang, Y.; Du, R.; Zhu, Q. STAT3-induced upregulation of lncRNA ABHD11-AS1 promotes tumour progression in papillary thyroid carcinoma by regulating miR-1301-3p/STAT3 axis and PI3K/AKT signalling pathway. Cell Prolif. 2019, 52, e12569. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Characteristic, Mean ± SE (Range) or no. (%) | Obese Children (n = 12) | Normal Children (n = 24) | p-Value 1 |
|---|---|---|---|
| Child | |||
| Age, months | 70.1 ± 0.4 (69–73) | 70.1 ± 0.3 (69–73) | 1.0 |
| Sex, no. of boys (%) | 6 (50.0) | 12 (50.0) | 1.0 |
| Weight, kg | 25.6 ± 0.8 (21.5–30.5) | 20.4 ± 0.4 (17.5–25.8) | <0.0001 |
| Height, cm | 118.5 ± 1.2 (112.3–125.1) | 115.9 ± 0.6 (111.1–124.6) | 0.0475 |
| BMI, kg/m2 | 18.2 ± 0.3 (17.0–21.1) | 15.1 ± 0.2 (13.7–17.0) | <0.0001 |
| Vigorous physical activity, mins/day | 32.5 ± 9.9 (0–120) | 41.5 ± 6.1 (0–120) | 0.4245 |
| Calorie intake, kcal/day | 1671.8 ± 129.5 (939.2–2646.4) | 1462.4 ± 66.4 (853.9–2099.9) | 0.1184 |
| Current drug use, no. (%) | |||
| No | 12 (100) | 22 (91.6) | 0.5890 |
| Flu | 0 (0) | 1 (4.2) | |
| Rhinitis | 0 (0) | 1 (4.2) | |
| Adiponectin, µg/mL | 10.2 ± 0.7 (6.0–13.2) | 9.0 ± 0.5 (5.5–17.6) | 0.1744 |
| Leptin, ng/mL | 9.2 ± 1.5 (4.0–21.8) | 5.7 ± 0.6 (3.0–15.6) | 0.0373 |
| Parent | |||
| Mother BMI (before pregnancy), kg/m2 | 21.3 ± 0.7 (16.6–25.6) | 21.1 ± 0.4 (18.1–25.4) | 0.8159 |
| Father BMI, kg/m2 | 25.3 ± 0.9 (19.6–29.8) | 25.5 ± 0.6 (21.0–31.7) | 0.8178 |
| 95% CI | |||||
|---|---|---|---|---|---|
| miRNA | Obesity Indicator | β or OR | Lower CI | Upper CI | p-Value |
| miR-328-3p | BMI | −1.83 | −3.14 | −0.52 | 0.0079 |
| miR-1301-3p | BMI | −1.79 | −3.51 | −0.08 | 0.0412 |
| miR-4685-3p | BMI | −1.49 | −2.63 | −0.35 | 0.0121 |
| miR-6803-3p | BMI | −1.30 | −2.25 | −0.35 | 0.0091 |
| miR-328-3p | BMI z-score | −1.04 | −1.85 | −0.22 | 0.0143 |
| miR-1301-3p | BMI z-score | −1.08 | −2.13 | −0.03 | 0.0444 |
| miR-4685-3p | BMI z-score | −0.89 | −1.59 | −0.19 | 0.0142 |
| miR-6803-3p | BMI z-score | −0.78 | −1.37 | −0.20 | 0.0103 |
| miR-328-3p | Obesity development | 0.04 | 0.003 | 0.59 | 0.0192 |
| miR-1301-3p | Obesity development | 0.06 | 0.004 | 0.97 | 0.0475 |
| miR-4685-3p | Obesity development | 0.07 | 0.01 | 0.62 | 0.0178 |
| miR-6803-3p | Obesity development | 0.07 | 0.01 | 0.75 | 0.0289 |
| 95% CI | |||||
|---|---|---|---|---|---|
| Independent Variable | Dependent Variable | β or OR | Lower CI | Upper CI | p-Value |
| miR-328-3p | Leptin | −1.65 | −4.80 | 1.50 | 0.2936 |
| miR-1301-3p | Leptin | −0.88 | −4.88 | 3.11 | 0.6540 |
| miR-4685-3p | Leptin | −2.01 | −4.65 | 0.64 | 0.1310 |
| miR-6803-3p | Leptin | −2.26 | −4.42 | −0.10 | 0.0413 |
| Leptin | BMI | 0.28 | 0.15 | 0.42 | 0.0002 |
| Leptin | BMI z-score | 0.15 | 0.06 | 0.24 | 0.0020 |
| Leptin | Obesity development | 1.66 | 1.09 | 2.54 | 0.0180 |
| miRNA Name | KEGG Pathways | No. of Database-Matched Genes | No. of Pathway-Related Targets (%) | p-Value |
|---|---|---|---|---|
| miR-328-3p | hsa01100:Metabolic pathways | 15,209 | 921 (6.1) | 0.0030 |
| hsa05200:Pathways in cancer | 15,209 | 348 (2.3) | <0.0001 | |
| hsa04151:PI3K-Akt signaling pathway | 15,209 | 281 (1.8) | 0.0001 | |
| hsa04010:MAPK signaling pathway | 15,209 | 224 (1.5) | <0.0001 | |
| hsa04080:Neuroactive ligand-receptor interaction | 15,209 | 216 (1.4) | 0.0231 | |
| hsa04144:Endocytosis | 15,209 | 213 (1.4) | <0.0001 | |
| hsa05166:HTLV-I infection | 15,209 | 211 (1.4) | 0.0001 | |
| hsa04014:Ras signaling pathway | 15,209 | 199 (1.3) | <0.0001 | |
| hsa04015:Rap1 signaling pathway | 15,209 | 180 (1.2) | <0.0001 | |
| hsa05205:Proteoglycans in cancer | 15,209 | 179 (1.2) | <0.0001 | |
| miR-1301-3p | hsa01100:Metabolic pathways | 16,121 | 980 (6.1) | 0.0074 |
| hsa05200:Pathways in cancer | 16,121 | 356 (2.2) | <0.0001 | |
| hsa04151:PI3K-Akt signaling pathway | 16,121 | 303 (1.9) | <0.0001 | |
| hsa04010:MAPK signaling pathway | 16,121 | 230 (1.4) | <0.0001 | |
| hsa04080:Neuroactive ligand-receptor interaction | 16,121 | 229 (1.4) | 0.0316 | |
| hsa04144:Endocytosis | 16,121 | 222 (1.4) | <0.0001 | |
| hsa05166:HTLV-I infection | 16,121 | 220 (1.4) | 0.0003 | |
| hsa04014:Ras signaling pathway | 16,121 | 205 (1.3) | <0.0001 | |
| hsa04060:Cytokine-cytokine receptor interaction | 16,121 | 201 (1.2) | 0.0426 | |
| hsa04510:Focal adhesion | 16,121 | 192 (1.2) | <0.0001 | |
| miR-4685-3p | hsa05200:Pathways in cancer | 13,811 | 320 (2.3) | <0.0001 |
| hsa04151:PI3K-Akt signaling pathway | 13,811 | 260 (1.9) | 0.0001 | |
| hsa04010:MAPK signaling pathway | 13,811 | 208 (1.5) | <0.0001 | |
| hsa05166:HTLV-I infection | 13,811 | 199 (1.4) | <0.0001 | |
| hsa04144:Endocytosis | 13,811 | 196 (1.4) | <0.0001 | |
| hsa04014:Ras signaling pathway | 13,811 | 186 (1.3) | <0.0001 | |
| hsa04015:Rap1 signaling pathway | 13,811 | 170 (1.2) | <0.0001 | |
| hsa05205:Proteoglycans in cancer | 13,811 | 169 (1.2) | <0.0001 | |
| hsa04510:Focal adhesion | 13,811 | 168 (1.2) | <0.0001 | |
| hsa04024:cAMP signaling pathway | 13,811 | 165 (1.2) | <0.0001 | |
| miR-6803-3p | hsa01100:Metabolic pathways | 17,852 | 1076 (6.0) | 0.0086 |
| hsa05200:Pathways in cancer | 17,852 | 379 (2.1) | <0.0001 | |
| hsa04151:PI3K-Akt signaling pathway | 17,852 | 318 (1.8) | 0.0004 | |
| hsa04080:Neuroactive ligand-receptor interaction | 17,852 | 256 (1.4) | 0.0012 | |
| hsa04010:MAPK signaling pathway | 17,852 | 251 (1.4) | <0.0001 | |
| hsa05166:HTLV-I infection | 17,852 | 237 (1.3) | 0.0004 | |
| hsa04144:Endocytosis | 17,852 | 228 (1.3) | <0.0001 | |
| hsa04014:Ras signaling pathway | 17,852 | 215 (1.2) | <0.0001 | |
| hsa04015:Rap1 signaling pathway | 17,852 | 204 (1.1) | <0.0001 | |
| hsa04810:Regulation of actin cytoskeleton | 17,852 | 202 (1.1) | <0.0001 | |
| Total | hsa01100:Metabolic pathways | 19,922 | 1173 (5.9) | <0.0001 |
| hsa05200:Pathways in cancer | 19,922 | 390 (2.0) | <0.0001 | |
| hsa04151:PI3K-Akt signaling pathway | 19,922 | 334 (1.7) | 0.0016 | |
| hsa04080:Neuroactive ligand-receptor interaction | 19,922 | 272 (1.4) | 0.0001 | |
| hsa04010:MAPK signaling pathway | 19,922 | 253 (1.3) | <0.0001 | |
| hsa05166:HTLV-I infection | 19,922 | 249 (1.2) | 0.0004 | |
| hsa04144:Endocytosis | 19,922 | 237 (1.2) | 0.0002 | |
| hsa04014:Ras signaling pathway | 19,922 | 221 (1.1) | 0.0019 | |
| hsa04810:Regulation of actin cytoskeleton | 19,922 | 208 (1.0) | 0.0001 | |
| hsa04015:Rap1 signaling pathway | 19,922 | 207 (1.0) | 0.0004 |
| miRNA Name | GAD Disease Classes | No. of Database-Matched Genes | No. of Disease Class-Related Targets (%) | p-Value |
|---|---|---|---|---|
| miR-328-3p | METABOLIC | 15,209 | 4014 (26.4) | <0.0001 |
| CARDIOVASCULAR | 15,209 | 3286 (21.6) | <0.0001 | |
| CHEMDEPENDENCY | 15,209 | 2919 (19.2) | <0.0001 | |
| NEUROLOGICAL | 15,209 | 2218 (14.6) | 0.0003 | |
| PHARMACOGENOMIC | 15,209 | 2178 (14.3) | <0.0001 | |
| PSYCH | 15,209 | 1573 (10.3) | <0.0001 | |
| UNKNOWN | 15,209 | 1253 (8.2) | 0.0004 | |
| OTHER | 15,209 | 1242 (8.2) | 0.0002 | |
| DEVELOPMENTAL | 15,209 | 1172 (7.7) | 0.0002 | |
| HEMATOLOGICAL | 15,209 | 1169 (7.7) | 0.0191 | |
| miR-1301-3p | METABOLIC | 16,121 | 4422 (27.4) | <0.0001 |
| CARDIOVASCULAR | 16,121 | 3565 (22.1) | <0.0001 | |
| CHEMDEPENDENCY | 16,121 | 3208 (19.9) | <0.0001 | |
| NEUROLOGICAL | 16,121 | 2396 (14.9) | <0.0001 | |
| PHARMACOGENOMIC | 16,121 | 2318 (14.4) | <0.0001 | |
| INFECTION | 16,121 | 1762 (10.9) | 0.0304 | |
| PSYCH | 16,121 | 1703 (10.6) | <0.0001 | |
| UNKNOWN | 16,121 | 1356 (8.4) | <0.0001 | |
| OTHER | 16,121 | 1332 (8.3) | 0.0003 | |
| DEVELOPMENTAL | 16,121 | 1290 (8.0) | <0.0001 | |
| miR-4685-3p | METABOLIC | 13,811 | 3686 (26.7) | 0.0001 |
| CARDIOVASCULAR | 13,811 | 3017 (21.8) | <0.0001 | |
| CHEMDEPENDENCY | 13,811 | 2752 (19.9) | <0.0001 | |
| NEUROLOGICAL | 13,811 | 2051 (14.9) | 0.0002 | |
| PHARMACOGENOMIC | 13,811 | 2006 (14.5) | <0.0001 | |
| PSYCH | 13,811 | 1448 (10.5) | <0.0001 | |
| UNKNOWN | 13,811 | 1145 (8.3) | 0.0065 | |
| OTHER | 13,811 | 1129 (8.2) | 0.0113 | |
| DEVELOPMENTAL | 13,811 | 1099 (8.0) | <0.0001 | |
| HEMATOLOGICAL | 13,811 | 1088 (7.9) | 0.0046 | |
| miR-6803-3p | METABOLIC | 17,852 | 4678 (26.2) | <0.0001 |
| CARDIOVASCULAR | 17,852 | 3773 (21.1) | <0.0001 | |
| CHEMDEPENDENCY | 17,852 | 3340 (18.7) | <0.0001 | |
| NEUROLOGICAL | 17,852 | 2546 (14.3) | 0.0069 | |
| PHARMACOGENOMIC | 17,852 | 2493 (14.0) | <0.0001 | |
| PSYCH | 17,852 | 1790 (10.0) | <0.0001 | |
| UNKNOWN | 17,852 | 1452 (8.1) | <0.0001 | |
| DEVELOPMENTAL | 17,852 | 1356 (7.6) | <0.0001 | |
| HEMATOLOGICAL | 17,852 | 1356 (7.6) | 0.0074 | |
| RENAL | 17,852 | 1288 (7.2) | <0.0001 | |
| Total | METABOLIC | 19,922 | 4959 (24.9) | <0.0001 |
| CARDIOVASCULAR | 19,922 | 4003 (20.1) | <0.0001 | |
| CHEMDEPENDENCY | 19,922 | 3524 (17.7) | <0.0001 | |
| IMMUNE | 19,922 | 2786 (14.0) | 0.0408 | |
| NEUROLOGICAL | 19,922 | 2701 (13.6) | 0.0189 | |
| PHARMACOGENOMIC | 19,922 | 2650 (13.3) | <0.0001 | |
| INFECTION | 19,922 | 2057 (10.3) | <0.0001 | |
| PSYCH | 19,922 | 1880 (9.4) | <0.0001 | |
| UNKNOWN | 19,922 | 1556 (7.8) | <0.0001 | |
| OTHER | 19,922 | 1508 (7.6) | 0.0006 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.H.; Kim, D.H.; Lim, Y.-H.; Shin, C.H.; Lee, Y.A.; Kim, B.-N.; Kim, J.I.; Hong, Y.-C. Childhood Obesity-Related Mechanisms: MicroRNome and Transcriptome Changes in a Nested Case-Control Study. Biomedicines 2021, 9, 878. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9080878
Kim JH, Kim DH, Lim Y-H, Shin CH, Lee YA, Kim B-N, Kim JI, Hong Y-C. Childhood Obesity-Related Mechanisms: MicroRNome and Transcriptome Changes in a Nested Case-Control Study. Biomedicines. 2021; 9(8):878. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9080878
Chicago/Turabian StyleKim, Jin Hee, Da Hae Kim, Youn-Hee Lim, Choong Ho Shin, Young Ah Lee, Bung-Nyun Kim, Johanna Inhyang Kim, and Yun-Chul Hong. 2021. "Childhood Obesity-Related Mechanisms: MicroRNome and Transcriptome Changes in a Nested Case-Control Study" Biomedicines 9, no. 8: 878. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9080878