
The Effect of Acute and Chronic Thermotherapy on Type 2 Diabetic Skeletal Muscle Gene Expression and Inflammatory Markers

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Skeletal Muscle Cell-Line Demographics
2.2. Media Preparation
2.3. Skeletal Muscle Cell Maintenance
2.4. Sub-Culturing the Culture Vessel
2.5. Skeletal Muscle Cell Count and Viability
2.6. Thermotherapy Treatments
2.7. RNA Purification
2.8. Gene Expression
2.9. Cytokine Inflammatory Markers
2.10. Statistical Analysis
3. Results
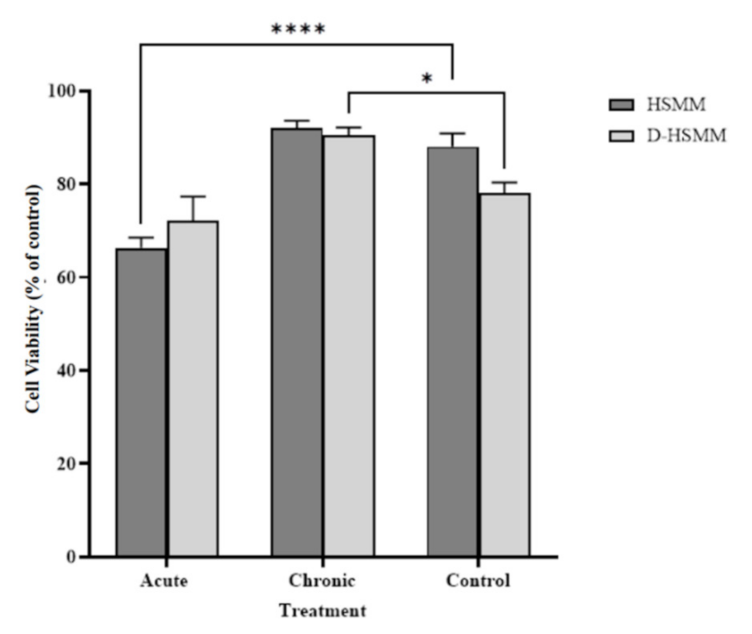
3.1. Skeletal Muscle Cell Viability
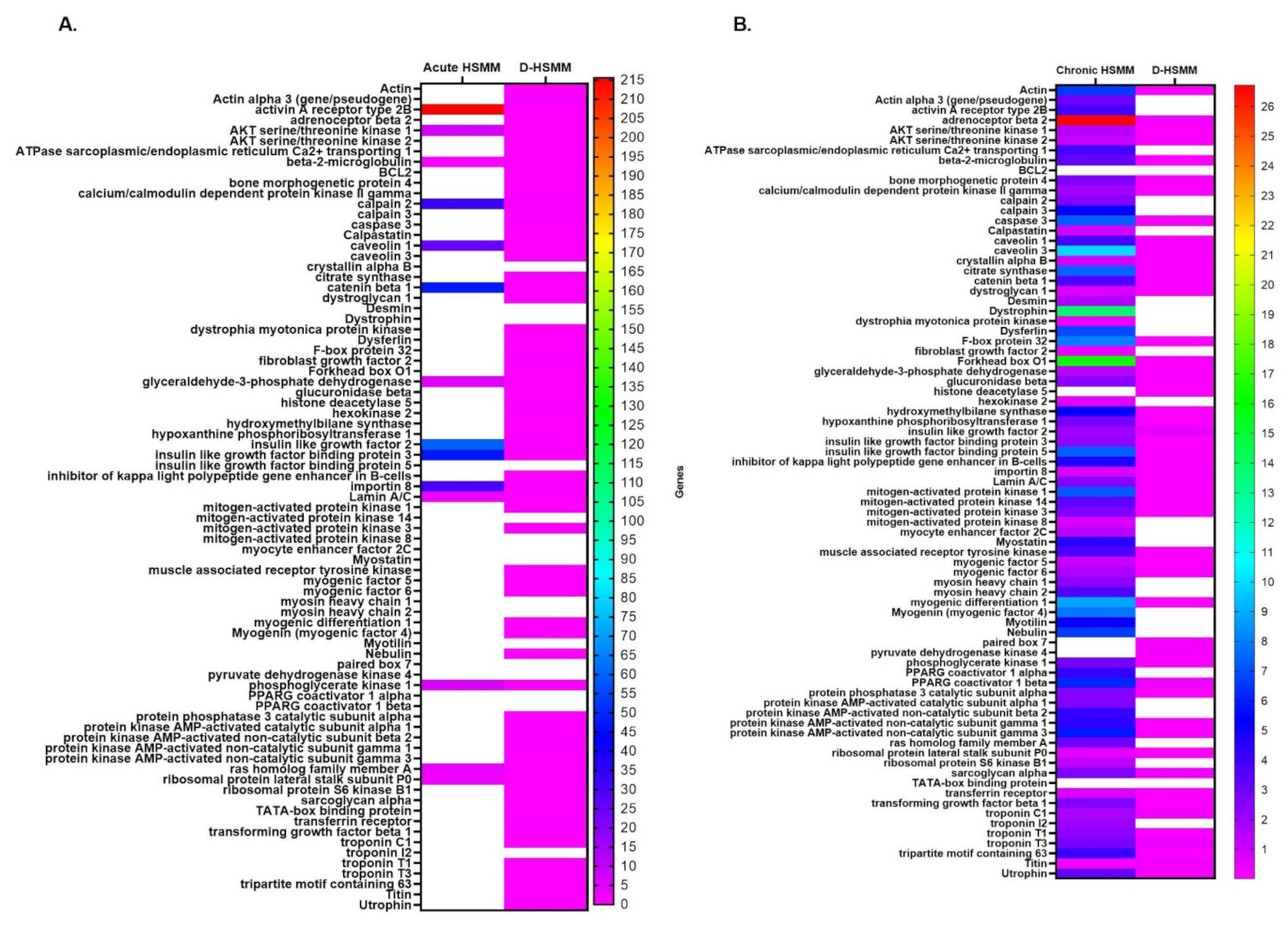
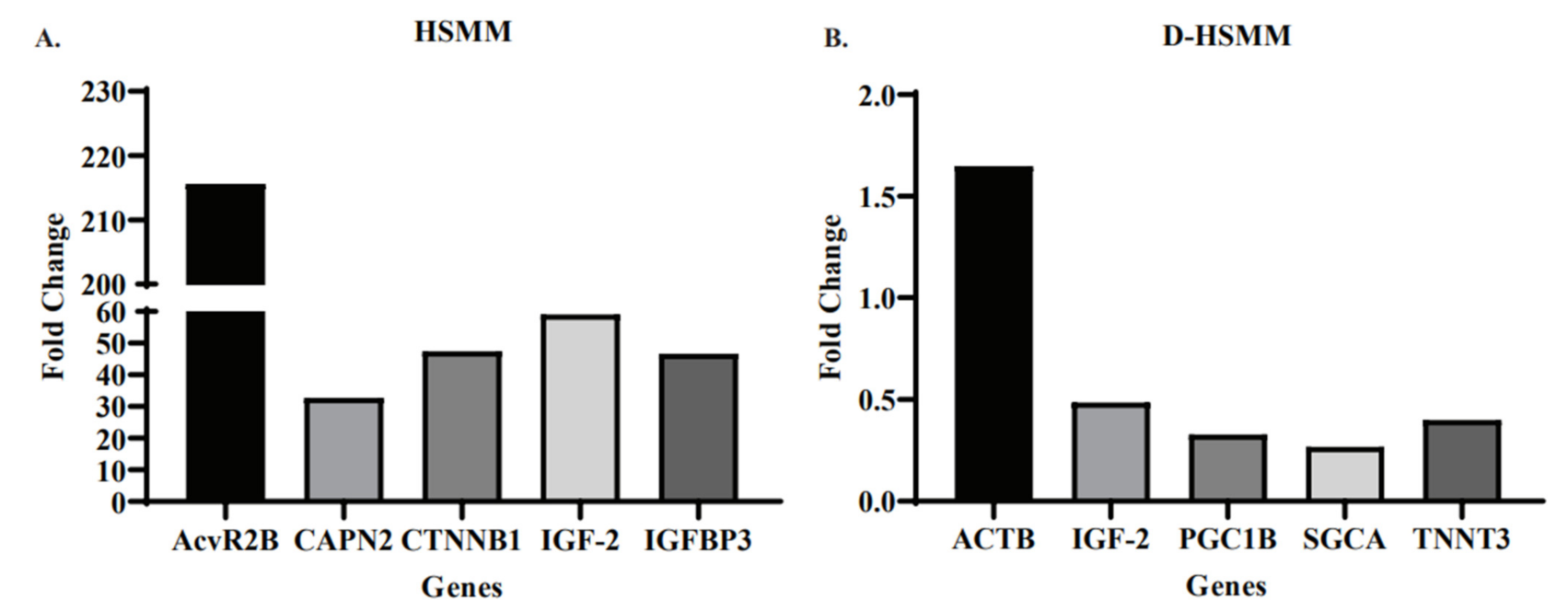
3.2. Gene Expression
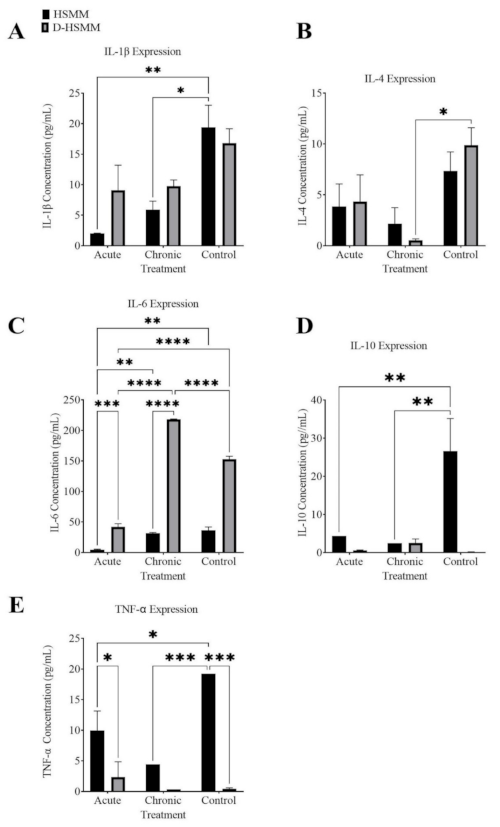
3.3. Cytokine Inflammatory Markers
3.3.1. Interleukin-1β
3.3.2. Interleukin-4
3.3.3. Interleukin-6
3.3.4. Interleukin-10
3.3.5. Tumor Necrosis Factor-α
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- American Diabetes Association. Classification and diagnosis of diabetes: Standards of medical care in diabetes—2018. Diabetes Care 2018, 41, S13–S27. [Google Scholar] [CrossRef] [Green Version]
- Meng, Y.Y.; Pickett, M.C.; Babey, S.H.; Davis, A.C.; Goldstein, H. Diabetes tied to a third of California hospital stays, driving health care costs higher. Policy Brief UCLA Cent. Health Policy Res. 2014, 1–7. [Google Scholar]
- American Diabetes Association. 2. Classification and diagnosis of diabetes: Standards of medical care in diabetes-2021. Diabetes Care 2021, 44, S15–S33. [Google Scholar] [CrossRef]
- Scott, D.; de Courten, B.; Ebeling, P.R. Sarcopenia: A potential cause and consequence of type 2 diabetes in Australia’s ageing population? Med. J. Aust. 2016, 205, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Vienberg, S.; Geiger, J.; Madsen, S.; Dalgaard, L.T. MicroRNAs in metabolism. Acta Physiol. 2017, 219, 346–361. [Google Scholar] [CrossRef] [PubMed]
- Lowell, B.B. Mitochondrial dysfunction and type 2 diabetes. Science 2005, 307, 384–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunt, V.E.; Eymann, T.M.; Francisco, M.A.; Howard, M.J.; Minson, C.T. Passive heat therapy improves cutaneous microvascular function in sedentary humans via improved nitric oxide-dependent dilation. J. Appl. Physiol. 2016, 121, 716–723. [Google Scholar] [CrossRef] [Green Version]
- Périard, J.D.; Racinais, S.; Sawka, M.N. Adaptations and mechanisms of human heat acclimation: Applications for competitive athletes and sports: Adaptations and mechanisms of heat acclimation. Scand. J. Med. Sci. Sports 2015, 25, 20–38. [Google Scholar] [CrossRef] [PubMed]
- Welc, S.S.; Morse, D.A.; Mattingly, A.J.; Laitano, O.; King, M.A.; Clanton, T.L. The impact of hyperthermia on receptor-mediated interleukin-6 regulation in mouse skeletal muscle. PLoS ONE 2016, 11, e0148927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takii, R.; Inouye, S.; Fujimoto, M.; Nakamura, T.; Shinkawa, T.; Prakasam, R.; Tan, K.; Hayashida, N.; Ichikawa, H.; Hai, T.; et al. Heat shock transcription Factor 1 inhibits expression of IL-6 through activating transcription Factor 3. J. Immunol. 2010, 184, 1041–1048. [Google Scholar] [CrossRef] [Green Version]
- Hooper, P. Hot-tub therapy for type 2 diabetes mellitus. N. Engl. J. Med. 1999, 341, 917–925. [Google Scholar] [CrossRef] [Green Version]
- Counting Cells Using a Hemocytometer n.d. Available online: https://www.abcam.com/protocols/counting-cells-using-a-haemocytometer (accessed on 27 February 2019).
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kelley, D.E.; Mandarino, L.J. Fuel selection in human skeletal muscle in insulin resistance: A reexamination. Diabetes 2000, 49, 677–683. [Google Scholar] [CrossRef] [Green Version]
- Chomentowski, P.; Coen, P.M.; Radiková, Z.; Goodpaster, B.H.; Toledo, F.G.S. Skeletal muscle mitochondria in insulin resistance: Differences in intermyofibrillar versus subsarcolemmal subpopulations and relationship to metabolic flexibility. J. Clin. Endocrinol. Metab. 2011, 96, 494–503. [Google Scholar] [CrossRef]
- Nielsen, J.; Mogensen, M.; Vind, B.F.; Sahlin, K.; Højlund, K.; Schrøder, H.D.; Ørtenblad, N. Increased subsarcolemmal lipids in type 2 diabetes: Effect of training on localization of lipids, mitochondria, and glycogen in sedentary human skeletal muscle. Am. J. Physiol.-Endocrinol. Metab. 2010, 298, E706–E713. [Google Scholar] [CrossRef] [Green Version]
- Goto, K.; Kojima, A.; Kobayashi, T.; Uehara, K.; Morioka, S.; Naito, T.; Akema, T.; Sugiura, T.; Ohira, Y.; Yoshioka, T. Heat stress as a countermeasure for prevention of muscle atrophy in microgravity environment. Jpn. J. Aerosp. Environ. Med. 2005, 42, 51–59. [Google Scholar]
- Uehara, K.; Goto, K.; Kobayashi, T.; Kojima, A.; Akema, T.; Sugiura, T.; Yamada, S.; Ohira, Y.; Yoshioka, T.; Aoki, H. Heat-Stress Enhances Proliferative Potential in Rat Soleus Muscle. Jpn. J. Physiol. 2004, 54, 263–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, K.; Okuyama, R.; Sugiyama, H.; Honda, M.; Kobayashi, T.; Uehara, K.; Akema, T.; Sugiura, T.; Yamada, S.; Ohira, Y.; et al. Effects of heat stress and mechanical stretch on protein expression in cultured skeletal muscle cells. Pflügers Arch. Eur. J. Physiol. 2003, 447, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Goto, K.; Ohno, Y.; Goto, A.; Ikuta, A.; Suzuki, M.; Ohira, T.; Tsuchiya, N.; Nishizawa, S.; Koya, T.; Egawa, T.; et al. Some aspects of heat stress on the plasticity of skeletal muscle cells. J. Phys. Fit. Sports Med. 2012, 1, 197–204. [Google Scholar] [CrossRef]
- Goto, K.; Oda, H.; Morioka, S.; Naito, T.; Akema, T.; Kato, H.; Fujiya, H.; Nakajima, Y.; Sugiura, T.; Ohira, Y.; et al. Skeletal muscle hypertrophy induced by low-intensity exercise with heat-stress in healthy human subjects. Jpn. J. Aerosp. Environ. Med. 2007, 44, 13–16. [Google Scholar]
- Goto, K.; Oda, H.; Kondo, H.; Igaki, M.; Suzuki, A.; Tsuchiya, S.; Murase, T.; Hase, T.; Fujiya, H.; Matsumoto, I.; et al. Responses of muscle mass, strength and gene transcripts to long-term heat stress in healthy human subjects. Eur. J. Appl. Physiol. 2011, 111, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Yamashita-Goto, K.; Ohira, Y.; Okuyama, R.; Sugiyama, H.; Honda, M.; Sugiura, T.; Yamada, S.; Akema, T.; Yoshioka, T. Heat stress facilitates stretch-induced hypertrophy of cultured muscle cells. J. Gravit. Physiol. 2002, 9, P145–P146. [Google Scholar]
- Latres, E.; Amini, A.R.; Amini, A.A.; Griffiths, J.; Martin, F.J.; Wei, Y.; Lin, H.C.; Yancopoulos, G.D.; Glass, D.J. Insulin-like growth Factor-1 (IGF-1) inversely regulates atrophy-induced genes via the Phosphatidylinositol 3-Kinase/Akt/Mammalian target of Rapamycin (PI3K/Akt/mTOR) pathway. J. Biol. Chem. 2005, 280, 2737–2744. [Google Scholar] [CrossRef] [Green Version]
- Hulmi, J.J.; Hentilä, J.; DeRuisseau, K.C.; Oliveira, B.M.; Papaioannou, K.G.; Autio, R.; Kujala, U.M.; Ritvos, O.; Kainulainen, H.; Korkmaz, A.; et al. Effects of muscular dystrophy, exercise and blocking activin receptor IIB ligands on the unfolded protein response and oxidative stress. Free. Radic. Biol. Med. 2016, 99, 308–322. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.L.; Walton, K.L.; Winbanks, C.E.; Murphy, K.T.; Thomson, R.E.; Makanji, Y.; Qian, H.; Lynch, G.S.; Harrison, C.A.; Gregorevic, P. Elevated expression of activins promotes muscle wasting and cachexia. FASEB J. 2013, 28, 1711–1723. [Google Scholar] [CrossRef]
- IGF2 Gene. U.S. National Library of Medicine (Bethesda, MD, USA), 2020. Available online: https://ghr.nlm.nih.gov/gene/IGF2 (accessed on 17 September 2021).
- Armstrong, D.D.; Esser, K.A. Wnt/β-catenin signaling activates growth-control genes during overload-induced skeletal muscle hypertrophy. Am. J. Physiol.-Cell Physiol. 2005, 289, C853–C859. [Google Scholar] [CrossRef] [PubMed]
- Novak, A.; Dedhar, S. Signaling through β-catenin and Lef/Tcf. Cell. Mol. Life Sci. 1999, 56, 523–537. [Google Scholar] [CrossRef]
- Regulated Subset of G1 Growth-Control Genes in Response to Derepression by the Wnt Pathway|PNAS n.d. Available online: https://www.pnas.org/content/100/6/3245.short (accessed on 2 June 2020).
- Xin, H.; Zhang, X.; Sun, D.; Zhang, C.; Hao, Y.; Gu, X. Chronic heat stress increases insulin-like growth factor-1(IGF-1) but does not affect IGF-binding proteins in growing pigs. J. Therm. Biol. 2018, 77, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, K.K.W. The calpain family and human disease. Trends Mol. Med. 2001, 7, 355–362. [Google Scholar] [CrossRef]
- Garach-Jehoshua, O.; Ravid, A.; Liberman, U.A.; Reichrath, J.; Glaser, T.; Koren, R. Upregulation of the calcium-dependent protease, calpain, during keratinocyte differentiation. Br. J. Dermatol. 1998, 139, 950–957. [Google Scholar] [CrossRef]
- ACTB Gene. U.S. National Library of Medicine, 2020. Available online: https://ghr.nlm.nih.gov/gene/ACTB (accessed on 17 September 2021).
- Joseph, R.; Srivastava, O.P.; Pfister, R.R. Downregulation of β-actin gene and human antigen R in human keratoconus. Investig. Opthalmology Vis. Sci. 2012, 53, 4032–4041. [Google Scholar] [CrossRef] [Green Version]
- Bunnell, T.M.; Burbach, B.J.; Shimizu, Y.; Ervasti, J.M. β-Actin specifically controls cell growth, migration, and the G-actin pool. Mol. Biol. Cell 2011, 22, 4047–4058. [Google Scholar] [CrossRef]
- TNNT3 Gene. Gene Cards: The Hman Gene Database n.d. Available online: https://www.genecards.org/cgi-bin/carddisp.pl?gene=TNNT3 (accessed on 14 March 2020).
- Pinto, J.R.; Muller-Delp, J.; Chase, P.B. Will you still need me (Ca2+, TnT, and DHPR), will you still cleave me (calpain), when I’m 64? Aging Cell 2017, 16, 202–204. [Google Scholar] [CrossRef]
- Delbono, O.; Xia, J.; Treves, S.; Wang, Z.-M.; Jimenez-Moreno, R.; Payne, A.M.; Messi, M.L.; Briguet, A.; Schaerer, F.; Nishi, M.; et al. Loss of skeletal muscle strength by ablation of the sarcoplasmic reticulum protein JP45. Proc. Natl. Acad. Sci. USA 2007, 104, 20108–20113. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.L.; Foletta, V.C.; Wright, C.R.; Sepulveda, P.V.; Konstantopoulos, N.; Sanigorski, A.; Della Gatta, P.; Cameron-Smith, D.; Kralli, A.; Russell, A. PGC-1α and PGC-1β increase protein synthesis via ERRα in C2C12 myotubes. Front. Physiol. 2018, 9, 1336. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. SGCA Sarcoglycan Alpha; National Center for Biotechnology Information: Bethesda, MD, USA, 2020. [Google Scholar]
- Welc, S.S.; Clanton, T.L. The regulation of interleukin-6 implicates skeletal muscle as an integrative stress sensor and endocrine organ. Exp. Physiol. 2013, 98, 359–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, J.G.; Fielding, R.A.; Fiatarone, M.A.; Orencole, S.F.; Dinarello, C.A.; Evans, W.J. Increased interleukin 1 beta in human skeletal muscle after exercise. Am. J. Physiol. Integr. Comp. Physiol. 1989, 257, R451–R455. [Google Scholar] [CrossRef]
- Song, M.; Pinsky, M.R.; Kellum, J.A. Heat shock factor 1 inhibits nuclear factor–κB nuclear binding activity during endotoxin tolerance and heat shock. J. Crit. Care 2008, 23, 406–415. [Google Scholar] [CrossRef]
- Welc, S.S.; Clanton, T.L.; Dineen, S.M.; Leon, L.R. Heat stroke activates a stress-induced cytokine response in skeletal muscle. J. Appl. Physiol. 2013, 115, 1126–1137. [Google Scholar] [CrossRef]
- Welc, S.S.; Phillips, N.A.; Oca-Cossio, J.; Wallet, S.M.; Chen, D.L.; Clanton, T.L. Hyperthermia increases interleukin-6 in mouse skeletal muscle. Am. J. Physiol. Physiol. 2012, 303, C455–C466. [Google Scholar] [CrossRef] [PubMed]
- Robins, H.I.; Kutz, M.; Wiedemann, G.J.; Katschinski, D.M.; Paul, D.; Grosen, E.; Tiggelaar, C.L.; Spriggs, D.; Gillis, W.; d’Oleire, F. Cytokine induction by 41.8 °C whole body hyperthermia. Cancer Lett. 1995, 97, 195–201. [Google Scholar] [CrossRef]
- Bouchama, A.; Al Mohanna, F.; El-Sayed, R.; Eldali, A.; Saussereau, E.; Chollet-Martin, S.; Ollivier, V.; de Prost, D.; Roberts, G. Experimental heatstroke in baboon: Analysis of the systemic inflammatory response. Shock 2005, 24, 332–335. [Google Scholar] [CrossRef] [PubMed]
- Leon, L.R.; Blaha, M.D.; DuBose, D.A. Time course of cytokine, corticosterone, and tissue injury responses in mice during heat strain recovery. J. Appl. Physiol. 2006, 100, 1400–1409. [Google Scholar] [CrossRef] [PubMed]
- Idriss, H.T.; Naismith, J.H. TNF α and the TNF receptor superfamily: Structure-function relationship(s). Microsc. Res. Tech. 2000, 50, 184–195. [Google Scholar] [CrossRef]
- Imao, M.; Nagaki, M.; Moriwaki, H. Dual effects of heat stress on tumor necrosis factor- α -induced hepatocyte apoptosis in mice. Lab. Investig. 2006, 86, 959–967. [Google Scholar] [CrossRef] [Green Version]
- Ferat-Osorio, E.; Sánchez-Anaya, A.; Gutiérrez-Mendoza, M.; Boscó-Gárate, I.; Wong-Baeza, I.; Pastelin-Palacios, R.; Pedraza-Alva, G.; Bonifaz, L.C.; Cortés-Reynosa, P.; Pérez-Salazar, E.; et al. Heat shock protein 70 down-regulates the production of toll-like receptor-induced pro-inflammatory cytokines by a heat shock factor-1/constitutive heat shock element-binding factor-dependent mechanism. J. Inflamm. 2014, 11, 19. [Google Scholar] [CrossRef] [Green Version]
- Neto, J.C.R.; Lira, F.S.; Zanchi, N.E.; Oyama, L.M.; Pimentel, G.D.; Santos, R.V.; Seelaender, M.; Nascimento, C.M.O.D. Acute exhaustive exercise regulates IL-2, IL-4 and MyoD in skeletal muscle but not adipose tissue in rats. Lipids Health Dis. 2011, 10, 97. [Google Scholar] [CrossRef] [Green Version]
- Luzina, I.G.; Keegan, A.D.; Heller, N.M.; Rook, G.A.W.; Shea-Donohue, T.; Atamas, S.P. Regulation of inflammation by interleukin-4: A review of “alternatives”. J. Leukoc. Biol. 2012, 92, 753–764. [Google Scholar] [CrossRef] [Green Version]
- Horsley, V.; Jansen, K.M.; Mills, S.T.; Pavlath, G.K. IL-4 Acts as a myoblast recruitment factor during Mammalian muscle growth. Cell 2003, 113, 483–494. [Google Scholar] [CrossRef] [Green Version]
- Luo, G.; Hershko, D.D.; Robb, B.W.; Wray, C.J.; Hasselgren, P.-O. IL-1beta stimulates IL-6 production in cultured skeletal muscle cells through activation of MAP kinase signaling pathway and NF-κ B. Am. J. Physiol. Integr. Comp. Physiol. 2003, 284, R1249–R1254. [Google Scholar] [CrossRef]
- Papanicolaou, D.A.; Wilder, R.L.; Manolagas, S.C.; Chrousos, G.P. The pathophysiologic roles of interleukin-6 in human disease. Ann. Intern. Med. 1998, 128, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Kamimura, D.; Hirano, T. Pleiotropy and specificity: Insights from the interleukin 6 family of cytokines. Immunity 2019, 50, 812–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Wu, Y.; Zhang, Y.; Jin, L.; Luo, L.; Xue, B.; Lu, C.; Zhang, X.; Yin, Z. Hsp70 inhibits lipopolysaccharide-induced NF-κB activation by interacting with TRAF6 and inhibiting its ubiquitination. FEBS Lett. 2006, 580, 3145–3152. [Google Scholar] [CrossRef] [Green Version]
- Park, P.-H.; McMullen, M.R.; Huang, H.; Thakur, V.; Nagy, L.E. Short-term treatment of RAW264.7 macrophages with adiponectin increases tumor necrosis factor-alpha (TNF-α) expression via ERK1/2 activation and Egr-1 expression: Role of TNF-alpha in adiponectin-stimulated interleukin-10 production. J. Biol. Chem. 2007, 282, 21695–21703. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bachnak, L.; Sparks, J.; Newmire, D.E.; Gonzales, X.F.; Omoruyi, F.O. The Effect of Acute and Chronic Thermotherapy on Type 2 Diabetic Skeletal Muscle Gene Expression and Inflammatory Markers. Biomedicines 2021, 9, 1276. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9091276
Bachnak L, Sparks J, Newmire DE, Gonzales XF, Omoruyi FO. The Effect of Acute and Chronic Thermotherapy on Type 2 Diabetic Skeletal Muscle Gene Expression and Inflammatory Markers. Biomedicines. 2021; 9(9):1276. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9091276
Chicago/Turabian StyleBachnak, Louay, Jean Sparks, Daniel E. Newmire, Xavier F. Gonzales, and Felix O. Omoruyi. 2021. "The Effect of Acute and Chronic Thermotherapy on Type 2 Diabetic Skeletal Muscle Gene Expression and Inflammatory Markers" Biomedicines 9, no. 9: 1276. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9091276