
The Role of Non-Coding RNAs in the Regulation of the Proto-Oncogene MYC in Different Types of Cancer

 , and
, and
Abstract
:1. Introduction
2. miRNAs
2.1. miRNAs Controlling the Expression of the MYC Gene in Various Types of Cancer
2.2. miRNAs That Control the Expression of the MYC Gene in Breast Cancer Cells
2.3. miRNAs That Control the Expression of the MYC Gene in the Cells of Tumors of the Digestive System
2.4. miRNAs That Control MYC Gene Expression in Lung Cancer Cells
2.5. miRNAs That Control the Expression of the MYC Gene in Prostate Cancer Cells
2.6. miRNAs That Control MYC Gene Expression in Blood Cancer Cells
2.7. miRNAs That Control the Expression of the MYC Gene in the Cells of Tumors of the Nervous System
2.8. miRNAs That Control the Expression of the MYC Gene in Thyroid Tumor Cells
3. lncRNA
3.1. lncRNAs Controlling MYC Gene Expression in Different Tumor Types
3.2. lncRNAs Controlling MYC Gene Expression in Digestive Tumors
3.3. lncRNAs Controlling MYC Gene Expression in Urinary Tumor Cells
3.4. lncRNAs Controlling MYC Gene Expression in Prostate Cancer Cells
3.5. lncRNAs That Control MYC Gene Expression in Breast Cancer Cells
3.6. lncRNAs That Control MYC Gene Expression in Lung Cancer Cells
3.7. lncRNAs Controlling MYC Gene Expression in Myeloma Cells
3.8. lncRNAs Controlling MYC Gene Expression in Medulloblastoma Cells
4. Circular RNA
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- DePinho, R.; Mitsock, L.; Hatton, K.; Ferrier, P.; Zimmerman, K.; LeGouy, E.; Tesfaye, A.; Collum, R.; Yancopoulos, G.; Nisen, P.; et al. Myc family of cellular oncogenes. J. Cell. Biochem. 1987, 33, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Colby, W.W.; Chen, E.Y.; Smith, D.H.; Levinson, A.D. Identification and nucleotide sequence of a human locus homologous to the v-myc oncogene of avian myelocytomatosis virus MC29. Nat. Cell Biol. 1983, 301, 722–725. [Google Scholar] [CrossRef] [PubMed]
- Kohl, N.; Gee, C.; Alt, F. Activated expression of the N-myc gene in human neuroblastomas and related tumors. Science 1984, 226, 1335–1337. [Google Scholar] [CrossRef]
- Nau, M.M.; Brooks, B.J.; Battey, J.F.; Sausville, E.; Gazdar, A.F.; Kirsch, I.R.; McBride, O.W.; Bertness, V.L.; Hollis, G.F.; Minna, J.D. L-myc, a new myc-related gene amplified and expressed in human small cell lung cancer. Nat. Cell Biol. 1985, 318, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Dang, C.V.; O’Donnell, K.A.; Zeller, K.I.; Nguyen, T.; Osthus, R.C.; Li, F. The c-Myc target gene network. Semin. Cancer Biol. 2006, 16, 253–264. [Google Scholar] [CrossRef]
- Karn, J.; Watson, J.V.; Lowe, A.D.; Green, S.M.; Vedeckis, W. Regulation of cell cycle duration by c-myc levels. Oncogene 1989, 4, 773–787. [Google Scholar]
- Hanson, K.D.; Shichiri, M.; Follansbee, M.R.; Sedivy, J.M. Effects of c-myc expression on cell cycle progression. Mol. Cell. Biol. 1994, 14, 5748–5755. [Google Scholar] [CrossRef] [Green Version]
- Benaud, C.M.; Dickson, R.B. Adhesion-regulated G1 cell cycle arrest in epithelial cells requires the downregulation of c-Myc. Oncogene 2001, 20, 4554–4567. [Google Scholar] [CrossRef] [Green Version]
- Hirning, U.; Schmid, P.; Schulz, W.; Rettenberger, G.; Hameister, H. A comparative analysis of N-myc and c-myc expression and cellular proliferation in mouse organogenesis. Mech. Dev. 1991, 33, 119–125. [Google Scholar] [CrossRef]
- Hirning, U.; Schmid, P.; Schulz, W.; Kozak, L.; Hameister, H. In developing brown adipose tissue c-myc protooncogene expression is restricted to early differentiation stages. Cell Differ. Dev. 1989, 27, 243–248. [Google Scholar] [CrossRef]
- Schmid, P.; Schulz, W.; Hameister, H. Dynamic expression pattern of the myc protooncogene in midgestation mouse embryos. Science 1989, 243, 226–229. [Google Scholar] [CrossRef]
- Douglas, N.; Jacobs, H.; Bothwell, A.L.M.; Hayday, A.C. Defining the specific physiological requirements for c-Myc in T cell development. Nat. Immunol. 2001, 2, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Iritani, B.M.; Eisenman, R.N. c-Myc enhances protein synthesis and cell size during B lymphocyte development. Proc. Natl. Acad. Sci. USA 1999, 96, 13180–13185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loftus, R.; Assmann, N.; Kedia-Mehta, N.; O’Brien, K.L.; Garcia, A.; Gillespie, C.; Hukelmann, J.; Oefner, P.J.; Lamond, A.; Gardiner, C.; et al. Amino acid-dependent cMyc expression is essential for NK cell metabolic and functional responses in mice. Nat. Commun. 2018, 9, 1–15. [Google Scholar] [CrossRef]
- Wei, Y.; Resetca, D.; Li, Z.; Johansson-Åkhe, I.; Ahlner, A.; Helander, S.; Wallenhammar, A.; Morad, V.; Raught, B.; Wallner, B.; et al. Multiple direct interactions of TBP with the MYC oncoprotein. Nat. Struct. Mol. Biol. 2019, 26, 1035–1043. [Google Scholar] [CrossRef]
- Xu, D.; Dwyer, J.; Li, H.; Duan, W.; Liu, J.-P. Ets2 Maintains hTERT Gene Expression and Breast Cancer Cell Proliferation by Interacting with c-Myc. J. Biol. Chem. 2008, 283, 23567–23580. [Google Scholar] [CrossRef] [Green Version]
- Brenner, C.; Deplus, R.; Loriot, A.; De Smet, C.; Gutierrez, A.; Danovi, D.; Bernard, D.; Boon, T.; Pelicci, P.G.; Amati, B.; et al. Myc represses transcription through recruitment of DNA methyltransferase corepressor. EMBO J. 2004, 24, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Ullius, A.; Lüscher-Firzlaff, J.; Costa, I.G.; Walsemann, G.; Forst, A.H.; Gusmao, E.G.; Kapelle, K.; Kleine, H.; Kremmer, E.; Vervoorts, J.; et al. The interaction of MYC with the trithorax protein ASH2L promotes gene transcription by regulating H3K27 modification. Nucleic Acids Res. 2014, 42, 6901–6920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.; Papenhausen, P.; Shao, H. The Role of c-MYC in B-Cell Lymphomas: Diagnostic and Molecular Aspects. Genes 2017, 8, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casulo, C.; Friedberg, J.W. Burkitt lymphoma- a rare but challenging lymphoma. Best Pr. Res. Clin. Haematol. 2018, 31, 279–284. [Google Scholar] [CrossRef]
- Alves, R.D.C.S.; Meurer, R.T.; Roehe, A.V. MYC amplification is associated with poor survival in small cell lung cancer: A chromogenic in situ hybridization study. J. Cancer Res. Clin. Oncol. 2014, 140, 2021–2025. [Google Scholar] [CrossRef] [PubMed]
- Wolfer, A.; Wittner, B.S.; Irimia, D.; Flavin, R.; Lupien, M.; Gunawardane, R.N.; Meyer, C.A.; Lightcap, E.S.; Tamayo, P.; Mesirov, J.P.; et al. MYC regulation of a “poor-prognosis” metastatic cancer cell state. Proc. Natl. Acad. Sci. USA 2010, 107, 3698–3703. [Google Scholar] [CrossRef] [Green Version]
- Pan, W.; Wang, W.; Huang, J.; Lu, K.; Huang, S.; Jiang, D.; Bu, D.; Liu, J.; Jing, H.; Yao, J.; et al. The prognostic role of c-MYC amplification in schistosomiasis-associated colorectal cancer. Jpn. J. Clin. Oncol. 2020, 50, 446–455. [Google Scholar] [CrossRef] [PubMed]
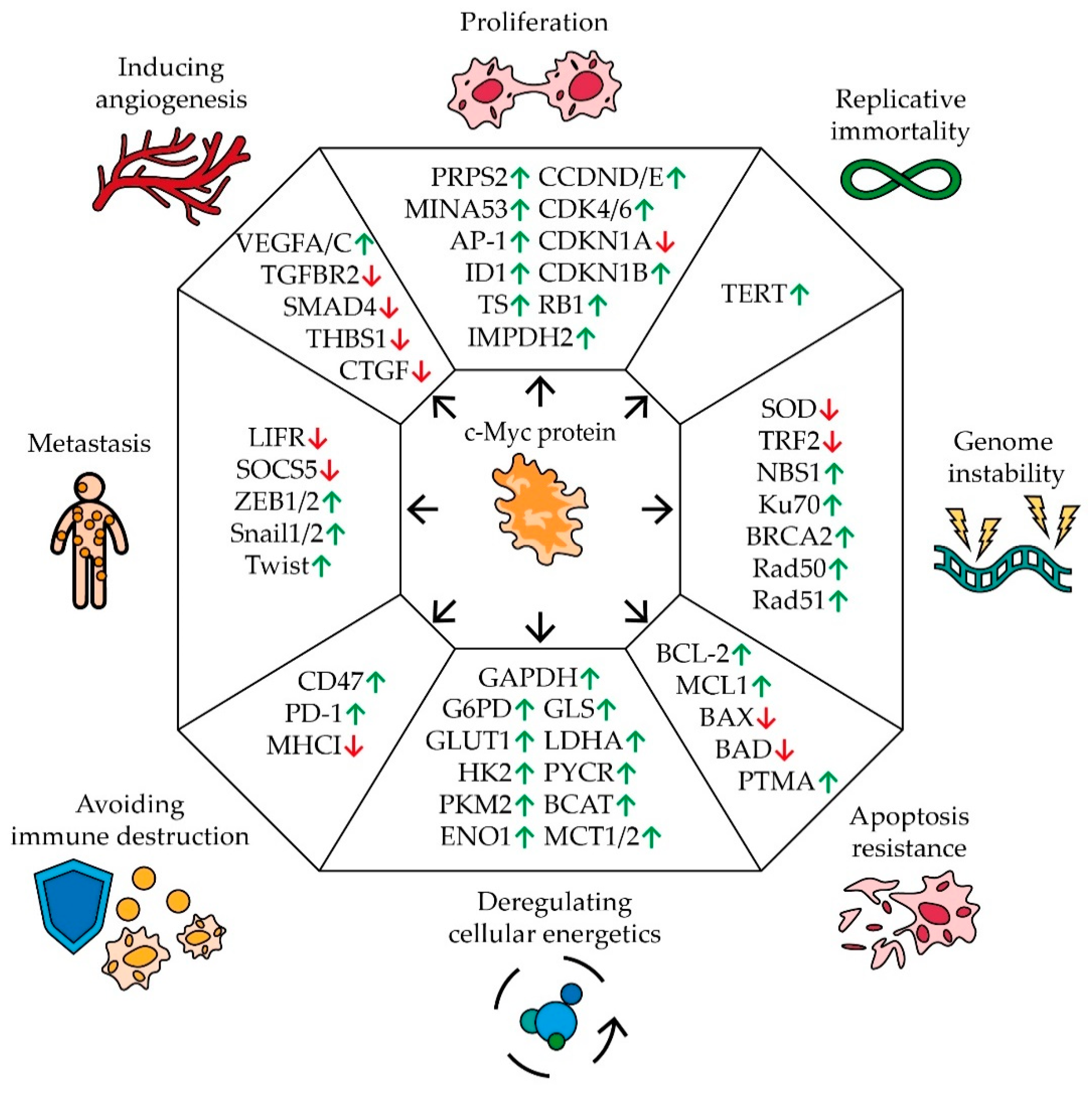
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chanvorachote, P.; Sriratanasak, N.; Nonpanya, N. C-myc Contributes to Malignancy of Lung Cancer: A Potential Anticancer Drug Target. Anticancer. Res. 2020, 40, 609–618. [Google Scholar] [CrossRef]
- Cappellen, D.; Schlange, T.; Bauer, M.; Maurer, F.; Hynes, N. Novel c-MYC target genes mediate differential effects on cell proliferation and migration. EMBO Rep. 2006, 8, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, H.; Li, Z.; Wu, Q.; Lathia, J.D.; McLendon, R.E.; Hjelmeland, A.B.; Rich, J.N. c-Myc Is Required for Maintenance of Glioma Cancer Stem Cells. PLoS ONE 2008, 3, e3769. [Google Scholar] [CrossRef]
- Eilers, M. Control of cell proliferation by Myc family genes. Mol. Cells 1999, 9, 1–6. [Google Scholar]
- Pelengaris, S.; Rudolph, B.; Littlewood, T. Action of Myc in vivo — proliferation and apoptosis. Curr. Opin. Genet. Dev. 2000, 10, 100–105. [Google Scholar] [CrossRef]
- Swier, L.J.Y.M.; Dzikiewicz-Krawczyk, A.; Winkle, M.; Berg, A.V.D.; Kluiver, J. Intricate crosstalk between MYC and non-coding RNA s regulates hallmarks of cancer. Mol. Oncol. 2019, 13, 26–45. [Google Scholar] [CrossRef] [Green Version]
- Aziz, N.; Hong, Y.H.; Jo, M.K.; Kim, J.K.; Kim, K.-H.; Ashktorab, H.; Smoot, D.T.; Hur, H.; Yoo, B.C.; Cho, A.J.Y. Molecular Signatures of JMJD10/MINA53 in Gastric Cancer. Cancers 2020, 12, 1141. [Google Scholar] [CrossRef]
- Tsuneoka, M.; Koda, Y.; Soejima, M.; Teye, K.; Kimura, H. A Novel Myc Target Gene, mina53, That Is Involved in Cell Proliferation. J. Biol. Chem. 2002, 277, 35450–35459. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Zhang, T.; Zhu, Q.; Zhang, P.; Yu, L.; Shen, B.; Yi, W.; Qiu, M.; Zhu, C. Mina53 regulates the differentiation and proliferation of leukemia cells. Hematol. Oncol. 2019, 37, 513–515. [Google Scholar] [CrossRef] [PubMed]
- Swarbrick, A.; Åkerfeldt, M.C.; Lee, C.S.L.; Sergio, C.M.; Caldon, C.E.; Hunter, L.-J.K.; Sutherland, R.L.; Musgrove, E. Regulation of cyclin expression and cell cycle progression in breast epithelial cells by the helix–loop–helix protein Id1. Oncogene 2004, 24, 381–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.-J.; Tsai, J.-W.; Hsieh, K.-C.; Yang, Y.-C.; Chen, Y.-J.; Huang, M.-S.; Yuan, S.-S. Id1 promotes lung cancer cell proliferation and tumor growth through Akt-related pathway. Cancer Lett. 2011, 307, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Mannava, S.; Grachtchouk, V.; Wheeler, L.J.; Im, M.; Zhuang, D.; Slavina, E.G.; Mathews, C.K.; Shewach, D.S.; Nikiforov, M.A. Direct role of nucleotide metabolism in C-MYC-dependent proliferation of melanoma cells. Cell Cycle 2008, 7, 2392–2400. [Google Scholar] [CrossRef] [PubMed]
- Cerni, C. Telomeres, telomerase, and myc. An update. Mutat. Res. Mutat. Res. 2000, 462, 31–47. [Google Scholar] [CrossRef]
- Louis, S.F.; Vermolen, B.J.; Garini, Y.; Young, I.T.; Guffei, A.; Lichtensztejn, Z.; Kuttler, F.; Chuang, T.C.Y.; Moshir, S.; Mougey, V.; et al. c-Myc induces chromosomal rearrangements through telomere and chromosome remodeling in the interphase nucleus. Proc. Natl. Acad. Sci. USA 2005, 102, 9613–9618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ermler, S.; Krunic, D.; Knoch, T.A.; Moshir, S.; Mai, S.; Greulich-Bode, K.M.; Boukamp, P. Cell cycle-dependent 3D distribution of telomeres and telomere repeat-binding factor 2 (TRF2) in HaCaT and HaCaT-myc cells. Eur. J. Cell Biol. 2004, 83, 681–690. [Google Scholar] [CrossRef]
- Okamoto, K.; Bartocci, C.; Ouzounov, I.; Diedrich, J.K.; Iii, J.R.Y.; Denchi, E.L. A two-step mechanism for TRF2-mediated chromosome-end protection. Nat. Cell Biol. 2013, 494, 502–505. [Google Scholar] [CrossRef] [Green Version]
- Vafa, O.; Wade, M.; Kern, S.; Beeche, M.; Pandita, T.K.; Hampton, G.M.; Wahl, G.M. c-Myc Can Induce DNA Damage, Increase Reactive Oxygen Species, and Mitigate p53 Function: A Mechanism for Oncogene-Induced Genetic Instability. Mol. Cell 2002, 9, 1031–1044. [Google Scholar] [CrossRef]
- Tanaka, H.; Matsumura, I.; Ezoe, S.; Satoh, Y.; Sakamaki, T.; Albanese, C.; Machii, T.; Pestell, R.G.; Kanakura, Y. E2F1 and c-Myc Potentiate Apoptosis through Inhibition of NF-κB Activity that Facilitates MnSOD-Mediated ROS Elimination. Mol. Cell 2002, 9, 1017–1029. [Google Scholar] [CrossRef]
- Dang, C.V.; Li, F.; Lee, L.A. Could MYC Induction of Mitochondrial Biogenesis be linked to ROS Production and Genomic Instability? Cell Cycle 2005, 4, 1465–1466. [Google Scholar] [CrossRef] [Green Version]
- Kumari, A.; Folk, W.P.; Sakamuro, D. The Dual Roles of MYC in Genomic Instability and Cancer Chemoresistance. Genes 2017, 8, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, T.; White, J.; Esdale, W.; Burton, M.; DeCruz, E. Tumour cells surviving in vivo cisplatin chemotherapy display elevated c-myc expression. Br. J. Cancer 1996, 73, 610–614. [Google Scholar] [CrossRef] [Green Version]
- Elbadawy, M.; Usui, T.; Yamawaki, H.; Sasaki, K. Emerging Roles of C-Myc in Cancer Stem Cell-Related Signaling and Resistance to Cancer Chemotherapy: A Potential Therapeutic Target Against Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 2340. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Y.; Yang, D.; Yu, Y.; Xiang, M.; Li, H.; Yang, J.; Li, J.; Jiang, D.; Zhou, H.; Xu, Z.; et al. Niclosamide enhances the cytotoxic effect of cisplatin in cisplatin-resistant human lung cancer cells via suppression of lung resistance-related protein and c-myc. Mol. Med. Rep. 2017, 17, 3497–3502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonetti, C.; Biroccio, A.; Candiloro, A.; Citro, G.; Fornari, C.; Mottolese, M.; Del Bufalo, D.; Zupi, G. Increase of cisplatin sensitivity by c-myc antisense oligodeoxynucleotides in a human metastatic melanoma inherently resistant to cisplatin. Clin. Cancer Res. 1999, 5, 2588–2595. [Google Scholar] [PubMed]
- Reyes-González, J.M.; Armaiz-Pena, G.N.; Mangala, L.S.; Valiyeva, F.; Ivan, C.; Pradeep, S.; Echevarría-Vargas, I.M.; Rivera-Reyes, A.; Sood, A.K.; Vivas-Mejía, P.E. Targeting c-MYC in Platinum-Resistant Ovarian Cancer. Mol. Cancer Ther. 2015, 14, 2260–2269. [Google Scholar] [CrossRef] [Green Version]
- Daga, M.; Pizzimenti, S.; Dianzani, C.; Cucci, M.A.; Cavalli, R.; Grattarola, M.; Ferrara, B.; Scariot, V.; Trotta, F.; Barrera, G. Ailanthone inhibits cell growth and migration of cisplatin resistant bladder cancer cells through down-regulation of Nrf2, YAP, and c-Myc expression. Phytomedicine 2019, 56, 156–164. [Google Scholar] [CrossRef]
- Chen, R.; Guo, S.; Yang, C.; Sun, L.; Zong, B.; Li, K.; Liu, L.; Tu, G.; Liu, M.; Liu, S. Although c-MYC contributes to tamoxifen resistance, it improves cisplatin sensitivity in ER-positive breast cancer. Int. J. Oncol. 2020, 56, 932–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.-T.; Lu, H.-P.; Chao, C.C.-K. Oncogenic c-Myc and prothymosin-alpha protect hepatocellular carcinoma cells against sorafenib-induced apoptosis. Biochem. Pharmacol. 2015, 93, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Campaner, S.; Amati, B. Two sides of the Myc-induced DNA damage response: From tumor suppression to tumor maintenance. Cell Div. 2012, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Murphy, D.; Junttila, M.R.; Pouyet, L.; Karnezis, A.; Shchors, K.; Bui, D.A.; Brown-Swigart, L.; Johnson, L.; Evan, G.I. Distinct Thresholds Govern Myc’s Biological Output In Vivo. Cancer Cell 2008, 14, 447–457. [Google Scholar] [CrossRef] [Green Version]
- McMahon, S.B. MYC and the Control of Apoptosis. Cold Spring Harb. Perspect. Med. 2014, 4, a014407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Ye, H.; He, M.; Zhou, X.; Sun, N.; Guo, W.; Lin, X.; Huang, H.; Lin, Y.; Yao, R.; et al. LncRNA PDIA3P interacts with c-Myc to regulate cell proliferation via induction of pentose phosphate pathway in multiple myeloma. Biochem. Biophys. Res. Commun. 2018, 498, 207–213. [Google Scholar] [CrossRef]
- Bose, S.; Le, A. Glucose Metabolism in Cancer. Adv. Exp. Med. Biol. 2018, 1063, 3–12. [Google Scholar] [CrossRef]
- Gan, L.; Xiu, R.; Ren, P.; Yue, M.; Su, H.; Guo, G.; Xiao, D.; Yu, J.; Jiang, H.; Liu, H.; et al. Metabolic targeting of oncogene MYC by selective activation of the proton-coupled monocarboxylate family of transporters. Oncogene 2016, 35, 3037–3048. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Cao, L.; Fan, P.; Mei, Y.; Wu, M. Lnc RNA - MIF, a c-Myc-activated long non-coding RNA, suppresses glycolysis by promoting Fbxw7-mediated c-Myc degradation. EMBO Rep. 2016, 17, 1204–1220. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Tu, R.; Liu, H.; Qing, G. Regulation of cancer cell metabolism: Oncogenic MYC in the driver’s seat. Signal Transduct. Target. Ther. 2020, 5, 1–11. [Google Scholar] [CrossRef]
- Osthus, R.C.; Shim, H.; Kim, S.; Li, Q.; Reddy, R.; Mukherjee, M.; Xu, Y.; Wonsey, D.; Lee, L.A.; Dang, C.V. Deregulation of Glucose Transporter 1 and Glycolytic Gene Expression by c-Myc. J. Biol. Chem. 2000, 275, 21797–21800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehla, K.; Singh, P.K. Metabolic Regulation of Macrophage Polarization in Cancer. Trends Cancer 2019, 5, 822–834. [Google Scholar] [CrossRef] [PubMed]
- Casey, S.C.; Baylot, V.; Felsher, D.W. The MYC oncogene is a global regulator of the immune response. Blood 2018, 131, 2007–2015. [Google Scholar] [CrossRef] [Green Version]
- Casey, S.C.; Tong, L.; Li, Y.; Do, R.; Walz, S.; Fitzgerald, K.N.; Gouw, A.; Baylot, V.; Gütgemann, I.; Eilers, M.; et al. MYC regulates the antitumor immune response through CD47 and PD-L1. Science 2016, 352, 227–231. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zheng, J. Functions of Immune Checkpoint Molecules Beyond Immune Evasion. Adv. Exp. Med. Biol. 2020, 1248, 201–226. [Google Scholar] [CrossRef]
- De Jonge, A.V.; Mutis, T.; Roemer, M.G.M.; Scheijen, B.; Chamuleau, M.E.D. Impact of MYC on Anti-Tumor Immune Responses in Aggressive B Cell Non-Hodgkin Lymphomas: Consequences for Cancer Immunotherapy. Cancers 2020, 12, 3052. [Google Scholar] [CrossRef] [PubMed]
- Wolfer, A.; Ramaswamy, S. MYC and Metastasis: Figure 1. Cancer Res. 2011, 71, 2034–2037. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; Liu, L.; Mao, J.; Zhang, Z.; Wang, Q.; Li, Q. PIM1 mediates epithelial-mesenchymal transition by targeting Smads and c-Myc in the nucleus and potentiates clear-cell renal-cell carcinoma oncogenesis. Cell Death Dis. 2018, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, G.; Wu, X.; Jiang, Z.; Kasman, I.; Yao, J.; Guan, Y.; Oeh, J.; Modrusan, Z.; Bais, C.; Sampath, D.; et al. Tumour-secreted miR-9 promotes endothelial cell migration and angiogenesis by activating the JAK-STAT pathway. EMBO J. 2012, 31, 3513–3523. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Sun, Y.; Wei, Y.; Zhang, P.; Rezaeian, A.H.; Teruya-Feldstein, J.; Gupta, S.; Liang, H.; Lin, H.-K.; Hung, M.-C.; et al. LIFR is a breast cancer metastasis suppressor upstream of the Hippo-YAP pathway and a prognostic marker. Nat. Med. 2012, 18, 1511–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapp, U.R.; Korn, C.; Ceteci, F.; Karreman, C.; Luetkenhaus, K.; Serafin, V.; Zanucco, E.; Castro, I.; Potapenko, T. Myc Is a Metastasis Gene for Non-Small-Cell Lung Cancer. PLoS ONE 2009, 4, e6029. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.Y.; Cha, J.; Kim, S.K.; Park, J.H.; Song, K.H.; Kim, P.; Kim, M.-Y. c-MYC Drives Breast Cancer Metastasis to the Brain, but Promotes Synthetic Lethality with TRAIL. Mol. Cancer Res. 2019, 17, 544–554. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Jin, Y.; Hu, Y.; Jiang, L.; Liu, F.; Zhang, Y.; Hao, Y.; Chen, S.; Wu, X.; Liu, Y. The PLGF /c- MYC /miR-19a axis promotes metastasis and stemness in gallbladder cancer. Cancer Sci. 2018, 109, 1532–1544. [Google Scholar] [CrossRef]
- Song, G.; Li, Y.; Zhang, Z.; Ren, X.; Li, H.; Zhang, W.; Wei, R.; Pan, S.; Shi, L.; Bi, K.; et al. c-myc but not Hif-1α-dependent downregulation of VEGF influences the proliferation and differentiation of HL-60 cells induced by ATRA. Oncol. Rep. 2013, 29, 2378–2384. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.; Chu, P.; Hung, W.; Shan, Y.; Lin, H.; Huang, K.; Chang, J.S.; Chen, L.; Tsai, H. c-Myc promotes lymphatic metastasis of pancreatic neuroendocrine tumor through VEGFC upregulation. Cancer Sci. 2021, 112, 243–253. [Google Scholar] [CrossRef]
- Dews, M.; Fox, J.L.; Hultine, S.; Sundaram, P.; Wang, E.; Liu, Y.Y.; Furth, E.; Enders, G.H.; Eldeiry, W.S.; Schelter, J.M.; et al. The Myc–miR-17∼92 Axis Blunts TGFβ Signaling and Production of Multiple TGFβ-Dependent Antiangiogenic Factors. Cancer Res. 2010, 70, 8233–8246. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.L.; Dews, M.; Minn, A.J.; Thomas-Tikhonenko, A. Targeting of TGF signature and its essential component CTGF by miR-18 correlates with improved survival in glioblastoma. RNA 2013, 19, 177–190. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-H.; Liu, S.; Zhang, G.; Zhou, C.-Q.; Zhu, H.-X.; Zhou, X.-B.; Quan, L.-P.; Bai, J.-F.; Xu, N.-Z. Knockdown of c-Myc expression by RNAi inhibits MCF-7 breast tumor cells growth in vitro and in vivo. Breast Cancer Res. 2005, 7, R220–R228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Ge, Y.-L.; Tian, R.-H. The knockdown of c-myc expression by RNAi inhibits cell proliferation in human colon cancer HT-29 cells in vitro and in vivo. Cell. Mol. Biol. Lett. 2009, 14, 305–318. [Google Scholar] [CrossRef]
- Niu, Z.; Liu, H.; Zhou, M.; Wang, H.; Liu, Y.; Li, X.; Xiong, W.; Ma, J.; Li, G. Knockdown of c-Myc inhibits cell proliferation by negatively regulating the Cdk/Rb/E2F pathway in nasopharyngeal carcinoma cells. Acta Biochim. Et Biophys. Sin. 2015, 47, 183–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Liu, H.; Qing, G. Targeting oncogenic Myc as a strategy for cancer treatment. Signal Transduct. Target. Ther. 2018, 3, 5. [Google Scholar] [CrossRef] [Green Version]
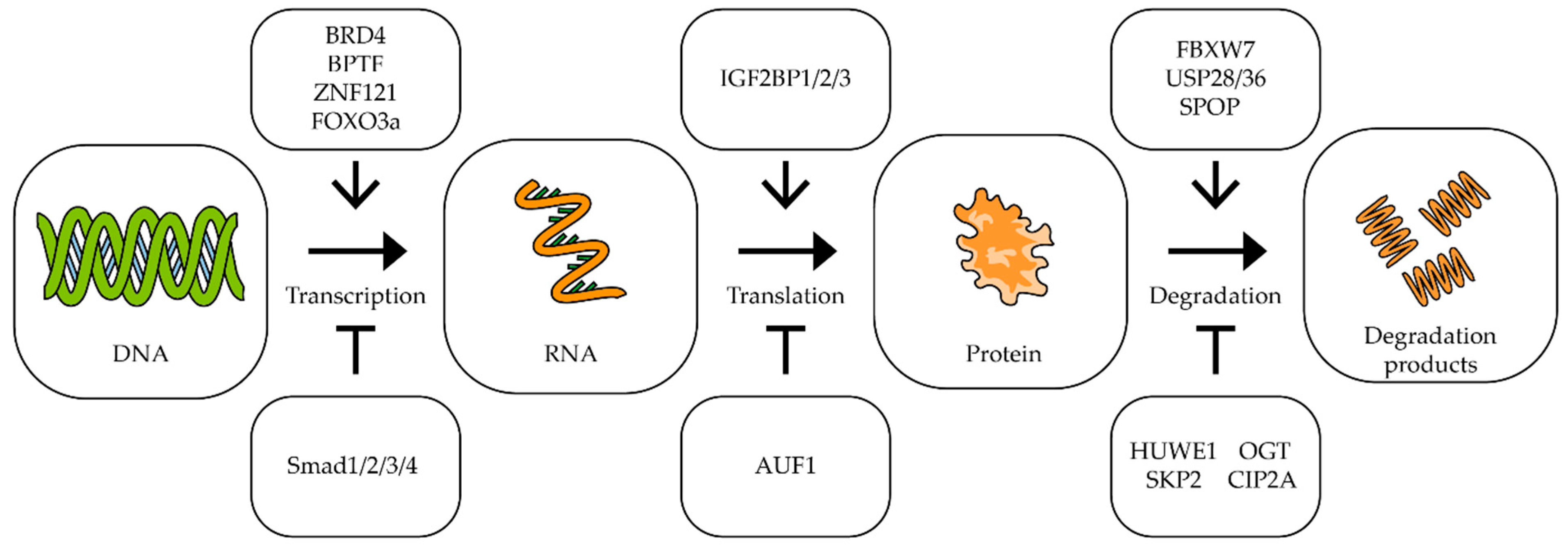
- Otto, C.; Schmidt, S.; Kastner, C.; Denk, S.; Kettler, J.; Müller, N.; Germer, C.; Wolf, E.; Gallant, P.; Wiegering, A. Targeting bromodomain-containing protein 4 (BRD4) inhibits MYC expression in colorectal cancer cells. Neoplasia 2019, 21, 1110–1120. [Google Scholar] [CrossRef]
- Zhang, Y.; Duan, S.; Jang, A.; Mao, L.; Liu, X.; Huang, G. JQ1, a selective inhibitor of BRD4, suppresses retinoblastoma cell growth by inducing cell cycle arrest and apoptosis. Exp. Eye Res. 2021, 202, 108304. [Google Scholar] [CrossRef] [PubMed]
- Andrews, F.H.; Singh, A.R.; Joshi, S.; Smith, C.; Morales, G.; Garlich, J.R.; Durden, D.L.; Kutateladze, T.G. Dual-activity PI3K–BRD4 inhibitor for the orthogonal inhibition of MYC to block tumor growth and metastasis. Proc. Natl. Acad. Sci. USA 2017, 114, E1072–E1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, S.; Singh, A.R.; Liu, K.X.; Pham, T.V.; Zulcic, M.; Skola, D.; Chun, H.B.; Glass, C.K.; Morales, G.A.; Garlich, J.R.; et al. SF2523: Dual PI3K/BRD4 Inhibitor Blocks Tumor Immunosuppression and Promotes Adaptive Immune Responses in Cancer. Mol. Cancer Ther. 2019, 18, 1036–1044. [Google Scholar] [CrossRef] [Green Version]
- Richart, L.; Pau, E.C.D.S.; Río-Machín, A.; de Andres, M.P.; Cigudosa, J.C.; Lobo, V.J.S.-A.; Real, F.X. BPTF is required for c-MYC transcriptional activity and in vivo tumorigenesis. Nat. Commun. 2016, 7, 10153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, L.; Mao, X.; Guo, Y.; Zhou, Y.; Chen, M.; Chen, P.; Yang, S.; Li, L. Discovery of selective BPTF bromodomain inhibitors by screening and structure-based optimization. Biochem. Biophys. Res. Commun. 2021, 545, 125–131. [Google Scholar] [CrossRef]
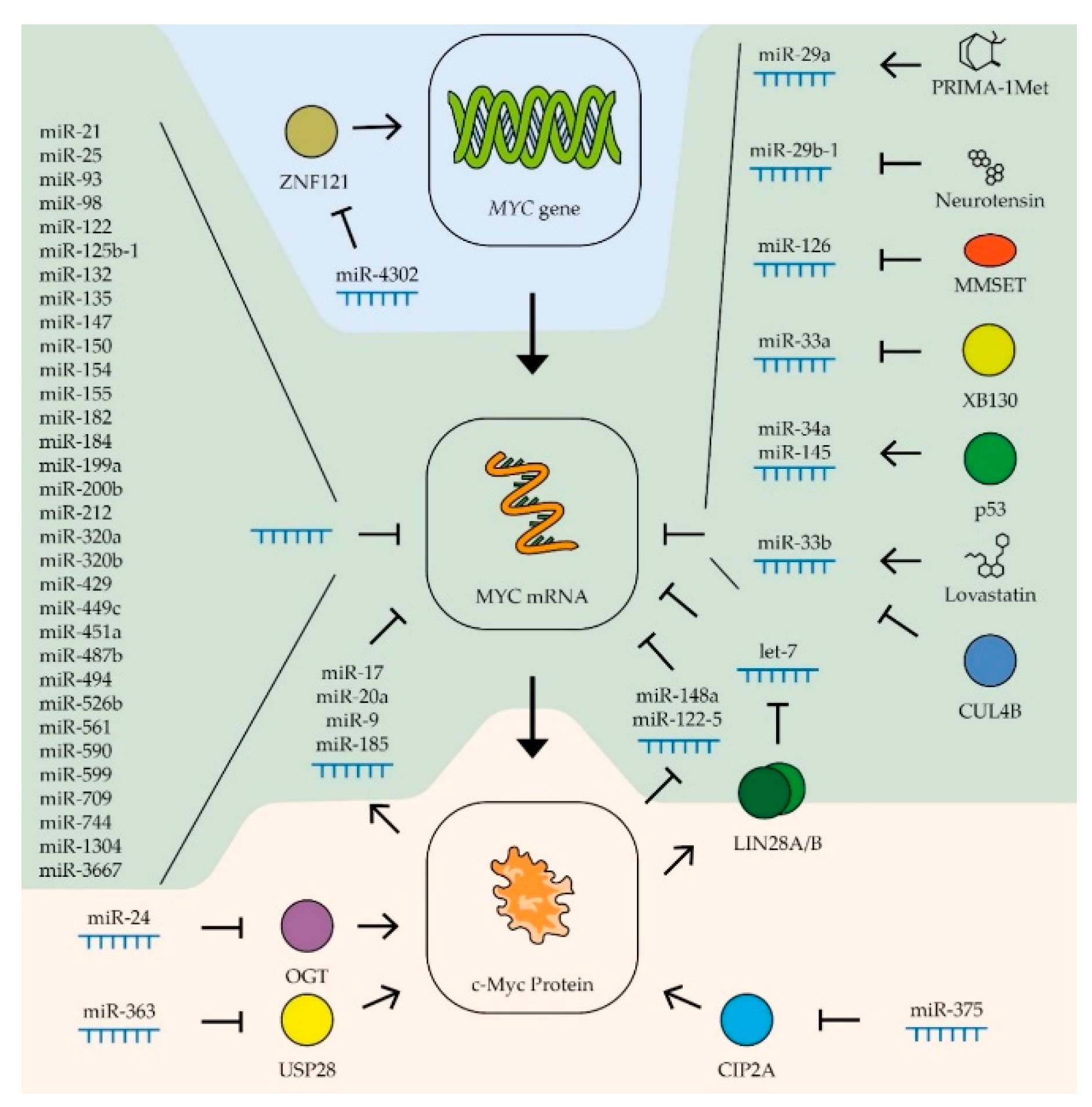
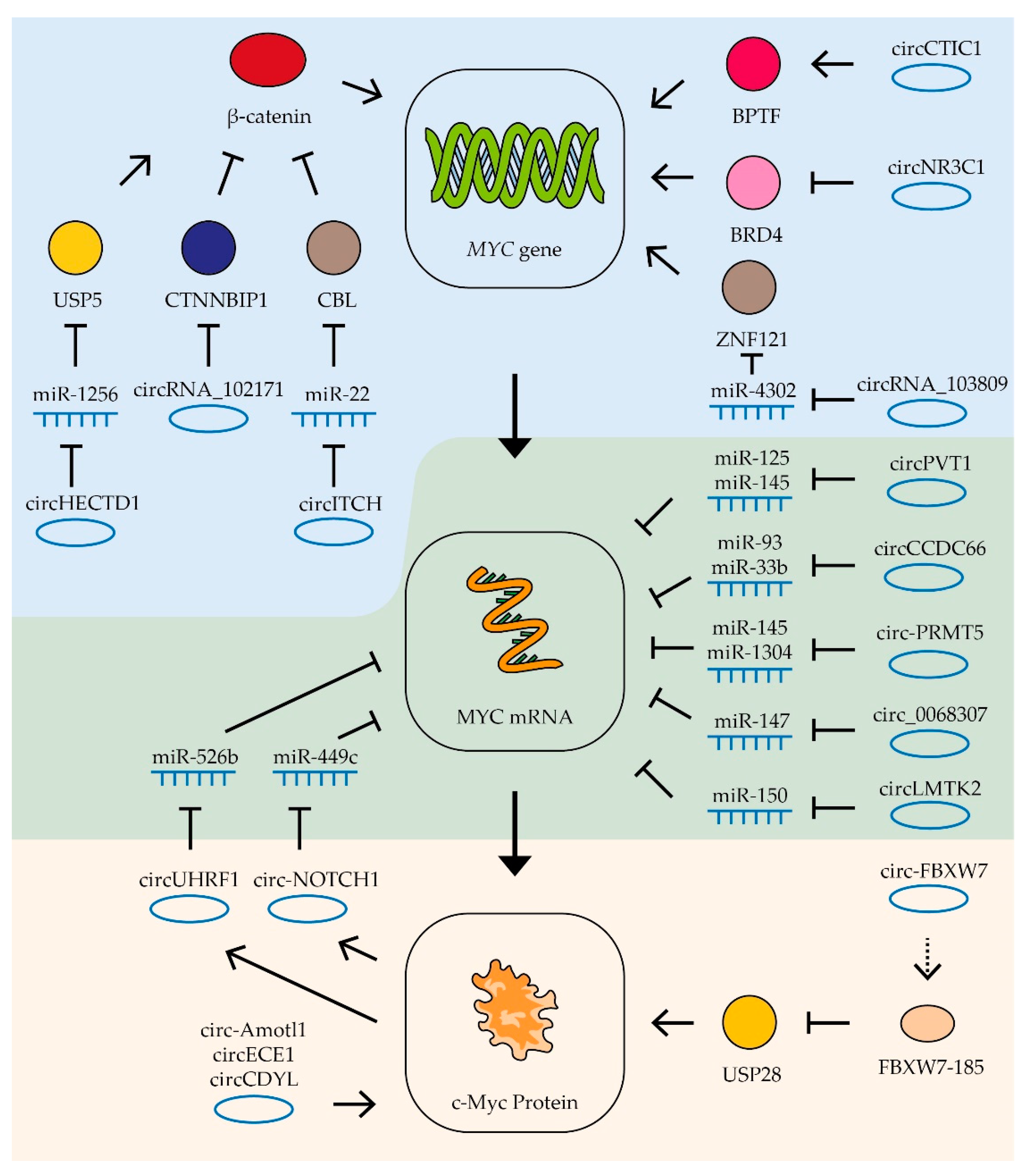
- Liu, W.; Ma, W.; Yuan, Y.; Zhang, Y.; Sun, S. Circular RNA hsa_circRNA_103809 promotes lung cancer progression via facilitating ZNF121-dependent MYC expression by sequestering miR-4302. Biochem. Biophys. Res. Commun. 2018, 500, 846–851. [Google Scholar] [CrossRef]
- Luo, A.; Zhang, X.; Fu, L.; Zhu, Z.; Dong, J.-T. Zinc finger factor ZNF121 is a MYC-interacting protein functionally affecting MYC and cell proliferation in epithelial cells. J. Genet. Genom. 2016, 43, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, S.; Ogawa, K.; Ikei, T.; Fujiki, T.; Katakura, Y. FOXO3a Potentiates hTERT Gene Expression by Activating c-MYC and Extends the Replicative Life-Span of Human Fibroblast. PLoS ONE 2014, 9, e101864. [Google Scholar] [CrossRef] [Green Version]
- Yagi, K.; Furuhashi, M.; Aoki, H.; Goto, D.; Kuwano, H.; Sugamura, K.; Miyazono, K.; Kato, M. c-myc Is a Downstream Target of the Smad Pathway. J. Biol. Chem. 2002, 277, 854–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wierstra, I.; Alves, J. The c-myc Promoter: Still MysterY and Challenge. Adv. Cancer Res. 2008, 99, 113–333. [Google Scholar] [CrossRef]
- Huang, H.; Weng, H.; Sun, W.; Qin, X.; Shi, H.; Wu, H.; Zhao, B.S.; Mesquita, A.; Liu, C.; Yuan, C.L.; et al. Recognition of RNA N6-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat. Cell Biol. 2018, 20, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Liao, B.; Hu, Y.; Brewer, G. Competitive binding of AUF1 and TIAR to MYC mRNA controls its translation. Nat. Struct. Mol. Biol. 2007, 14, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Pan, C.; Zhang, L.; Ding, C.; Chen, F.; Wang, Q.; Wang, K.; Zhang, X. ING4 inhibits the translation of proto-oncogene MYC by interacting with AUF1. FEBS Lett. 2013, 587, 1597–1604. [Google Scholar] [CrossRef]
- Yeh, C.-H.; Bellon, M.; Nicot, C. FBXW7: A critical tumor suppressor of human cancers. Mol. Cancer 2018, 17, 1–19. [Google Scholar] [CrossRef]
- Luo, L.; Tang, H.; Ling, L.; Li, N.; Jia, X.; Zhang, Z.; Wang, X.; Shi, L.; Yin, J.; Qiu, N.; et al. LINC01638 lncRNA activates MTDH-Twist1 signaling by preventing SPOP-mediated c-Myc degradation in triple-negative breast cancer. Oncogene 2018, 37, 6166–6179. [Google Scholar] [CrossRef]
- Crawford, L.J.; Campbell, D.C.; Morgan, J.J.; Lawson, M.A.; Down, J.M.; Chauhan, D.; McAvera, R.M.; Morris, T.C.; Hamilton, C.; Krishnan, A.; et al. The E3 ligase HUWE1 inhibition as a therapeutic strategy to target MYC in multiple myeloma. Oncogene 2020, 39, 5001–5014. [Google Scholar] [CrossRef]
- Sun, X.-X.; He, X.; Yin, L.; Komada, M.; Sears, R.C.; Dai, M.-S. The nucleolar ubiquitin-specific protease USP36 deubiquitinates and stabilizes c-Myc. Proc. Natl. Acad. Sci. USA 2015, 112, 3734–3739. [Google Scholar] [CrossRef] [Green Version]
- Popov, N.; Wanzel, M.; Madiredjo, M.; Zhang, D.; Beijersbergen, R.; Bernards, R.; Moll, R.; Elledge, S.J.; Eilers, M. The ubiquitin-specific protease USP28 is required for MYC stability. Nat. Cell Biol. 2007, 9, 765–774. [Google Scholar] [CrossRef]
- Itkonen, H.M.; Urbanucci, A.; Martin, S.E.; Khan, A.; Mathelier, A.; Thiede, B.; Walker, S.; Mills, I. High OGT activity is essential for MYC-driven proliferation of prostate cancer cells. Theranostics 2019, 9, 2183–2197. [Google Scholar] [CrossRef] [PubMed]
- Itkonen, H.; Minner, S.; Guldvik, I.J.; Sandmann, M.J.; Tsourlakis, M.C.; Berge, V.; Svindland, A.; Schlomm, T.; Mills, I.G. O-GlcNAc Transferase Integrates Metabolic Pathways to Regulate the Stability of c-MYC in Human Prostate Cancer Cells. Cancer Res. 2013, 73, 5277–5287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y.; Mi, W.; Ge, Y.; Liu, H.; Fan, Q.; Han, C.; Yang, J.; Han, F.; Lu, X.; Yu, W. GlcNAcylation Plays an Essential Role in Breast Cancer Metastasis. Cancer Res. 2010, 70, 6344–6351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mi, W.; Gu, Y.; Han, C.; Liu, H.; Fan, Q.; Zhang, X.; Cong, Q.; Yu, W. O-GlcNAcylation is a novel regulator of lung and colon cancer malignancy. Biochim. Et Biophys. Acta BBA Mol. Basis Dis. 2011, 1812, 514–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böckelman, C.; Koskensalo, S.; Hagström, J.; Lundin, M.; Ristimäki, A.; Haglund, C. CIP2A overexpression is associated with c-Myc expression in colorectal cancer. Cancer Biol. Ther. 2012, 13, 289–295. [Google Scholar] [CrossRef]
- Li, W.; Ge, Z.; Liu, C.; Liu, Z.; Björkholm, M.; Jia, J.; Xu, D. CIP2A Is Overexpressed in Gastric Cancer and Its Depletion Leads to Impaired Clonogenicity, Senescence, or Differentiation of Tumor Cells. Clin. Cancer Res. 2008, 14, 3722–3728. [Google Scholar] [CrossRef] [Green Version]
- Vaarala, M.H.; Väisänen, M.-R.; Ristimäki, A. CIP2A expression is increased in prostate cancer. J. Exp. Clin. Cancer Res. 2010, 29, 136. [Google Scholar] [CrossRef] [Green Version]
- Bedewy, A.M.; Elmaghraby, S.M. CIP2A expression in Bortezomib-treated multiple myeloma. J. BUON 2020, 25, 395–400. [Google Scholar]
- Junttila, M.R.; Puustinen, P.; Niemelä, M.; Ahola, R.; Arnold, H.; Böttzauw, T.; Ala-Aho, R.; Nielsen, C.; Ivaska, J.; Taya, Y.; et al. CIP2A Inhibits PP2A in Human Malignancies. Cell 2007, 130, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Russo, F.; Fiscon, G.; Conte, F.; Rizzo, M.; Paci, P.; Pellegrini, M. Interplay Between Long Noncoding RNAs and MicroRNAs in Cancer. In Methods in Molecular Biology; Springer Science and Business Media LLC: Berlin, Germany, 2018; Volume 1819, pp. 75–92. [Google Scholar]
- Chen, X.; Fan, S.; Song, E. Noncoding RNAs: New Players in Cancers. Adv. Exp. Med. Biol. 2016, 927, 1–47. [Google Scholar] [CrossRef]
- Dizaji, B.F. Strategies to target long non-coding RNAs in cancer treatment: Progress and challenges. Egypt. J. Med Hum. Genet. 2020, 21, 1–15. [Google Scholar] [CrossRef]
- Zeng, M.-S.; Song, E. Noncoding RNAs in Cancer Diagnosis. Chem. Biol. Pteridines Folates 2016, 927, 391–427. [Google Scholar] [CrossRef]
- Wei, J.T.; Feng, Z.; Partin, A.W.; Brown, E.; Thompson, I.; Sokoll, L.; Chan, D.W.; Lotan, Y.; Kibel, A.S.; Busby, J.E.; et al. Can Urinary PCA3 Supplement PSA in the Early Detection of Prostate Cancer? J. Clin. Oncol. 2014, 32, 4066–4072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, Y.; Ye, M.; Jiang, X.; Sun, W.; Ding, X.; Liu, Z.; Ye, G.; Zhang, X.; Xiao, B.; Guo, J. Gastric juice long noncoding RNA used as a tumor marker for screening gastric cancer. Cancer 2014, 120, 3320–3328. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.M.; Patel, R.S.; Phillips, B.L.; Wang, H.; Cohen, D.M.; Reinhold, W.; Chang, L.-J.; Yang, L.-J.; Chan, E.K.L. Tumor suppressor miR-375 regulates MYC expression via repression of CIP2A coding sequence through multiple miRNA–mRNA interactions. Mol. Biol. Cell 2013, 24, 1638–1648. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Huang, H.; Liu, M.; Wu, Q.; Li, W.; Zhang, J. MicroRNA-24-1 suppresses mouse hepatoma cell invasion and metastasis via directly targeting O -GlcNAc transferase. Biomed. Pharmacother. 2017, 91, 731–738. [Google Scholar] [CrossRef]
- Kasinski, A.; Slack, F.J. MicroRNAs en route to the clinic: Progress in validating and targeting microRNAs for cancer therapy. Nat. Rev. Cancer 2011, 11, 849–864. [Google Scholar] [CrossRef] [Green Version]
- Shams, R.; Aghdaei, H.A.; Behmanesh, A.; Sadeghi, A.; Zali, M.; Salari, S.; Padrón, J.M. MicroRNAs Targeting MYC Expression: Trace of Hope for Pancreatic Cancer Therapy. A Systematic Review. Cancer Manag. Res. 2020, 12, 2393–2404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampson, V.B.; Rong, N.H.; Han, J.; Yang, Q.; Aris, V.; Soteropoulos, P.; Petrelli, N.J.; Dunn, S.P.; Krueger, L.J. MicroRNA Let-7a Down-regulates MYC and Reverts MYC-Induced Growth in Burkitt Lymphoma Cells. Cancer Res. 2007, 67, 9762–9770. [Google Scholar] [CrossRef] [Green Version]
- Bueno, M.J.; De Cedrón, M.G.; Gómez-López, G.; De Castro, I.P.; Di Lisio, L.; Montes-Moreno, S.; Martínez, N.; Guerrero, M.; Martínez, R.S.; Santos, J.; et al. Combinatorial effects of microRNAs to suppress the Myc oncogenic pathway. Blood 2011, 117, 6255–6266. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, X.; Shi, J.; Pan, Y.; Chen, Q.; Leng, P.; Wang, Y. miR-451 suppresses bladder cancer cell migration and invasion via directly targeting c-Myc. Oncol. Rep. 2016, 36, 2049–2058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, T.; Wang, C.; Xing, J.; Wu, D. miR-429 Modulates the expression of c-myc in human gastric carcinoma cells. Eur. J. Cancer 2011, 47, 2552–2559. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Kluiver, J.; Koerts, J.; De Jong, D.; Rutgers, B.; Razak, F.R.A.; Terpstra, M.; Plaat, B.; Nolte, I.M.; Diepstra, A.; et al. miR-24-3p Is Overexpressed in Hodgkin Lymphoma and Protects Hodgkin and Reed-Sternberg Cells from Apoptosis. Am. J. Pathol. 2017, 187, 1343–1355. [Google Scholar] [CrossRef] [Green Version]
- Su, B.; Xu, T.; Bruce, J.P.; Yip, K.W.; Zhang, N.; Huang, Z.; Zhang, G.; Liu, F.-F.; Liang, J.; Yang, H. hsa-miR-24 suppresses metastasis in nasopharyngeal carcinoma by regulating the c-Myc/epithelial-mesenchymal transition axis. Oncol. Rep. 2018, 40, 2536–2546. [Google Scholar] [CrossRef] [Green Version]
- Khodadadi-Jamayran, A.; Akgol-Oksuz, B.; Afanasyeva, Y.; Heguy, A.; Thompson, M.; Ray, K.; Perafita, A.G.; Sánchez, I.; Wu, X.; Tripathy, D.; et al. Prognostic role of elevated mir-24-3p in breast cancer and its association with the metastatic process. Oncotarget 2018, 9, 12868–12878. [Google Scholar] [CrossRef]
- Liu, D.; Dong, L.; Liu, Y.; Wen, D.; Gao, D.; Sun, H.-C.; Fan, J.; Wu, W. A c-Myc/miR-17-5p feedback loop regulates metastasis and invasion of hepatocellular carcinoma. Tumor Biol. 2016, 37, 5039–5047. [Google Scholar] [CrossRef]
- Guo, Z.; Maki, M.; Ding, R.; Yang, Y.; Zhang, B.; Xiong, L. Genome-wide survey of tissue-specific microRNA and transcription factor regulatory networks in 12 tissues. Sci. Rep. 2014, 4, 5150. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Wan, S.; Yang, Z.; Teschendorff, A.E.; Zou, Q. Tumor origin detection with tissue-specific miRNA and DNA methylation markers. Bioinformatics 2018, 34, 398–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Chan, Z.-L.; Bi, C.; Lu, X.; Chong, P.S.; Chooi, J.-Y.; Cheong, L.-L.; Liu, S.-C.; Ching, Y.Q.; Zhou, Y.; et al. LIN28B Activation by PRL-3 Promotes Leukemogenesis and a Stem Cell–like Transcriptional Program in AML. Mol. Cancer Res. 2017, 15, 294–303. [Google Scholar] [CrossRef] [Green Version]
- Feng, C.; Neumeister, V.; Ma, W.; Xu, J.; Lu, L.; Bordeaux, J.; Maihle, N.J.; Rimm, D.L.; Huang, Y. Lin28 regulates HER2 and promotes malignancy through multiple mechanisms. Cell Cycle 2012, 11, 2486–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Shen, J.; Xie, S.; Jiang, Z.; Huang, L.; Wang, L. Positive expression of Lin28 is correlated with poor survival in gastric carcinoma. Med Oncol. 2013, 30, 382. [Google Scholar] [CrossRef]
- Qiu, J.-L.; Huang, P.-Z.; You, J.-H.; Zou, R.-H.; Wang, L.; Hong, J.; Li, B.-K.; Zhou, K.; Yuan, Y.-F. LIN28 expression and prognostic value in hepatocellular carcinoma patients who meet the Milan criteria and undergo hepatectomy. Chin. J. Cancer 2012, 31, 223–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diskin, S.J.; Capasso, M.; Schnepp, R.W.; Cole, K.A.; Attiyeh, E.F.; Hou, C.; Diamond, M.; Carpenter, E.L.; Winter, C.; Lee, H.; et al. Common variation at 6q16 within HACE1 and LIN28B influences susceptibility to neuroblastoma. Nat. Genet. 2012, 44, 1126–1130. [Google Scholar] [CrossRef]
- Albino, D.; Civenni, G.; Dallavalle, C.; Roos, M.; Jahns, H.; Curti, L.; Rossi, S.; Pinton, S.; D’Ambrosio, G.; Sessa, F.; et al. Activation of the Lin28/let-7 Axis by Loss of ESE3/EHF Promotes a Tumorigenic and Stem-like Phenotype in Prostate Cancer. Cancer Res. 2016, 76, 3629–3643. [Google Scholar] [CrossRef] [Green Version]
- Bhat-Nakshatri, P.; Wang, G.; Collins, N.R.; Thomson, M.J.; Geistlinger, T.R.; Carroll, J.S.; Brown, M.; Hammond, S.; Srour, E.F.; Liu, Y.; et al. Estradiol-regulated microRNAs control estradiol response in breast cancer cells. Nucleic Acids Res. 2009, 37, 4850–4861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Ye, X.-L.; Xu, J.; Cao, M.-G.; Fang, Z.-Y.; Li, L.-Y.; Guan, G.-H.; Liu, Q.; Qian, Y.-H.; Xie, D. The lncRNA H19 mediates breast cancer cell plasticity during EMT and MET plasticity by differentially sponging miR-200b/c and let-7b. Sci. Signal. 2017, 10, eaak9557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, F.; Li, T.-T.; Wang, K.-L.; Xiao, G.-Q.; Wang, J.-H.; Zhao, H.-D.; Kang, Z.-J.; Fan, W.-J.; Zhu, L.-L.; Li, M.; et al. H19/let-7/LIN28 reciprocal negative regulatory circuit promotes breast cancer stem cell maintenance. Cell Death Dis. 2018, 8, e2569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, F.-F.; Wang, H.; Chen, Y.; Chan, C.-Y.; Ng, S.S.; Li, K.; Xie, D.; He, M.-L.; Lin, M.C.; Kung, H.-F. Hsa-let-7g inhibits proliferation of hepatocellular carcinoma cells by downregulation of c-Myc and upregulation of p16INK4A. Int. J. Cancer 2010, 128, 319–331. [Google Scholar] [CrossRef] [PubMed]
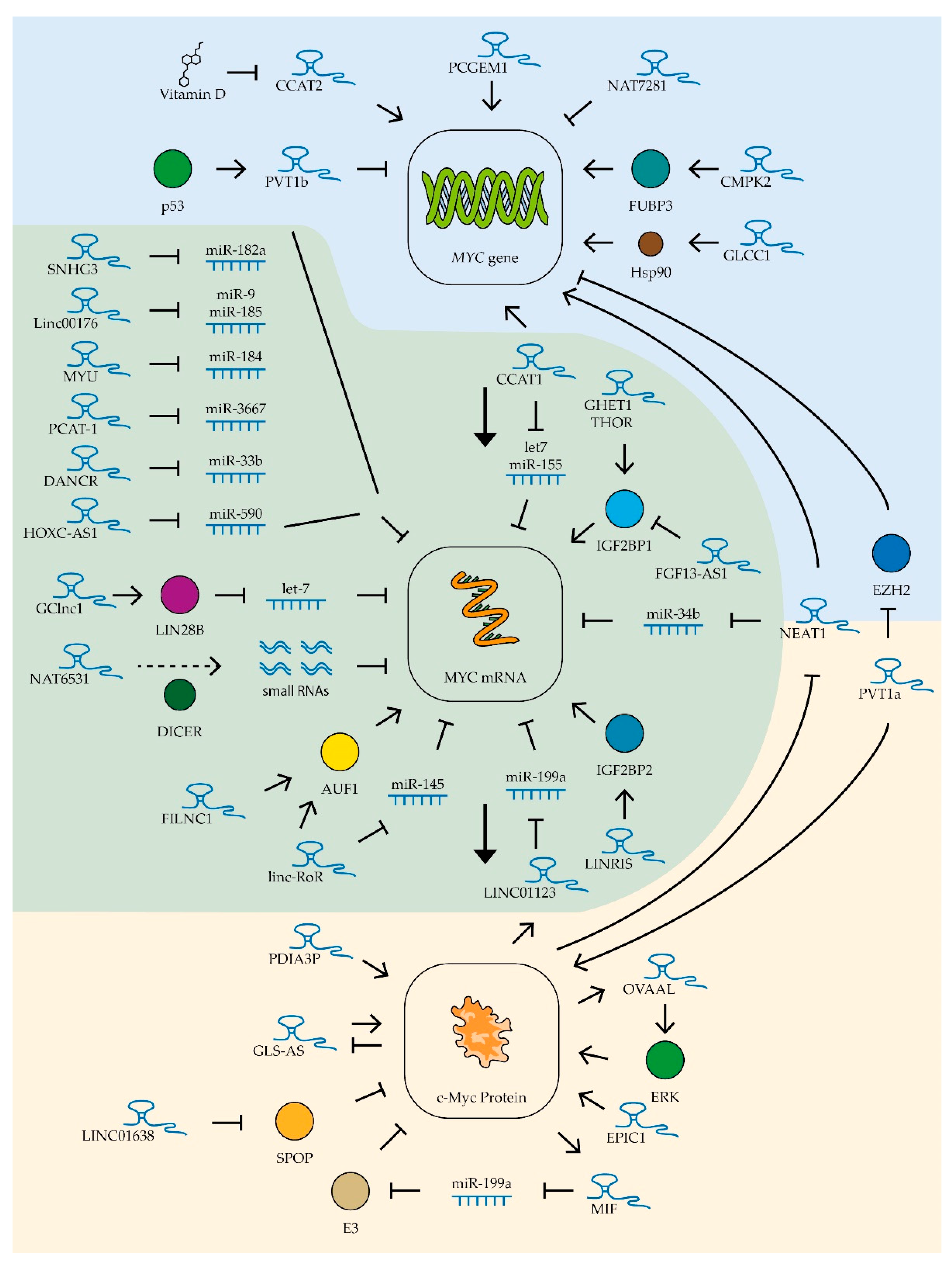
- Deng, L.; Yang, S.-B.; Xu, F.-F.; Zhang, J.-H. Long noncoding RNA CCAT1 promotes hepatocellular carcinoma progression by functioning as let-7 sponge. J. Exp. Clin. Cancer Res. 2015, 34, 1–10. [Google Scholar] [CrossRef] [Green Version]
- He, X.-Y.; Chen, J.-X.; Zhang, Z.; Li, C.-L.; Peng, Q.; Peng, H.-M. The let-7a microRNA protects from growth of lung carcinoma by suppression of k-Ras and c-Myc in nude mice. J. Cancer Res. Clin. Oncol. 2009, 136, 1023–1028. [Google Scholar] [CrossRef]
- Pang, M.; Wu, G.; Hou, X.; Hou, N.; Liang, L.; Jia, G.; Shuai, P.; Luo, B.; Wang, K.; Xiaolin, H. LIN28B Promotes Colon Cancer Migration and Recurrence. PLoS ONE 2014, 9, e109169. [Google Scholar] [CrossRef]
- Akao, Y.; Nakagawa, Y.; Naoe, T. let-7 MicroRNA Functions as a Potential Growth Suppressor in Human Colon Cancer Cells. Biol. Pharm. Bull. 2006, 29, 903–906. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Sun, P.; Wang, C.; Sun, T. Downregulation of microRNA-155 accelerates cell growth and invasion by targeting c-myc in human gastric carcinoma cells. Oncol. Rep. 2014, 32, 951–956. [Google Scholar] [CrossRef]
- He, W.; Li, Y.; Chen, X.; Lü, L.; Tang, B.; Wang, Z.; Pan, Y.; Cai, S.; He, Y.; Ke, Z. miR-494 acts as an anti-oncogene in gastric carcinoma by targeting c-myc. J. Gastroenterol. Hepatol. 2014, 29, 1427–1434. [Google Scholar] [CrossRef]
- Chen, D.; Huang, J.; Zhang, K.; Pan, B.; Chen, J.; De, W.; Wang, R.; Chen, L. MicroRNA-451 induces epithelial–mesenchymal transition in docetaxel-resistant lung adenocarcinoma cells by targeting proto-oncogene c-Myc. Eur. J. Cancer 2014, 50, 3050–3067. [Google Scholar] [CrossRef]
- Roush, S.; Slack, F.J. The let-7 family of microRNAs. Trends Cell Biol. 2008, 18, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.-L.; Deng, S.; Fang, H.-S.; Yu, G.; Peng, H. Hsa-let-7g promotes osteosarcoma by reducing HOXB1 to activate NF-kB pathway. Biomed. Pharmacother. 2019, 109, 2335–2341. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, W.; Cao, L.; Li, Z.; Wang, X. Long Non-Coding RNA CCAT1 Acts as a Competing Endogenous RNA to Regulate Cell Growth and Differentiation in Acute Myeloid Leukemia. Mol. Cells 2016, 39, 330–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Yin, Q.; Mao, Y.; Zhang, Z.; Wu, S.; Cheng, Z.; Chen, X.; Xu, H.; Jin, S.; Jiang, H.; et al. Hsa_circ_0068307 mediates bladder cancer stem cell-like properties via miR-147/c-Myc axis regulation. Cancer Cell Int. 2020, 20, 1–10. [Google Scholar] [CrossRef]
- Leucci, E.; Cocco, M.; Onnis, A.; De Falco, G.; Van Cleef, P.; Bellan, C.; Van Rijk, A.; Nyagol, J.; Byakika, B.; Lazzi, S.; et al. MYC translocation-negative classical Burkitt lymphoma cases: An alternative pathogenetic mechanism involving miRNA deregulation. J. Pathol. 2008, 216, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Wang, C.; Wang, M.; Li, Z.; Casimiro, M.C.; Liu, M.; Wu, K.; Whittle, J.; Ju, X.; Hyslop, T.; et al. A cyclin D1/microRNA 17/20 regulatory feedback loop in control of breast cancer cell proliferation. J. Cell Biol. 2008, 182, 509–517. [Google Scholar] [CrossRef]
- Lujambio, A.; Calin, G.; Villanueva, A.; Ropero, S.; Sanchez-Cespedes, M.; Blanco, D.; Montuenga, L.; Rossi, S.; Nicoloso, M.; Faller, W.; et al. A microRNA DNA methylation signature for human cancer metastasis. Proc. Natl. Acad. Sci. USA 2008, 105, 13556–13561. [Google Scholar] [CrossRef] [Green Version]
- Sachdeva, M.; Zhu, S.; Wu, F.; Wu, H.; Walia, V.; Kumar, S.; Elble, R.; Watabe, K.; Mo, Y.-Y. p53 represses c-Myc through induction of the tumor suppressor miR-145. Proc. Natl. Acad. Sci. USA 2009, 106, 3207–3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, K.-Y.; Lin, Y.-C.; Gupta, S.K.; Chang, N.; Yen, L.; Sun, H.S.; Tsai, S.-J. Noncoding Effects of Circular RNA CCDC66 Promote Colon Cancer Growth and Metastasis. Cancer Res. 2017, 77, 2339–2350. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Cao, F.; Li, X.; Miao, H.; E, J.; Xing, J.; Fu, C.-G. miR-320b suppresses cell proliferation by targeting c-Myc in human colorectal cancer cells. BMC Cancer 2015, 15, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Tian, Y.; Dong, S.; Cha, Y.; Li, J.; Guo, X.; Yuan, X. The long non-coding RNA SNHG3 functions as a competing endogenous RNA to promote malignant development of colorectal cancer. Oncol. Rep. 2017, 38, 1402–1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, Z.; Wei, J.; You, W.; Wang, R.; Shang, J.; Xiong, Y.; Yang, H.; Yang, X.; Fu, Z. Disruption of the c-Myc/miR-200b-3p/PRDX2 regulatory loop enhances tumor metastasis and chemotherapeutic resistance in colorectal cancer. J. Transl. Med. 2017, 15, 1–18. [Google Scholar] [CrossRef]
- Qian, C.-S.; Li, L.-J.; Huang, H.-W.; Yang, H.-F.; Wu, D.-P. MYC-regulated lncRNA NEAT1 promotes B cell proliferation and lymphomagenesis via the miR-34b-5p-GLI1 pathway in diffuse large B-cell lymphoma. Cancer Cell Int. 2020, 20, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Wang, F.; Xu, X.-F.; Mo, W.-H.; Xia, Y.-J.; Wan, R.; Wang, X.-P.; Guo, C.-Y. Down-regulation of miR-212 expression by DNA hypermethylation in human gastric cancer cells. Med. Oncol. 2010, 28, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Song, P.; Su, R.; Yang, G.; Dong, L.; Luo, M.; Wang, B.; Gong, B.; Liu, C.; Song, W.; et al. DNA Methylation mediated down-regulating of MicroRNA-33b and its role in gastric cancer. Sci. Rep. 2016, 6, 18824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Peng, Z.; Zhao, Y.; Chen, L. microRNA-25 Inhibits Cell Apoptosis of Human Gastric Adenocarcinoma Cell Line AGS via Regulating CCNE1 and MYC. Med. Sci. Monit. 2016, 22, 1415–1420. [Google Scholar] [CrossRef]
- Yoon, J.H.; Choi, W.S.; Kim, O.; Choi, B.J.; Nam, S.W.; Lee, J.Y.; Park, W.S. Gastrokine 1 inhibits gastric cancer cell migration and invasion by downregulating RhoA expression. Gastric Cancer 2016, 20, 274–285. [Google Scholar] [CrossRef] [Green Version]
- Qian, K.; Mao, B.; Zhang, W.; Chen, H. MicroRNA-561 inhibits gastric cancercell proliferation and invasion by downregulating c-Myc expression. Am. J. Transl. Res. 2016, 8, 3802–3811. [Google Scholar]
- Dong, Y.; Li, X.; Lin, Z.; Zou, W.; Liu, Y.; Qian, H.; Jia, J. HOXC-AS1-MYC regulatory loop contributes to the growth and metastasis in gastric cancer. J. Exp. Clin. Cancer Res. 2019, 38, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Tang, D.; Wang, W.; Yang, Y.; Wu, X.; Wang, L.; Wang, D. circLMTK2 acts as a sponge of miR-150-5p and promotes proliferation and metastasis in gastric cancer. Mol. Cancer 2019, 18, 1–17. [Google Scholar] [CrossRef]
- Du, W.; Li, D.; Guo, X.; Li, P.; Li, X.; Tong, S.; Tong, J.; Kuang, L.; Liang, D. Circ-PRMT5 promotes gastric cancer progression by sponging miR-145 and miR-1304 to upregulate MYC. Artif. Cells Nanomed. Biotechnol. 2019, 47, 4120–4130. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhong, Q.; Cheng, X.; Wang, S.; Wu, R.; Leng, X.; Shao, L. miR-449c-5p availability is antagonized by circ-NOTCH1 for MYC-induced NOTCH1 upregulation as well as tumor metastasis and stemness in gastric cancer. J. Cell. Biochem. 2020, 121, 4052–4063. [Google Scholar] [CrossRef]
- Chen, J.; Li, Y.; Zheng, Q.; Bao, C.; He, J.; Chen, B.; Lyu, D.; Zheng, B.; Xu, Y.; Long, Z.; et al. Circular RNA profile identifies circPVT1 as a proliferative factor and prognostic marker in gastric cancer. Cancer Lett. 2017, 388, 208–219. [Google Scholar] [CrossRef]
- Ouyang, Q.; Chen, G.; Zhou, J.; Li, L.; Dong, Z.; Yang, R.; Xu, L.; Cui, H.; Xu, M.; Yi, L. Neurotensin signaling stimulates glioblastoma cell proliferation by upregulating c-Myc and inhibiting miR-29b-1 and miR-129-3p. Neuro Oncol. 2015, 18, 216–226. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Zhou, G.; Li, M.; Hu, D.; Zhang, L.; Liu, P.; Lin, K. Long noncoding RNA DANCR mediates cisplatin resistance in glioma cells via activating AXL/PI3K/Akt/NF-κB signaling pathway. Neurochem. Int. 2018, 118, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Sun, C.; Zhou, J.; Wang, Q.; Yu, L.; Bian, X.-W.; Zhou, X.; Hua, D.; Wang, R.; Rao, C.; et al. miR-135a-5p Functions as a Glioma Proliferation Suppressor by Targeting Tumor Necrosis Factor Receptor–Associated Factor 5 and Predicts Patients’ Prognosis. Am. J. Pathol. 2019, 189, 162–176. [Google Scholar] [CrossRef] [Green Version]
- Lo, W.-L.; Yu, C.-C.; Chiou, G.-Y.; Chen, Y.-W.; Huang, P.-I.; Chien, C.-S.; Tseng, L.-M.; Chu, P.-Y.; Lu, K.-H.; Chang, K.-W.; et al. MicroRNA-200c attenuates tumour growth and metastasis of presumptive head and neck squamous cell carcinoma stem cells. J. Pathol. 2011, 223, 482–495. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Sun, D.; Li, W.; Shen, H.; Zhu, Y.; Li, C.; Chen, Y.; Lu, L.; Li, W.; Zhang, J.; et al. A c-Myc-MicroRNA functional feedback loop affects hepatocarcinogenesis. Hepatology 2013, 57, 2378–2389. [Google Scholar] [CrossRef]
- Lin, F.; Ding, R.; Zheng, S.; Xing, D.; Hong, W.; Zhou, Z.; Shen, J. Decrease expression of microRNA-744 promotes cell proliferation by targeting c-Myc in human hepatocellular carcinoma. Cancer Cell Int. 2014, 14, 58. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Hu, X.; Gao, W.; Zhang, J.; Chen, M.; Zhang, X.; Ma, J.; Yuan, H. Identification a novel tumor-suppressive hsa-miR-599 regulates cells proliferation, migration and invasion by targeting oncogenic MYC in hepatocellular carcinoma. Am. J. Transl. Res. 2016, 8, 2575–2584. [Google Scholar]
- Xie, F.; Yuan, Y.; Xie, L.; Ran, P.; Xiang, X.; Huang, Q.; Qi, G.; Guo, X.; Xiao, C.; Zheng, S. miRNA-320a inhibits tumor proliferation and invasion by targeting c-Myc in human hepatocellular carcinoma. OncoTargets Ther. 2017, 10, 885–894. [Google Scholar] [CrossRef] [Green Version]
- Tran, D.D.H.; Kessler, C.; Niehus, S.E.; Mahnkopf, M.; Koch, A.; Tamura, T. Myc target gene, long intergenic noncoding RNA, Linc00176 in hepatocellular carcinoma regulates cell cycle and cell survival by titrating tumor suppressor microRNAs. Oncogene 2018, 37, 75–85. [Google Scholar] [CrossRef]
- Wang, B.; Hsu, S.-H.; Wang, X.; Kutay, H.; Bid, H.K.; Yu, J.; Ganju, R.K.; Jacob, S.T.; Yuneva, M.; Ghoshal, K. Reciprocal regulation of microRNA-122 and c-Myc in hepatocellular cancer: Role of E2F1 and transcription factor dimerization partner 2. Hepatology 2013, 59, 555–566. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Zheng, W.; Tian, P.; He, Y.; Liu, H.; Peng, M.; Li, X.; Liu, X. Oncogenic hsa_circ_0091581 promotes the malignancy of HCC cell through blocking miR-526b from degrading c-MYC mRNA. Cell Cycle 2020, 19, 817–824. [Google Scholar] [CrossRef]
- Chen, Z.; Zeng, H.; Guo, Y.; Liu, P.; Pan, H.; Deng, A.; Hu, J. miRNA-145 inhibits non-small cell lung cancer cell proliferation by targeting c-Myc. J. Exp. Clin. Cancer Res. 2010, 29, 151. [Google Scholar] [CrossRef] [Green Version]
- Xi, S.; Xu, H.; Shan, J.; Tao, Y.; Hong, J.A.; Inchauste, S.; Zhang, M.; Kunst, T.F.; Mercedes, L.; Schrump, D.S. Cigarette smoke mediates epigenetic repression of miR-487b during pulmonary carcinogenesis. J. Clin. Investig. 2013, 123, 1241–1261. [Google Scholar] [CrossRef]
- Miao, L.-J.; Huang, S.-F.; Sun, Z.-T.; Gao, Z.-Y.; Zhang, R.-X.; Liu, Y.; Wang, J. MiR-449c targets c-Myc and inhibits NSCLC cell progression. FEBS Lett. 2013, 587, 1359–1365. [Google Scholar] [CrossRef] [Green Version]
- Bommer, G.; Gerin, I.; Feng, Y.; Kaczorowski, A.J.; Kuick, R.; Love, R.E.; Zhai, Y.; Giordano, T.; Qin, Z.; Moore, B.; et al. p53-Mediated Activation of miRNA34 Candidate Tumor-Suppressor Genes. Curr. Biol. 2007, 17, 1298–1307. [Google Scholar] [CrossRef] [Green Version]
- Hua, Q.; Jin, M.; Mi, B.; Xu, F.; Li, T.; Zhao, L.; Liu, J.; Huang, G. LINC01123, a c-Myc-activated long non-coding RNA, promotes proliferation and aerobic glycolysis of non-small cell lung cancer through miR-199a-5p/c-Myc axis. J. Hematol. Oncol. 2019, 12, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Takwi, A.A.L.; Li, Y.; Buscaglia, L.E.B.; Zhang, J.; Choudhury, S.; Park, A.K.; Liu, M.; Young, K.H.; Park, W.-Y.; Martin, R.C.G.; et al. A statin-regulated microRNA represses human c-Myc expression and function. EMBO Mol. Med. 2012, 4, 896–909. [Google Scholar] [CrossRef]
- Min, D.-J.; Ezponda, T.; Kim, M.K.; Will, C.M.; Martinez-Garcia, E.; Popovic, R.; Basrur, V.; Elenitoba-Johnson, K.; Licht, J.D. MMSET stimulates myeloma cell growth through microRNA-mediated modulation of c-MYC. Leukemia 2012, 27, 686–694. [Google Scholar] [CrossRef] [Green Version]
- Zhen, Y.; Liu, Z.; Yang, H.; Yu, X.; Wu, Q.; Hua, S.; Long, X.; Jiang, Q.; Song, Y.; Cheng, C.; et al. Tumor suppressor PDCD4 modulates miR-184-mediated direct suppression of C-MYC and BCL2 blocking cell growth and survival in nasopharyngeal carcinoma. Cell Death Dis. 2013, 4, e872. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Qu, Y.; Dang, S.; Yao, B.; Ji, M. MiR-145 inhibits oral squamous cell carcinoma (OSCC) cell growth by targeting c-Myc and Cdk6. Cancer Cell Int. 2013, 13, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arunkumar, G.; Rao, A.K.D.M.; Manikandan, M.; Arun, K.; Vinothkumar, V.; Revathidevi, S.; Rajkumar, K.S.; Rajaraman, R.; Munirajan, A.K. Expression profiling of long non-coding RNA identifies linc-RoR as a prognostic biomarker in oral cancer. Tumor Biol. 2017, 39, 1010428317698366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.; Cui, Y.; Liu, L.; Qi, X.; Liu, J.; Ma, S.; Hu, X.; Zhang, Z.; Wang, Y.; Li, H.; et al. Splicing factor derived circular RNA circUHRF1 accelerates oral squamous cell carcinoma tumorigenesis via feedback loop. Cell Death Differ. 2020, 27, 919–933. [Google Scholar] [CrossRef]
- Benassi, B.; Flavin, R.; Marchionni, L.; Zanata, S.; Pan, Y.; Chowdhury, D.; Marani, M.; Strano, S.; Muti, P.; Blandino, G.; et al. MYC Is Activated by USP2a-Mediated Modulation of MicroRNAs in Prostate Cancer. Cancer Discov. 2012, 2, 236–247. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Wang, S.; Zhang, W.; Qiu, J.; Shan, Y.; Yang, D.; Shen, B. Screening key microRNAs for castration-resistant prostate cancer based on miRNA/mRNA functional synergistic network. Oncotarget 2015, 6, 43819–43830. [Google Scholar] [CrossRef] [PubMed]
- Aghdam, A.M.; Amiri, A.; Salarinia, R.; Masoudifar, A.; Ghasemi, F.; Mirzaei, H. MicroRNAs as Diagnostic, Prognostic, and Therapeutic Biomarkers in Prostate Cancer. Crit. Rev. Eukaryot. Gene Expr. 2019, 29, 127–139. [Google Scholar] [CrossRef]
- Prensner, J.; Chen, W.; Han, S.; Iyer, M.K.; Cao, Q.; Kothari, V.; Evans, J.R.; Knudsen, K.; Paulsen, M.T.; Ljungman, M.; et al. The Long Non-Coding RNA PCAT-1 Promotes Prostate Cancer Cell Proliferation through cMyc. Neoplasia 2014, 16, 900–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, A.; Zhao, Q.; Zhou, X.; Sun, C.; Si, J.; Zhou, R.; Gan, L.; Zhang, H. MicroRNA-449a enhances radiosensitivity by downregulation of c-Myc in prostate cancer cells. Sci. Rep. 2016, 6, 27346. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Qi, M.; Li, X.; Hu, J.; Zhang, J.; Jiao, M.; Bai, X.; Peng, X.; Han, B. CUL4B/miR-33b/C-MYC axis promotes prostate cancer progression. Prostate 2019, 79, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yang, X.; Li, R.; Wang, L.; Gu, Y.; Zhao, Y.; Huang, K.H.; Cheng, T.; Yuan, Y.; Gao, S. Long non-coding RNA MYU promotes prostate cancer proliferation by mediating the miR-184/c-Myc axis. Oncol. Rep. 2018, 40, 2814–2825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamura, S.; Saini, S.; Majid, S.; Hirata, H.; Ueno, K.; Chang, I.; Tanaka, Y.; Gupta, A.; Dahiya, R. MicroRNA-34a suppresses malignant transformation by targeting c-Myc transcriptional complexes in human renal cell carcinoma. Carcinogenesis 2011, 33, 294–300. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Sanda, T.; Look, A.T.; Novina, C.D.; Von Boehmer, H. Repression of tumor suppressor miR-451 is essential for NOTCH1-induced oncogenesis in T-ALL. J. Exp. Med. 2011, 208, 663–675. [Google Scholar] [CrossRef]
- Marques, J.C.; Ricarte-Filho, J.C.M.; Fuziwara, C.S.; Yamashita, A.S.; Rezende, E.; Da-Silva, M.J.; Kimura, E.T. Effects of let-7 microRNA on Cell Growth and Differentiation of Papillary Thyroid Cancer. Transl. Oncol. 2009, 2, 236–241. [Google Scholar] [CrossRef]
- Takeshita, H.; Shiozaki, A.; Bai, X.-H.; Iitaka, D.; Kim, H.; Yang, B.B.; Keshavjee, S.; Liu, M. XB130, a New Adaptor Protein, Regulates Expression of Tumor Suppressive MicroRNAs in Cancer Cells. PLoS ONE 2013, 8, e59057. [Google Scholar] [CrossRef] [PubMed]
- Balzeau, J.; Menezes, M.R.; Cao, S.; Hagan, J.P. The LIN28/let-7 Pathway in Cancer. Front. Genet. 2017, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Cai, Q.; Medeiros, L.J.; Xu, X.; Young, K.H. MYC-driven aggressive B-cell lymphomas: Biology, entity, differential diagnosis and clinical management. Oncotarget 2015, 6, 38591–38616. [Google Scholar] [CrossRef] [Green Version]
- Saha, M.N.; Abdi, J.; Yang, Y.; Chang, H. miRNA-29a as a tumor suppressor mediates PRIMA-1Met-induced anti-myeloma activity by targeting c-Myc. Oncotarget 2016, 7, 7149–7160. [Google Scholar] [CrossRef]
- Sang, B.; Zhang, Y.; Guo, S.T.; Kong, L.F.; Cheng, Q.; Liu, G.; Thorne, R.F.; Zhang, X.D.; Jin, L.; Wu, M. Dual functions for OVAAL in initiation of RAF/MEK/ERK prosurvival signals and evasion of p27-mediated cellular senescence. Proc. Natl. Acad. Sci. USA 2018, 115, E11661–E11670. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.-J.; Chen, H.-Y.; Zeng, Z.; Deng, S.; Zhu, S.; Ye, Z.; He, C.; Liu, M.-L.; Huang, K.; Zhong, J.-X.; et al. Nutrient Stress–Dysregulated Antisense lncRNA GLS-AS Impairs GLS-Mediated Metabolism and Represses Pancreatic Cancer Progression. Cancer Res. 2019, 79, 1398–1412. [Google Scholar] [CrossRef] [PubMed]
- Nissan, A.; Stojadinovic, A.; Mitrani-Rosenbaum, S.; Halle, D.; Grinbaum, R.; Roistacher, M.; Bochem, A.; Dayanc, B.E.; Ritter, G.; Gomceli, I.; et al. Colon cancer associated transcript-1: A novel RNA expressed in malignant and pre-malignant human tissues. Int. J. Cancer 2012, 130, 1598–1606. [Google Scholar] [CrossRef] [PubMed]
- Cabanski, C.R.; White, N.M.; Dang, H.X.; Silva-Fisher, J.M.; Rauck, C.E.; Cicka, D.; Maher, C.A. Pan-cancer transcriptome analysis reveals long noncoding RNAs with conserved function. RNA Biol. 2015, 12, 628–642. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wang, W.; Lin, J.; Xiao, J.; Tian, Y. lncRNA CCAT1 promotes bladder cancer cell proliferation, migration and invasion. Int. Braz. J. Urol. 2019, 45, 549–559. [Google Scholar] [CrossRef] [PubMed]
- You, Z.; Liu, C.; Wang, C.; Ling, Z.; Wang, Y.; Wang, Y.; Zhang, M.; Chen, S.; Xu, B.; Guan, H.; et al. LncRNA CCAT1 Promotes Prostate Cancer Cell Proliferation by Interacting with DDX5 and MIR-28-5P. Mol. Cancer Ther. 2019, 18, 2469–2479. [Google Scholar] [CrossRef] [Green Version]
- Shan, L.; Liu, W.; Zhan, Y. Long Non-coding RNA CCAT1 Acts as an Oncogene and Promotes Sunitinib Resistance in Renal Cell Carcinoma. Front. Oncol. 2020, 10, 516552. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Tan, X.; Wang, X.; Jin, H.; Liu, L.; Ma, L.; Yu, H.; Fan, Z. C-Myc-activated long noncoding RNA CCAT1 promotes colon cancer cell proliferation and invasion. Tumor Biol. 2014, 35, 12181–12188. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.-Z.; Chu, B.-F.; Zhang, Y.; Weng, M.-Z.; Qin, Y.-Y.; Gong, W.; Quan, Z.-W. Long non-coding RNA CCAT1 promotes gallbladder cancer development via negative modulation of miRNA-218-5p. Cell Death Dis. 2015, 6, e1583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.-P.; Yang, J.-X.; Cao, D.-Y.; Shen, K. Identification of differentially expressed long non-coding RNAs in human ovarian cancer cells with different metastatic potentials. Cancer Biol. Med. 2013, 10, 138–141. [Google Scholar] [CrossRef]
- Wu, M.; Shen, J. From Super-Enhancer Non-coding RNA to Immune Checkpoint: Frameworks to Functions. Front. Oncol. 2019, 9, 1307. [Google Scholar] [CrossRef]
- Zhuang, C.; Ma, Q.; Zhuang, C.; Ye, J.; Zhang, F.; Gui, Y. LncRNA GClnc1 promotes proliferation and invasion of bladder cancer through activation of MYC. FASEB J. 2019, 33, 11045–11059. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Zhang, H.; Ke, J.; Shen, H.; Ou, H.; Liu, Y. Overexpression of LncRNA GHET1 predicts an unfavourable survival and clinical parameters of patients in various cancers. J. Cell. Mol. Med. 2019, 23, 4891–4899. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Li, W.; Jin, X.; Niu, T.; Cao, Y.; Zhou, P.; Zheng, M. Silencing long non-coding RNA NEAT1 enhances the suppression of cell growth, invasion, and apoptosis of bladder cancer cells under cisplatin chemotherapy. Int. J. Clin. Exp. Pathol. 2019, 12, 549–558. [Google Scholar]
- Zhuang, C.; Li, J.; Liu, Y.; Chen, M.; Yuan, J.; Fu, X.; Zhan, Y.; Liu, L.; Lin, J.; Zhou, Q.; et al. Tetracycline-inducible shRNA targeting long non-coding RNA PVT1 inhibits cell growth and induces apoptosis in bladder cancer cells. Oncotarget 2015, 6, 41194–41203. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yang, B.; Zhang, M.; Guo, W.; Wu, Z.; Wang, Y.; Jia, L.; Li, S.; Xie, W.; Yang, D.; et al. lncRNA Epigenetic Landscape Analysis Identifies EPIC1 as an Oncogenic lncRNA that Interacts with MYC and Promotes Cell-Cycle Progression in Cancer. Cancer Cell 2018, 33, 706–720.e9. [Google Scholar] [CrossRef] [Green Version]
- Ma, F.; Liu, X.; Zhou, S.; Li, W.; Liu, C.; Chadwick, M.; Qian, C. Long non-coding RNA FGF13-AS1 inhibits glycolysis and stemness properties of breast cancer cells through FGF13-AS1/IGF2BPs/Myc feedback loop. Cancer Lett. 2019, 450, 63–75. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, A.; Ho, T.-T.; Zhang, Z.; Zhou, N.; Ding, X.; Zhang, X.; Xu, M.; Mo, Y.-Y. Linc-RoR promotes c-Myc expression through hnRNP I and AUF1. Nucleic Acids Res. 2016, 44, 3059–3069. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cai, Q.; Li, W.; Feng, F.; Yang, L. Long non-coding RNA EPIC1 promotes cholangiocarcinoma cell growth. Biochem. Biophys. Res. Commun. 2018, 504, 654–659. [Google Scholar] [CrossRef]
- Zeng, C.; Liu, S.; Lu, S.; Yu, X.; Lai, J.; Wu, Y.; Chen, S.; Wang, L.; Yu, Z.; Luo, G.; et al. The c-Myc-regulated lncRNA NEAT1 and paraspeckles modulate imatinib-induced apoptosis in CML cells. Mol. Cancer 2018, 17, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.; Spizzo, R.; Atlasi, Y.; Nicoloso, M.; Shimizu, M.; Redis, R.S.; Nishida, N.; Gafà, R.; Song, J.; Guo, Z.; et al. CCAT2, a novel noncoding RNA mapping to 8q24, underlies metastatic progression and chromosomal instability in colon cancer. Genome Res. 2013, 23, 1446–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, Y.Y.; Moriarity, B.S.; Gong, W.; Akiyama, R.; Tiwari, A.; Kawakami, H.; Ronning, P.; Reuland, B.; Guenther, K.; Beadnell, T.C.; et al. PVT1 dependence in cancer with MYC copy-number increase. Nature 2014, 512, 82–86. [Google Scholar] [CrossRef]
- Lv, Y.; Yang, X.; Wang, L. Effect of lncRNA THOR on proliferation and igration of colon cancer cells. Oncol. Lett. 2019, 18, 2518–2522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Wei, J.; Zhao, C.; Xiang, S.; Shi, M.; Wang, Y. Targeting LncRNA EPIC1 to inhibit human colon cancer cell progression. Aging 2020, 12. [Google Scholar] [CrossRef]
- Zhou, J.; Li, X.; Wu, M.; Lin, C.; Guo, Y.; Tian, B. Knockdown of Long Noncoding RNA GHET1 Inhibits Cell Proliferation and Invasion of Colorectal Cancer. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2016, 23, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Yan, T.; Bao, Y.; Shen, C.; Yu, C.; Zhu, X.; Tian, X.; Guo, F.; Liang, Q.; Liu, Q.; et al. LncRNA GLCC1 promotes colorectal carcinogenesis and glucose metabolism by stabilizing c-Myc. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, J.-H.; Wu, Q.-N.; Jin, Y.; Wang, D.-S.; Chen, Y.-X.; Liu, J.; Luo, X.-J.; Meng, Q.; Pu, H.-Y.; et al. LncRNA LINRIS stabilizes IGF2BP2 and promotes the aerobic glycolysis in colorectal cancer. Mol. Cancer 2019, 18, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Hu, H.; Yuan, Z.; Zhang, Q.; Xiong, H.; Hu, Z.; Wu, H.; Huang, R.; Wang, G.; Tang, Q. LncRNA NEAT1 remodels chromatin to promote the 5-Fu resistance by maintaining colorectal cancer stemness. Cell Death Dis. 2020, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Gu, Y.; Cheng, X.; Jiang, H.; Huang, Y.; Zhang, Y.; Yu, G.; Cheng, Y.; Zhou, L. Upregulation lnc-NEAT1 contributes to colorectal cancer progression through sponging miR-486-5p and activating NR4A1/Wnt/β-catenin pathway. Cancer Biomark. 2021, 30, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Zhou, R.; Meng, Y.; Duan, R.; Wu, L.; Li, R.; Deng, F.; Lin, C.; Zhao, L. Long noncoding RNA CMPK2 promotes colorectal cancer progression by activating the FUBP3–c-Myc axis. Oncogene 2020, 39, 3926–3938. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wei, D.; Yang, C.; Sun, H.; Lu, T.; Kuang, D. Overexpression of long noncoding RNA, NEAT1 promotes cell proliferation, invasion and migration in endometrial endometrioid adenocarcinoma. Biomed. Pharmacother. 2016, 84, 244–251. [Google Scholar] [CrossRef]
- Yang, F.; Xue, X.; Bi, J.; Zheng, L.; Zhi, K.; Gu, Y.; Fang, G. Long noncoding RNA CCAT1, which could be activated by c-Myc, promotes the progression of gastric carcinoma. J. Cancer Res. Clin. Oncol. 2012, 139, 437–445. [Google Scholar] [CrossRef]
- Zhang, X.; Bo, P.; Liu, L.; Zhang, X.; Li, J. Overexpression of long non-coding RNA GHET1 promotes the development of multidrug resistance in gastric cancer cells. Biomed. Pharmacother. 2017, 92, 580–585. [Google Scholar] [CrossRef]
- Sur, S.; Nakanishi, H.; Steele, R.; Ray, R.B. Depletion of PCAT-1 in head and neck cancer cells inhibits tumor growth and induces apoptosis by modulating c-Myc-AKT1-p38 MAPK signalling pathways. BMC Cancer 2019, 19, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, B.; Yang, B.; Wang, Q.; Zheng, X.; Guo, Y.; Lu, W. lncRNA PVT1 promotes hepatitis B virus-positive liver cancer progression by disturbing histone methylation on the c-Myc promoter. Oncol. Rep. 2019, 43, 718–726. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, D.; Li, Y.; Yan, S.; Dang, H.; Yue, H.; Ling, J.; Chen, F.; Zhao, Y.; Gou, L.; et al. Long noncoding RNA CCAT2 promotes hepatocellular carcinoma proliferation and metastasis through up-regulation of NDRG1. Exp. Cell Res. 2019, 379, 19–29. [Google Scholar] [CrossRef]
- Guo, X.; Hua, Y. CCAT1: An oncogenic long noncoding RNA in human cancers. J. Cancer Res. Clin. Oncol. 2016, 143, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Lu, H.-Y.; Xia, Y.-H.; Jiang, A.-G.; Lv, Y.-X. Long non-coding RNA EPIC1 promotes human lung cancer cell growth. Biochem. Biophys. Res. Commun. 2018, 503, 1342–1348. [Google Scholar] [CrossRef]
- Yang, Y.-R.; Zang, S.-Z.; Zhong, C.-L.; Li, Y.-X.; Zhao, S.-S.; Feng, X.-J. Increased expression of the lncRNA PVT1 promotes tumorigenesis in non-small cell lung cancer. Int. J. Clin. Exp. Pathol. 2014, 7, 6929–6935. [Google Scholar]
- Olivero, C.E.; Martínez-Terroba, E.; Zimmer, J.; Liao, C.; Tesfaye, E.; Hooshdaran, N.; Schofield, J.; Bendor, J.; Fang, D.; Simon, M.D.; et al. p53 Activates the Long Noncoding RNA Pvt1b to Inhibit Myc and Suppress Tumorigenesis. Mol. Cell 2020, 77, 761–774.e8. [Google Scholar] [CrossRef]
- Hosono, Y.; Niknafs, Y.S.; Prensner, J.R.; Iyer, M.K.; Dhanasekaran, S.M.; Mehra, R.; Pitchiaya, S.; Tien, J.; Escara-Wilke, J.; Poliakov, A.; et al. Oncogenic Role of THOR, a Conserved Cancer/Testis Long Non-coding RNA. Cell 2017, 171, 1559–1572.e20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.-S.; Lv, Y.; Xu, X.-C.; Zuo, Y.; Song, Y.; Wu, G.-P.; Lu, P.-H.; Zhang, Z.-Q.; Chen, M.-B. Triptonide inhibits human nasopharyngeal carcinoma cell growth via disrupting Lnc-RNA THOR-IGF2BP1 signaling. Cancer Lett. 2019, 443, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, L.; He, X.; Zhang, J.; Zhu, Z.; Zhang, M.; Li, X. lncRNA CCAT2 promotes radiotherapy resistance for human esophageal carcinoma cells via the miR-145/p70S6K1 and p53 pathway. Int. J. Oncol. 2019, 56, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Sahebi, R.; Malakootian, M.; Balalaee, B.; Shahryari, A.; Khoshnia, M.; Abbaszadegan, M.R.; Moradi, A.; Mowla, S.J. Linc-ROR and its spliced variants 2 and 4 are significantly up-regulated in esophageal squamous cell carcinoma. Iran. J. Basic Med. Sci 2016, 19, 1131–1135. [Google Scholar]
- Yan, L.; Wu, X.; Yin, X.; Du, F.; Liu, Y.; Ding, X. LncRNA CCAT2 promoted osteosarcoma cell proliferation and invasion. J. Cell. Mol. Med. 2018, 22, 2592–2599. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Chen, M.; Xu, Y.; Chen, X.; Zhou, P.; Zhao, X.; Pang, F.; Liang, W. Long non-coding RNA THOR promotes human osteosarcoma cell growth in vitro and in vivo. Biochem. Biophys. Res. Commun. 2018, 499, 913–919. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, M.; Wang, Z.; Guo, W.; Yang, D. MYC-binding lncRNA EPIC1 promotes AKT-mTORC1 signaling and rapamycin resistance in breast and ovarian cancer. Mol. Carcinog. 2020, 59, 1188–1198. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhou, S.; Guo, B. Vitamin D Suppresses Ovarian Cancer Growth and Invasion by Targeting Long Non-Coding RNA CCAT2. Int. J. Mol. Sci. 2020, 21, 2334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napoli, S.; Piccinelli, V.; Mapelli, S.N.; Pisignano, G.; Catapano, C.V. Natural antisense transcripts drive a regulatory cascade controlling c-MYC transcription. RNA Biol. 2017, 14, 1742–1755. [Google Scholar] [CrossRef]
- Hung, C.-L.; Wang, L.-Y.; Yu, Y.-L.; Chen, H.-W.; Srivastava, S.; Petrovics, G.; Kung, H.-J. A long noncoding RNA connects c-Myc to tumor metabolism. Proc. Natl. Acad. Sci. USA 2014, 111, 18697–18702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Z.-D.; Han, L.; Lee, H.; Zhuang, L.; Zhang, Y.; Baddour, J.; Nagrath, D.; Wood, C.G.; Gu, J.; Wu, X.; et al. Energy stress-induced lncRNA FILNC1 represses c-Myc-mediated energy metabolism and inhibits renal tumor development. Nat. Commun. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- Ye, X.-T.; Huang, H.; Huang, W.-P.; Hu, W.-L. LncRNA THOR promotes human renal cell carcinoma cell growth. Biochem. Biophys. Res. Commun. 2018, 501, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y. LncRNA THOR acts as a retinoblastoma promoter through enhancing the combination of c-myc mRNA and IGF2BP1 protein. Biomed. Pharmacother. 2018, 106, 1243–1249. [Google Scholar] [CrossRef]
- Kozłowska, J.; Kozioł, K.; Stasiak, M.; Obacz, J.; Guglas, K.; Poter, P.; Mackiewicz, A.; Kolenda, T. The role of NEAT1 lncRNA in squamous cell carcinoma of the head and neck is still difficult to define. Współczesna Onkologia 2020, 24, 96–105. [Google Scholar] [CrossRef]
- Łaźniak, S.; Lutkowska, A.; Wareńczak-Florczak, Ż.; Sowińska, A.; Tsibulski, A.; Roszak, A.; Sajdak, S.; Jagodziński, P.P. The association of CCAT2 rs6983267 SNP with MYC expression and progression of uterine cervical cancer in the Polish population. Arch. Gynecol. Obstet. 2018, 297, 1285–1292. [Google Scholar] [CrossRef] [Green Version]
- Fox, A.H.; Lamond, A. Paraspeckles. Cold Spring Harb. Perspect. Biol. 2010, 2, a000687. [Google Scholar] [CrossRef]
- Yang, F.; Xue, X.; Zheng, L.; Bi, J.; Zhou, Y.; Zhi, K.; Gu, Y.; Fang, G. Long non-coding RNA GHET1 promotes gastric carcinoma cell proliferation by increasing c-Myc mRNA stability. FEBS J. 2014, 281, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; You, L.; Ren, X.; Zhao, W.; Liao, Q.; Zhao, Y. Long non-coding RNA PVT1 and cancer. Biochem. Biophys. Res. Commun. 2016, 471, 10–14. [Google Scholar] [CrossRef]
- Xie, J.; Wang, X.; Proud, C.G. mTOR inhibitors in cancer therapy. F1000Research 2016, 5, 2078. [Google Scholar] [CrossRef] [Green Version]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, E.N. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2012, 19, 141–157. [Google Scholar] [CrossRef] [Green Version]
- Pamudurti, N.R.; Bartok, O.; Jens, M.; Ashwal-Fluss, R.; Stottmeister, C.; Ruhe, L.; Hanan, M.; Wyler, E.; Perez-Hernandez, D.; Ramberger, E.; et al. Translation of CircRNAs. Mol. Cell 2017, 66, 9–21.e7. [Google Scholar] [CrossRef] [Green Version]
- Ghafouri-Fard, S.; Omrani, M.D.; Taheri, M. Long noncoding RNA PVT1: A highly dysregulated gene in malignancy. J. Cell. Physiol. 2020, 235, 818–835. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Chen, Y.; Fu, M.; Zang, X.; Cang, M.; Niu, Y.; Zhang, W.; Zhang, Y.; Mao, Z.; Shao, M.; et al. Circular RNA CCDC66 promotes gastric cancer progression by regulating c-Myc and TGF-β signaling pathways. J. Cancer 2020, 11, 2759–2768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Zhang, H.; Tao, D.; Xie, F.; Liu, F.; Gu, C.; Wang, M.; Wang, L.; Jiang, G.; Wang, Z.; et al. CircCDYL inhibits the expression of C-MYC to suppress cell growth and migration in bladder cancer. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1349–1356. [Google Scholar] [CrossRef] [Green Version]
- Xie, F.; Xiao, X.; Tao, D.; Huang, C.; Wang, L.; Liu, F.; Zhang, H.; Niu, H.; Jiang, G. circNR3C1 Suppresses Bladder Cancer Progression through Acting as an Endogenous Blocker of BRD4/C-myc Complex. Mol. Ther. Nucleic Acids 2020, 22, 510–519. [Google Scholar] [CrossRef]
- Yang, Q.; Du, W.W.; Wu, N.; Yang, W.; Awan, F.M.; Fang, L.; Ma, J.; Li, X.; Zeng, Y.; Yang, Z.; et al. A circular RNA promotes tumorigenesis by inducing c-myc nuclear translocation. Cell Death Differ. 2017, 24, 1609–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, W.; Liao, X.; Wang, Y.; Li, L.; Li, J.; Chen, Z.; Tian, T.; He, J. circCTIC1 promotes the self-renewal of colon TICs through BPTF-dependent c-Myc expression. Carcinogenesis 2019, 40, 560–568. [Google Scholar] [CrossRef]
- Cai, J.; Chen, Z.; Wang, J.; Wang, J.; Chen, X.; Liang, L.; Huang, M.; Zhang, Z.; Zuo, X. circHECTD1 facilitates glutaminolysis to promote gastric cancer progression by targeting miR-1256 and activating β-catenin/c-Myc signaling. Cell Death Dis. 2019, 10, 1–15. [Google Scholar] [CrossRef]
- Yang, Y.; Gao, X.; Zhang, M.; Yan, S.; Sun, C.; Xiao, F.; Huang, N.; Yang, X.; Zhao, K.; Zhou, H.; et al. Novel Role of FBXW7 Circular RNA in Repressing Glioma Tumorigenesis. J. Natl. Cancer Inst. 2018, 110, 304–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, S.; Yao, T.; Xu, Y.; Zhang, D.; Fan, S.; Ma, J. CircECE1 activates energy metabolism in osteosarcoma by stabilizing c-Myc. Mol. Cancer 2020, 19, 1–17. [Google Scholar] [CrossRef]
- Wang, M.; Chen, B.; Ru, Z.; Cong, L. CircRNA circ-ITCH suppresses papillary thyroid cancer progression through miR-22-3p/CBL/β-catenin pathway. Biochem. Biophys. Res. Commun. 2018, 504, 283–288. [Google Scholar] [CrossRef]
- Bi, W.; Huang, J.; Nie, C.; Liu, B.; He, G.; Han, J.; Pang, R.; Ding, Z.; Xu, J.; Zhang, J. CircRNA circRNA_102171 promotes papillary thyroid cancer progression through modulating CTNNBIP1-dependent activation of β-catenin pathway. J. Exp. Clin. Cancer Res. 2018, 37, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, C.V. MYC on the Path to Cancer. Cell 2012, 149, 22–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalkat, M.; De Melo, J.; Hickman, K.A.; Lourenco, C.; Redel, C.; Resetca, D.; Tamachi, A.; Tu, W.B.; Penn, L.Z. MYC Deregulation in Primary Human Cancers. Genes 2017, 8, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truica, M.I.; Burns, M.C.; Han, H.; Abdulkadir, S.A. Turning Up the Heat on MYC: Progress in Small-Molecule Inhibitors. Cancer Res. 2021, 81, 248–253. [Google Scholar] [CrossRef]
- Nair, S.K.; Burley, S.K. X-Ray Structures of Myc-Max and Mad-Max Recognizing DNA: Molecular Bases of Regulation by Proto-Oncogenic Transcription Factors. Cell 2003, 112, 193–205. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Jain, A.D.; Truica, M.I.; Izquierdo-Ferrer, J.; Anker, J.; Lysy, B.; Sagar, V.; Luan, Y.; Chalmers, Z.R.; Unno, K.; et al. Small-Molecule MYC Inhibitors Suppress Tumor Growth and Enhance Immunotherapy. Cancer Cell 2019, 36, 483–497.e15. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Shi, K.; Qu, Y.; Chu, B.; Qian, Z. Engineering Nanoparticles for Targeted Delivery of Nucleic Acid Therapeutics in Tumor. Mol. Ther. Methods Clin. Dev. 2019, 12, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Trivedi, P.; Jain, N.K. Advances in siRNA delivery in cancer therapy. Artif. Cells Nanomed. Biotechnol. 2018, 46, 274–283. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer | MiRNA | Alteration in Cancer | Mechanistically {Association with the Altered Level of this RNA in Tumor Cells} | References |
|---|---|---|---|---|
| Acute myeloid leukemia | let-7 | Down-regulated | {Poor prognosis} | [130] |
| miR-155 | Down-regulated | Inhibits cancer cell proliferation | [149] | |
| Bladder cancer | miR-147 | Down-regulated | Inhibits cancer cell proliferation | [150] |
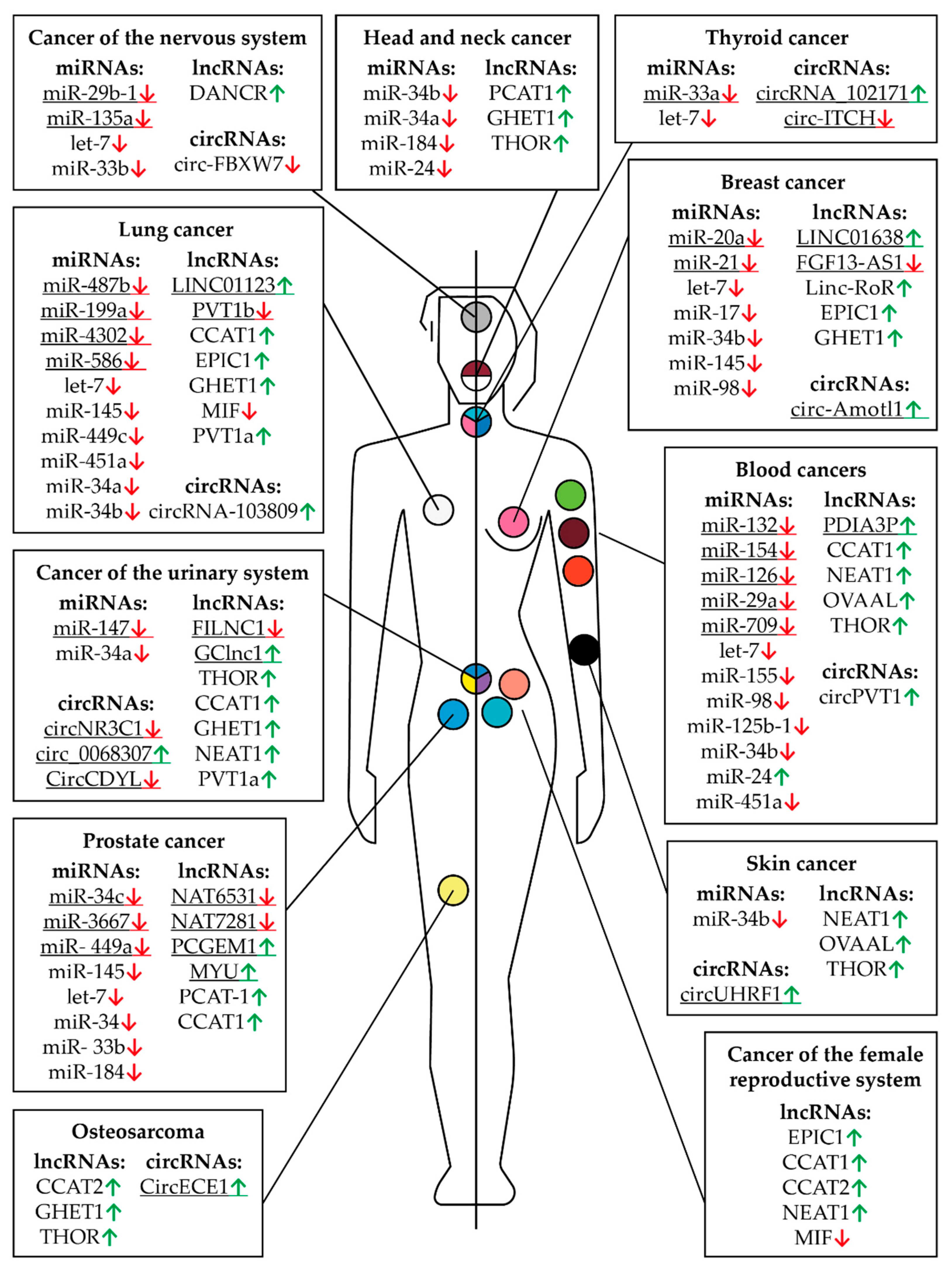
| Burkitt lymphoma | let-7a let-7b miR-98 | Down-regulated | Reverses MYC-induced growth | [120] |
| miR-34b-5p let-7c | Down-regulated | G1 arrest | [151] | |
| let-7-5p miR-132-5p miR-125b-1 miR-154 | Down-regulated | Inhibits cancer cell proliferation | [121] | |
| Breast cancer | miR-17-5p miR-20a-5p | Down-regulated | Inhibits cancer cell proliferation | [152] |
| miR-34b-3p | Down-regulated | {Associated with metastasis development} | [153] | |
| miR-145-5p | Down-regulated | {Poor prognosis} | [154] | |
| miR-21-5p miR-98-5p let-7 | Down-regulated | Suppresses cancer cell growth | [136] | |
| let-7 | Down-regulated | {Poor prognosis} | [131] | |
| let-7 | Down-regulated | Inhibits cancer cells proliferation | [137,138] | |
| Colon cancer | miR-34b-3p | Down-regulated | {Associated with metastasis development} | [153] |
| miR-145-5p | Down-regulated | {Poor prognosis} | [154] | |
| let-7 | Down-regulated | Suppresses cancer cell growth | [142,143] | |
| miR-33b miR-93 | Down-regulated | Inhibited cell proliferation, migration, and invasion | [155] | |
| Colorectal cancer | miR-320b | Down-regulated | Inhibits cancer cell proliferation | [156] |
| miR-182-5p | Down-regulated | Inhibits cancer cell proliferation | [156] | |
| miR-182a-5p | Down-regulated | Inhibits cancer cell proliferation | [157] | |
| miR-200b-3p | Down-regulated | Inhibits cancer cell proliferation | [158] | |
| Diffuse large B-cell lymphoma | miR-34b-5p | Down-regulated | Inhibits cancer cell proliferation | [159] |
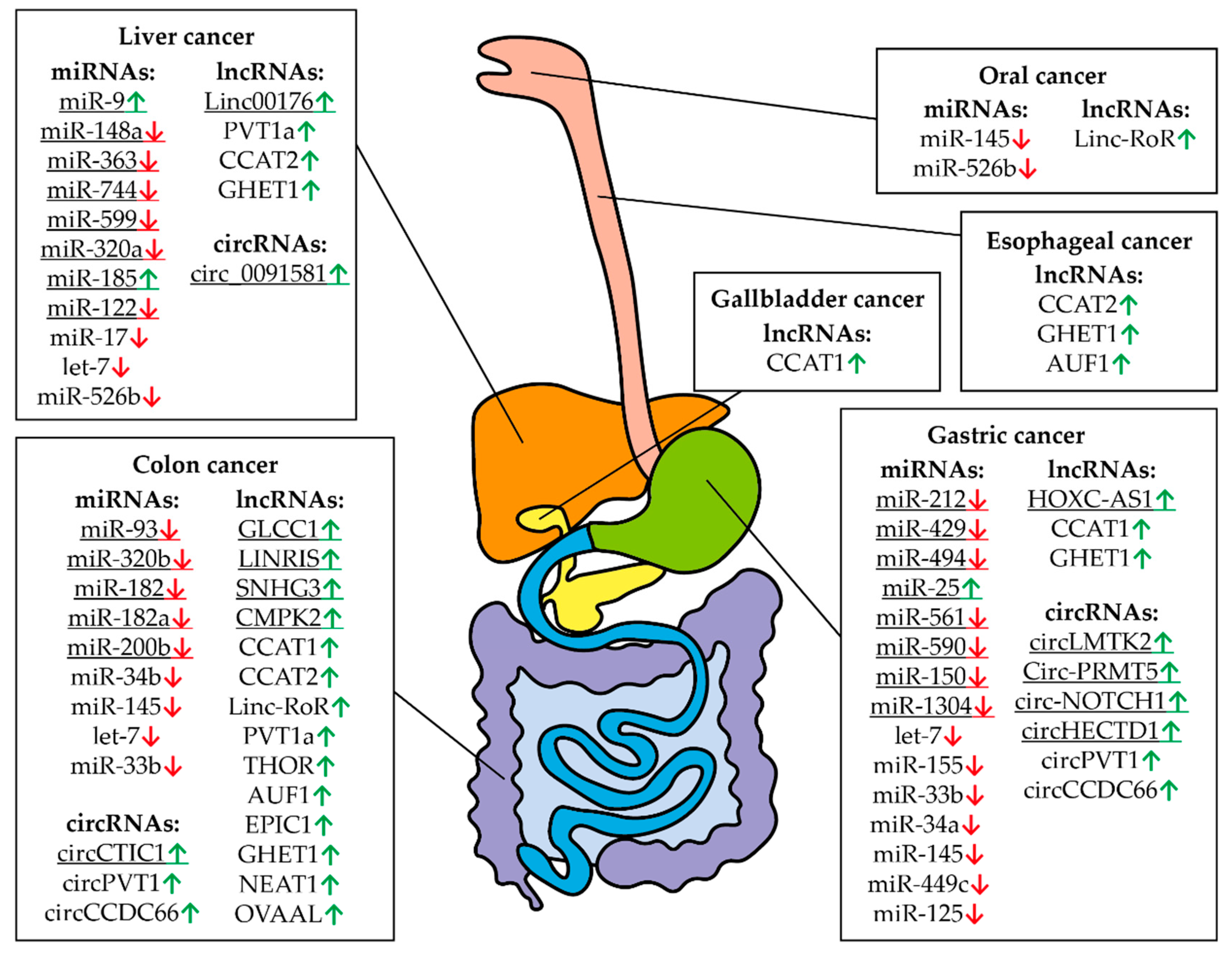
| Gastric cancer | miR-212-3p | Down-regulated | Inhibits cancer cell proliferation | [160] |
| miR-429 | Down-regulated | Inhibits cancer cell viability, proliferation, and attachment | [123] | |
| let-7 | Down-regulated | {Poor prognosis} | [132] | |
| miR-494-3p | Down-regulated | Inhibits cancer cells proliferation | [145] | |
| miR-155-5p | Down-regulated | Inhibits cancer cell growth and invasion | [144] | |
| miR-33b-5p | Down-regulated | Inhibited cell proliferation, migration, and invasion | [161] | |
| miR-25-5p | Up-regulated | Inhibits cancer cell apoptosis | [162] | |
| miR-34a | Down-regulated | Inhibits cancer cell growth and invasion | [163] | |
| miR-561-3p | Down-regulated | Inhibits cancer cell growth and invasion | [164] | |
| miR-590-3p | Down-regulated | Inhibits cancer cell proliferation | [165] | |
| miR-150-5p | Down-regulated | Inhibits cell proliferation and migration | [166] | |
| miR-145 miR-1304 | Down-regulated | Inhibits cell proliferation | [167] | |
| miR-449c-5p | Down-regulated | Inhibits cell proliferation and migration | [168] | |
| miR-125 | Down-regulated | Inhibits cell proliferation | [169] | |
| Glioma | miR-29b-1 | Down-regulated | Inhibits cancer cell proliferation | [170] |
| miR-33b-5p | Down-regulated | Inhibits cancer cell proliferation | [171] | |
| miR-135a-5p | Down-regulated | Inhibits cancer cell proliferation | [172] | |
| Head and neck carcinoma | miR-34b-3p | Down-regulated | {Associated with metastasis development} | [153] |
| miR-34a-5p | Down-regulated | Attenuates tumor growth and metastasis | [173] | |
| Hodgkin Lymphoma | miR-24-3p | Up-regulated | Protects cancer cells from apoptosis | [124] |
| Liver cancer | let-7g | Down-regulated | Inhibits proliferation of hepatocellular carcinoma cells | [139] |
| let-7 | Down-regulated | {Poor prognosis} | [133] | |
| miR-148a-5p miR-363-3p | Down-regulated | G1 arrest | [174] | |
| miR-744-5p | Down-regulated | Inhibits cancer cell proliferation | [175] | |
| Liver cancer | miR-599 | Down-regulated | Inhibits cancer cell proliferation, migration, and invasion | [176] |
| miR-320a | Down-regulated | Inhibits tumor proliferation and invasion | [177] | |
| let-7 | Down-regulated | Inhibits cancer cell proliferation | [140] | |
| miR-9 miR-185-5p | Up-regulated | Inhibits cancer cell proliferation and survival | [178] | |
| miR-17-5p | Down-regulated | Represses invasiveness and metastasis, increases survival | [127] | |
| miR-122-5p | Down-regulated | Inhibits cancer cell proliferation | [179] | |
| miR-526b | Down-regulated | Inhibits cancer cell proliferation | [180] | |
| Lung cancer | miR-34b-3p | Down-regulated | {Associated with metastasis development} | [153] |
| let-7a-5p | Down-regulated | Inhibits the growth of lung cancer | [141] | |
| miR-145-5p | Down-regulated | Inhibits cancer cell proliferation | [181] | |
| miR-487b-3p | Down-regulated | Inhibits cancer cell growth and invasion | [182] | |
| miR-449c-5p | Down-regulated | Inhibits cancer cell proliferation | [183] | |
| miR-451a | Down-regulated | Reverses EMT to mesenchymal–epithelial transition | [146] | |
| miR-34a-5p | Down-regulated | Inhibits cancer cell proliferation | [184] | |
| miR-199a-5p | Down-regulated | Inhibits cancer cell proliferation | [185] | |
| miR-4302 | Down-regulated | Inhibits cancer cell proliferation and invasion | [88] | |
| miR-586-5p | Up-regulated | Enhances cancer cell proliferation | [59] | |
| Medulloblastoma | miR-33b-5p | Down-regulated | Inhibits cancer cell proliferation | [186] |
| Melanomas | miR-34b-3p | Down-regulated | {Associated with metastasis development} | [153] |
| Myeloma | miR-126-5p | Down-regulated | Inhibits cancer cell proliferation | [187] |
| miR-29a-3p | Down-regulated | Inhibits cancer cell viability | ||
| Nasopharyngeal carcinoma | miR-184 | Down-regulated | Blocks cell growth and survival | [188] |
| miR-24-3p | Down-regulated | Suppresses metastasis | [125] | |
| Neuroblastoma | let-7 | Down-regulated | {Worse overall survival} | [134] |
| Oral squamous cell carcinoma | miR-145-5p | Down-regulated | Inhibits cancer cell proliferation | [189,190] |
| miR-526b-5p | Down-regulated | Inhibits cancer cell proliferation | [191] | |
| Prostate cancer | miR-34c-5p | Down-regulated | Inhibits cancer cell proliferation | [192] |
| miR-145-5p | Down-regulated | Inhibits cancer cell proliferation | [193] | |
| let-7 | Down-regulated | Higher in non-metastatic tumors | [135] | |
| miR-34 let-7 | Down-regulated | {Poor prognosis} | [194] | |
| miR-3667-3p | Down-regulated | Inhibits cancer cell proliferation | [195] | |
| miR- 449a | Down-regulated | Enhances cancer cell radiosensitivity | [196] | |
| miR- 33b | Down-regulated | Inhibits cancer cell proliferation | [197] | |
| miR-184 | Down-regulated | Inhibits cancer cell proliferation | [198] | |
| Renal cell carcinoma | miR-34a-5p | Down-regulated | Suppresses malignant transformation | [199] |
| T-cell acute lymphoblastic leukemia | miR-451a miR-709 | Down-regulated | Inhibits cancer cell proliferation | [200] |
| Thyroid cancer | let-7f-5p | Down-regulated | Inhibits cancer cell proliferation | [201] |
| miR-33a-5p | Down-regulated | Inhibits cancer cell proliferation | [202] |
| Cancer | lncRNA | Alteration in Cancer | Mechanistically {Association with the Altered Level of this RNA in Tumor Cells} | References |
|---|---|---|---|---|
| Acute myeloid leukemia | CCAT1 | Up-regulated | Promotes cancer cell proliferation and survival | [149] |
| Bladder cancer | GClnc1 | Up-regulated | Promotes cancer cell proliferation, metastasis, and invasiveness | [217] |
| GHET1 | Up-regulated | {Predicts an unfavorable survival} | [218] | |
| NEAT1 | Up-regulated | Promotes cancer cell proliferation, invasion, and survival | [219] | |
| PVT1a | Up-regulated | Promotes cancer cell proliferation and invasion | [220] | |
| Breast cancer | EPIC1 | Up-regulated | Promotes cancer cell proliferation and survival | [221] |
| GHET1 | Up-regulated | {Predicts an unfavorable survival} | [218] | |
| FGF13-AS1 | Down-regulated | Suppresses cancer cell proliferation, migration, and invasion | [222] | |
| LINC01638 | Up-regulated | {Predicts a poor outcome} | [97] | |
| Linc-RoR | Up-regulated | Suppresses cancer cell proliferation | [223] | |
| Cervical cancer | MIF | Down-regulated | Suppresses cancer cell proliferation | [59] |
| Cholangiocarcinoma | EPIC1 | Up-regulated | Promotes cancer cell proliferation | [224] |
| Chronic myeloid leukemia | NEAT1 | Up-regulated | Promotes cancer cell proliferation and survival | [225] |
| Colon cancer | CCAT1 | Up-regulated | Promotes cancer cell proliferation, migration, and invasion | [213] |
| CCAT2 | Up-regulated | Promotes metastatic progression and chromosomal instability in colon cancer | [226] | |
| Linc-RoR | Up-regulated | Promotes cancer cell proliferation | [223] | |
| PVT1a | Up-regulated | Promotes cancer cell proliferation and invasion | [227] | |
| THOR | Up-regulated | Promotes cancer cell proliferation and migration | [228] | |
| AUF1 | Up-regulated | Promotes cancer cell proliferation | [223] | |
| EPIC1 | Up-regulated | Promotes cancer cell proliferation and invasion | [229] | |
| Colorectal cancer | GHET1 | Up-regulated | Promotes cancer cell proliferation | [230] |
| GLCC1 | Up-regulated | Promotes cancer cell survival and proliferation | [231] | |
| LINRIS | Up-regulated | Promotes cancer cell proliferation | [232] | |
| NEAT1 | Up-regulated | Promotes cancer cell proliferation and survival | [233,234] | |
| SNHG3 | Up-regulated | Promotes cancer cell proliferation | [157] | |
| OVAAL | Up-regulated | Promotes cancer cell proliferation | [206] | |
| CMPK2 | Up-regulated | Promotes colorectal cancer progression | [235] | |
| Diffuse large B-cell lymphoma | NEAT1 | Up-regulated | Promotes cancer cell proliferation and survival | [159] |
| Endometrial adenocarcinoma | NEAT1 | Up-regulated | Promotes cancer cell proliferation, invasion, and migration | [236] |
| Gallbladder cancer | CCAT1 | Up-regulated | Promotes cancer cell proliferation and survival | [214] |
| Gastric cancer | CCAT1 | Up-regulated | Promotes cancer cell proliferation, migration, and invasion | [237] |
| GHET1 | Up-regulated | Promotes cancer cell proliferation | [238] | |
| HOXC-AS1 | Up-regulated | Promotes cancer cell proliferation and metastasis | [165] | |
| Glioma | DANCR | Up-regulated | Promotes cancer cell proliferation | [171] |
| Head and neck cancer | GHET1 | Up-regulated | {Predicts an unfavorable survival} | [218] |
| PCAT-1 | Up-regulated | Promotes cancer cell proliferation | [239] | |
| Hepatocellular carcinoma | PVT1a | Up-regulated | Promotes cancer cell proliferation and invasion | [240] |
| Liver cancer | CCAT2 | Up-regulated | Promotes cancer cell proliferation and invasion | [241] |
| GHET1 | Up-regulated | {Predicts an unfavorable survival} | [218] | |
| Linc00176 | Up-regulated | Promotes cancer cell proliferation | [178] | |
| Lung cancer | CCAT1 | Up-regulated | Promotes cancer cell proliferation and survival | [209,242] |
| EPIC1 | Up-regulated | Promotes cancer cell proliferation | [243] | |
| GHET1 | Up-regulated | {Predicts an unfavorable survival} | [218] | |
| LINC01123 | Up-regulated | Promotes cancer cell proliferation | [185] | |
| MIF | Down-regulated | Suppresses cancer cell proliferation | [59] | |
| PVT1a | Up-regulated | Promotes cancer cell proliferation and invasion | [244] | |
| PVT1b | Down-regulated | Suppresses cancer cell proliferation | [245] | |
| Medulloblastoma | DANCR | Up-regulated | Promotes cancer cell proliferation | [171] |
| Melanoma | OVAAL | Up-regulated | Promotes cancer cell proliferation | [206] |
| THOR | Up-regulated | Promotes cancer cell proliferation | [246] | |
| Multiple myeloma | PDIA3P | Up-regulated | Enhances cancer cell proliferation and drug resistance | [56] |
| Nasopharyngeal carcinoma | THOR | Up-regulated | Promotes cancer cell proliferation | [247] |
| Esophageal cancer | CCAT2 | Up-regulated | Promotes radiotherapy resistance | [248] |
| GHET1 | Up-regulated | {Predicts an unfavorable survival} | [218] | |
| AUF1 | Up-regulated | Promotes cancer cell proliferation | [249] | |
| Oral cancer | Linc-RoR | Up-regulated | {Associated with tumor recurrence and poor therapeutic response} | [190] |
| Osteosarcoma | CCAT2 | Up-regulated | Promotes cancer cell proliferation and invasion | [250] |
| GHET1 | Up-regulated | {Predicts an unfavorable survival} | [218] | |
| THOR | Up-regulated | Promotes cancer cell proliferation | [251] | |
| Ovarian cancer | EPIC1 | Up-regulated | Promotes cancer cell proliferation and survival | [252] |
| CCAT1 | Up-regulated | Promotes cancer cell proliferation and survival | [215] | |
| CCAT2 | Up-regulated | Promotes cancer cell proliferation and invasion | [253] | |
| Pancreatic cancer | GHET1 | Up-regulated | {Predicts an unfavorable survival} | [218] |
| GLS-AS | Down-regulated | Suppresses cancer cell proliferation | [207] | |
| Prostate cancer | CCAT1 | Up-regulated | Promotes cancer cell proliferation and survival | [211] |
| NAT6531 NAT7281 | Down-regulated | [254] | ||
| PCAT-1 | Up-regulated | Promotes cancer cell proliferation | [195] | |
| PCGEM1 | Up-regulated | Promotes cancer cell proliferation and survival | [255] | |
| MYU | Up-regulated | Promotes cancer cell proliferation | [198] | |
| Renal cancer | CCAT1 | Up-regulated | Promotes cancer cell proliferation and survival | [212] |
| FILNC1 | Down-regulated | Inhibits tumor development | [256] | |
| Renal cell carcinoma | THOR | Up-regulated | Promotes cancer cell proliferation | [257] |
| Retinoblastoma | THOR | Up-regulated | Promotes cancer cell proliferation | [258] |
| Squamous cell carcinoma | NEAT1 | Up-regulated | {Worse overall survival} | [259] |
| Uterine cervical cancer | CCAT2 | Up-regulated | Progression of uterine cervical cancer | [260] |
| Cancer | circRNA | Alteration in Cancer | Mechanistically | References |
|---|---|---|---|---|
| Bladder cancer | CircCDYL | Down-regulated | Suppresses cell growth and migration | [269] |
| circNR3C1 | Down-regulated | Suppresses cancer cell growth | [270] | |
| circ_0068307 | Up-regulated | Promotes cancer cell migration and proliferation | [150] | |
| Breast cancer | circ-Amotl1 | Up-regulated | Stimulates tumor growth | [271] |
| Colon cancer | circPVT1 | Up-regulated | Promotes cancer cell proliferation | [267] |
| circCTIC1 | Up-regulated | Promotes cancer cell proliferation | [272] | |
| circCCDC66 | Up-regulated | Promotes cancer cell proliferation and migration | [155] | |
| Gastric cancer | circLMTK2 | Up-regulated | Promotes cancer cell proliferation | [166] |
| Circ-PRMT5 | Up-regulated | Promotes cancer cell proliferation | [167] | |
| circ-NOTCH1 | Up-regulated | Promotes cancer cell migration and proliferation | [168] | |
| circPVT1 | Up-regulated | Promotes cancer cell proliferation | [267] | |
| circHECTD1 | Up-regulated | Promotes cancer cell proliferation | [273] | |
| circCCDC66 | Up-regulated | Promotes cancer cell proliferation | [268] | |
| Glioblastoma | circ-FBXW7 | Down-regulated | Suppresses cancer cell growth | [274] |
| Leukemia | circPVT1 | Up-regulated | Promotes cancer cell proliferation | [267] |
| Liver cancer | circ_0091581 | Up-regulated | Stimulates tumor growth | [180] |
| Lung cancer | circRNA_103809 | Up-regulated | Stimulates cancer cell proliferation and invasion | [88] |
| Osteosarcoma | CircECE1 | Up-regulated | Promotes cancer cell migration and proliferation | [275] |
| Squamous cell carcinoma | circUHRF1 | Up-regulated | Promotes cancer cell proliferation | [191] |
| Thyroid cancer | circ-ITCH | Down-regulated | Suppresses cancer cell migration and proliferation | [276] |
| circRNA_102171 | Up-regulated | Promotes cancer cell migration and proliferation | [277] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stasevich, E.M.; Murashko, M.M.; Zinevich, L.S.; Demin, D.E.; Schwartz, A.M. The Role of Non-Coding RNAs in the Regulation of the Proto-Oncogene MYC in Different Types of Cancer. Biomedicines 2021, 9, 921. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9080921
Stasevich EM, Murashko MM, Zinevich LS, Demin DE, Schwartz AM. The Role of Non-Coding RNAs in the Regulation of the Proto-Oncogene MYC in Different Types of Cancer. Biomedicines. 2021; 9(8):921. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9080921
Chicago/Turabian StyleStasevich, Ekaterina Mikhailovna, Matvey Mikhailovich Murashko, Lyudmila Sergeevna Zinevich, Denis Eriksonovich Demin, and Anton Markovich Schwartz. 2021. "The Role of Non-Coding RNAs in the Regulation of the Proto-Oncogene MYC in Different Types of Cancer" Biomedicines 9, no. 8: 921. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9080921