
Potential Efficacy of Bacillus coagulans BACO-17 to Modulate Gut Microbiota in Rats Fed High-Fat Diet

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal and Experimental Design
2.2. Feed Efficiency
2.3. Determination of Total Visceral Fat
2.4. Total Liver Lipids Analysis
2.5. Determination of Fecal Moisture and Total Fecal Lipids
2.6. Analysis of Gut Microbiota
2.7. Determination of SCFAs
2.8. Statistical Analysis
3. Results and Discussion
3.1. Body Weight Gain, Food Intake, and Feed Efficiency
3.2. Total Visceral Fat and Liver Total Lipids
3.3. Changes in Fecal Parameters
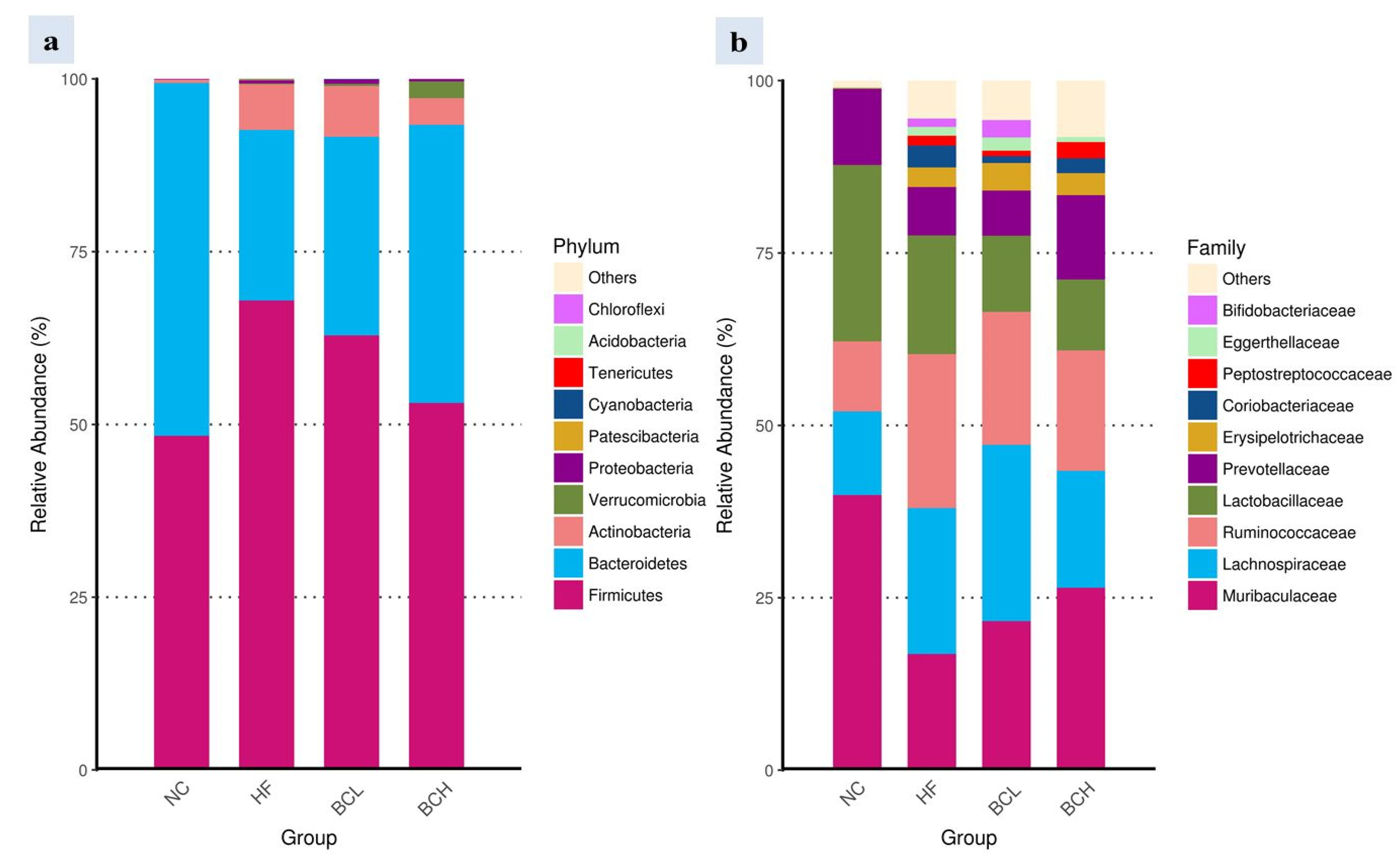
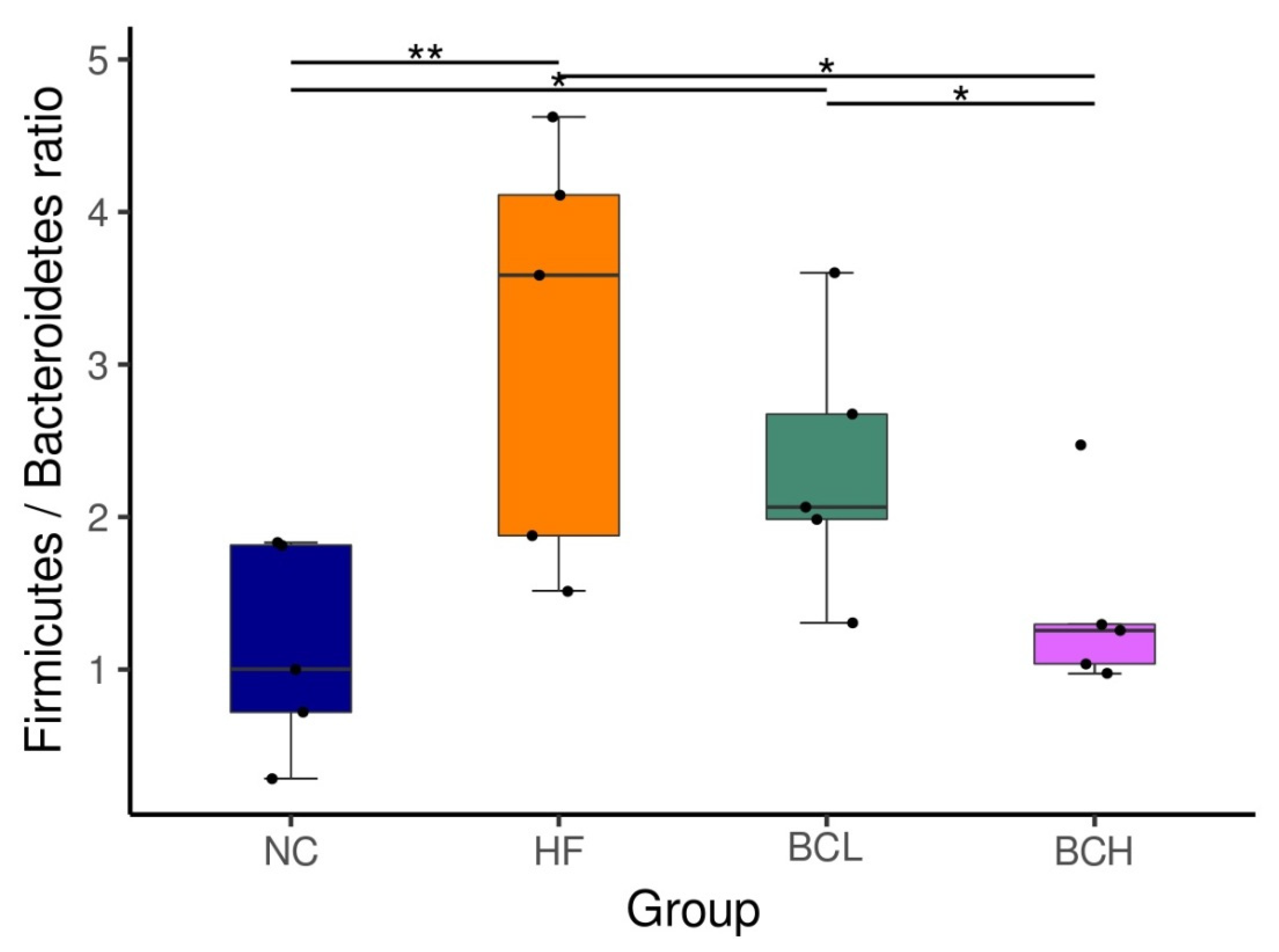
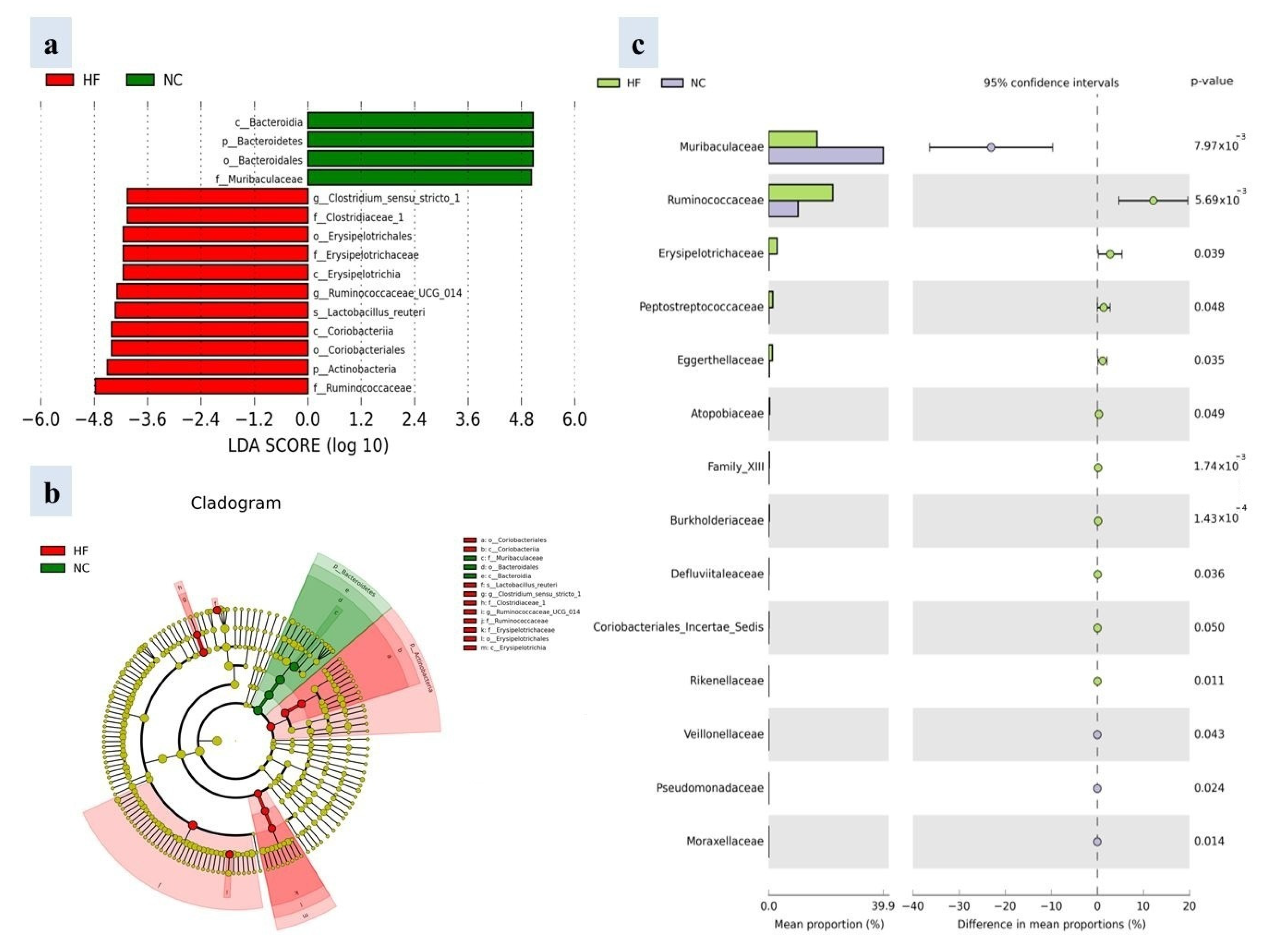
3.4. Profile of Gut Microbiota
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 8 December 2022).
- Apovian, C.M.; Aronne, L.J.; Bessesen, D.H.; McDonnell, M.E.; Murad, M.H.; Pagotto, U.; Ryan, D.H.; Still, C.D. Pharmacological management of obesity: An endocrine society clinical practice guideline. J. Clin. Endocrinol. Metab. 2015, 100, 342–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, N.N.; Wu, T.Y.; Chau, C.F. Natural Dietary and Herbal Products in Anti-Obesity Treatment. Molecules 2016, 21, 1351. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Long, X.; Yang, J.; Du, L.; Zhang, X.; Li, J.; Hou, C. Pomegranate peel polyphenols reduce chronic low-grade inflammatory responses by modulating gut microbiota and decreasing colonic tissue damage in rats fed a high-fat diet. Food Funct. 2019, 10, 8273–8285. [Google Scholar] [CrossRef]
- Liu, H.Y.; Walden, T.B.; Ahl, D.; Nyman, M.; Bertilsson, S.; Phillipson, M.; Holm, L. High-fat diet enriched with bilberry modifies colonic mucus dynamics and restores marked alterations of gut microbiome in rats. Mol. Nutr. Food Res. 2019, 63, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Carding, S.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L.J. Dysbiosis of the gut microbiota in disease. Microb. Ecol. Health Dis. 2015, 26, 1–9. [Google Scholar] [CrossRef]
- Gerritsen, J.; Smidt, H.; Rijkers, G.T.; de Vos, W.M. Intestinal microbiota in human health and disease: The impact of probiotics. Genes Nutr. 2011, 6, 209–240. [Google Scholar] [CrossRef] [Green Version]
- Azad, M.A.K.; Sarker, M.; Li, T.; Yin, J. Probiotic species in the modulation of gut microbiota: An overview. BioMed Res. Int. 2018, 2018, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Konuray, G.; Erginkaya, Z. Potential use of Bacillus coagulans in the food industry. Foods 2018, 7, 92. [Google Scholar] [CrossRef] [Green Version]
- Saw, C.Y.; Chang, T.J.; Chen, P.Y.; Dai, F.J.; Lau, Y.Q.; Chen, T.Y.; Chau, C.F. Presence of Bacillus coagulans spores and vegetative cells in rat intestine and feces and their physiological effects. Biosci. Biotechno. Biochem. 2019, 83, 2327–2333. [Google Scholar] [CrossRef]
- He, J.; Zhang, P.; Shen, L.; Niu, L.; Tan, Y.; Chen, L.; Zhao, Y.; Bai, L.; Hao, X.; Li, X.; et al. Short-chain fatty acids and their association with signaling pathways in inflammation, glucose and lipid metabolism. Int. J. Mol. Sci. 2020, 21, 6356. [Google Scholar] [CrossRef]
- Haldar, L.; Gandhi, D.N. Effect of oral administration of Bacillus coagulans B37 and Bacillus pumilus B9 strains on fecal coliforms, Lactobacillus and Bacillus spp. in rat animal model. Vet. World 2016, 9, 766–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abhari, K.; Saadati, S.; Yari, Z.; Hosseini, H.; Hedayati, M.; Abhari, S.; Alavian, S.M.; Hekmatdoost, A. The effects of Bacillus coagulans supplementation in patients with non-alcoholic fatty liver disease: A randomized, placebo-controlled, clinical trial. Clin. Nutr. ESPEN 2020, 39, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Sudha, M.R.; Jayanthi, N.; Aasin, M.; Dhanashri, R.D.; Anirudh, T. Efficacy of Bacillus coagulans Unique IS2 in treatment of irritable bowel syndrome in children: A double blind, randomised placebo controlled study. Benef. Microbes 2018, 9, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Yu, Z.; Liu, W.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Probiotic characteristics of Bacillus coagulans and associated implications for human health and diseases. J. Funct. Foods 2020, 64, 1–11. [Google Scholar] [CrossRef]
- Campana, R.; Van Hemert, S.; Baffone, W. Strain-specific probiotic properties of lactic acid bacteria and their interference with human intestinal pathogens invasion. Gut Pathog. 2017, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kao, W.M.; Chang, C.R.; Chang, T.J.; Li, S.Y.; Chen, W.J.; Chau, C.F. Inclusion of fructooligosaccharide and resistant maltodextrin in high fat diets promotes simultaneous improvements on body fat reduction and fecal parameters. Molecules 2018, 23, 2169. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, C.E.; You, F.N.; Dai, F.J.; Tung, Y.C.; Chen, W.J.; Chu, H.F.; Wu, S.H.; Chau, C.F. Bacillus coagulans BC198 and Lactobacillus paracasei S38 in combination reduce body fat accumulation and modulate gut microbiota. CyTA–J. Food 2020, 18, 764–775. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.Z.; Sun, T.C.; Huang, Y.W.; Wu, Y.C.; Chen, W.J.; Chu, H.F.; Liu, C.Y.; Chau, C.F. Bacillus coagulans BACO-17 alone or in combination with galacto-oligosaccharide ameliorates Salmonella-induced diarrhea and intestinal inflammation. Processes 2022, 10, 2123. [Google Scholar] [CrossRef]
- Macdiarmid, J.I.; Blundell, J.E. Dietary under-reporting: What people say about recording their food intake. Eur. J. Clin. Nutr. 1997, 51, 199–200. [Google Scholar] [CrossRef]
- Yadav, H.; Lee, J.H.; Lloyd, J.; Walter, P.; Rane, S.G. Beneficial metabolic effects of a probiotic via butyrate-induced GLP-1 hormone secretion. J. Biol. Chem. 2013, 288, 25088–25097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez-Arraño, V.; Martín-Peláez, S. Effects of Probiotics and Synbiotics on Weight Loss in Subjects with Overweight or Obesity: A Systematic Review. Nutrients 2021, 13, 3627. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.T.; Dai, B.; Yan, Y.; Xu, Y.L.; Wang, Y.X.; Yang, C.M. Bacillus licheniformis, a potential probiotic, inhibits obesity by modulating colonic microflora in C57BL/6J mice model. J. Appl. Microbiol. 2019, 127, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.C.; Jung, E.Y.; Kang, D.H.; Kim, J.M.; Chang, U.J.; Suh, H.J. Vitamin C increases the fecal fat excretion by chitosan in guinea-pigs, thereby reducing body weight gain. Phytother. Res. 2010, 24, 1234–1241. [Google Scholar] [CrossRef] [PubMed]
- Jakobsdottir, G.; Xu, J.; Molin, G.; Ahrne, S.; Nyman, M. High-fat diet reduces the formation of butyrate, but increases succinate, inflammation, liver fat and cholesterol in rats, while dietary fibre counteracts these effects. PLoS ONE 2013, 8, e80476. [Google Scholar] [CrossRef] [Green Version]
- Chávez-Carbajal, A.; Nirmalkar, K.; Pérez-Lizaur, A.; Hernández-Quiroz, F.; Ramírez-del-Alto, S.; García-Mena, J.; Hernández-Guerrero, C. Gut microbiota and predicted metabolic pathways in a sample of Mexican women affected by obesity and obesity plus metabolic syndrome. Int. J. Mol. Sci. 2019, 20, 438. [Google Scholar] [CrossRef] [Green Version]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The controversial role of human gut Lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef]
- Aoki, R.; Kamikado, K.; Suda, W.; Takii, H.; Mikami, Y.; Suganuma, N.; Hattori, M.; Koga, Y. A proliferative probiotic Bifidobacterium strain in the gut ameliorates progression of metabolic disorders via microbiota modulation and acetate elevation. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Grider, J.R.; Piland, B.E. The peristaltic reflex induced by short-chain fatty acids is mediated by sequential release of 5-HT and neuronal CGRP but not BDNF. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, 429–437. [Google Scholar] [CrossRef]
- Kimura, I.; Inoue, D.; Maeda, T.; Hara, T.; Ichimura, A.; Miyauchi, S.; Kobayashi, M.; Hirasawa, A.; Tsujimoto, G. Short-chain fatty acids and ketones directly regulate sympathetic nervous system via G protein-coupled receptor 41 (GPR41). Proc. Natl. Acad. Sci. USA 2011, 108, 8030–8035. [Google Scholar] [CrossRef]
- Kimura, I.; Ozawa, K.; Inoue, D.; Imamura, T.; Kimura, K.; Maeda, T.; Terasawa, K.; Kashihara, D.; Hirano, K.; Tani, T.; et al. The gut microbiota suppresses insulin-mediated fat accumulation via the short-chain fatty acid receptor GPR43. Nat. Commun. 2013, 4, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The influence of probiotics on the Firmicutes/Bacteroidetes ratio in the treatment of obesity and inflammatory bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef] [PubMed]
- Million, M.; Maraninchi, M.; Henry, M.; Armougom, F.; Richet, H.; Carrieri, P.; Valero, R.; Raccah, D.; Vialettes, B.; Raoult, D. Obesity-associated gut microbiota is enriched in Lactobacillus reuteri and depleted in Bifidobacterium animalis and Methanobrevibacter smithii. Int. J. Obes. 2012, 36, 817–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geurts, L.; Lazarevic, V.; Derrien, M.; Everard, A.; Van Roye, M.; Knauf, C.; Cani, P.D. Altered gut microbiota and endocannabinoid system tone in obese and diabetic leptin-resistant mice: Impact on apelin regulation in adipose tissue. Front. Microbiol. 2011, 2, 149. [Google Scholar] [CrossRef] [Green Version]
- Lecomte, V.; Kaakoush, N.O.; Maloney, C.A.; Raipuria, M.; Huinao, K.D.; Mitchell, H.M.; Morris, M.J. Changes in gut microbiota in rats fed a high fat diet correlate with obesity-associated metabolic parameters. PLoS ONE 2015, 10, e0126931. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Lyu, W.; Ren, Y.; Li, X.; Zhao, S.; Yang, H.; Xiao, Y. Allobaculum involves in the modulation of intestinal ANGPTLT4 expression in mice treated by high-fat diet. Front. Nutr. 2021, 8, 242. [Google Scholar] [CrossRef]
- Fleissner, C.K.; Huebel, N.; Abd El-Bary, M.M.; Loh, G.; Klaus, S.; Blaut, M. Absence of intestinal microbiota does not protect mice from diet-induced obesity. Br. J. Nutr. 2010, 104, 919–929. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Pan, L.; Cheng, J.; Wang, X.; Zheng, L.; Wang, S.; Wang, H. High-fat-diet-induced gut microbiome changes in mice. Stress Brain 2022, 2, 17. [Google Scholar] [CrossRef]
- Dalby, M.J.; Ross, A.W.; Walker, A.W.; Morgan, P.J. Dietary uncoupling of gut microbiota and energy harvesting from obesity and glucose tolerance in mice. Cell Rep. 2017, 21, 1521–1533. [Google Scholar] [CrossRef] [Green Version]
- Cao, W.; Chin, Y.; Chen, X.; Mi, Y.; Xue, C.; Wang, Y.; Tang, Q. The role of gut microbiota in the resistance to obesity in mice fed a high fat diet. Int. J. Food Sci. Nutr. 2020, 71, 453–463. [Google Scholar] [CrossRef]
- Dao, M.C.; Everard, A.; Aron-Wisnewsky, J.; Sokolovska, N.; Prifti, E.; Verger, E.O.; Kayser, B.; Levenez, F.; Chilloux, J.; Hoyles, L.; et al. Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: Relationship with gut microbiome richness and ecology. Gut 2016, 65, 426–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NC | HF | BCL | BCH | |
|---|---|---|---|---|
| Body weight gain (g) | 191.1 ± 14.0 d | 321.1 ± 29.0 a | 277.8 ± 23.1 b | 245.1 ± 25.0 c |
| Total food intake (g) | 2122.1 ± 56.9 a | 2034.8 ± 143.4 a | 1986.8 ± 185.7 ab | 1817.1 ± 119.5 b |
| Total fat intake (g) | 106.1 ± 2.8 c | 457.8 ± 32.3 a | 447.0 ± 41.8 ab | 408.8 ± 26.9 b |
| Total calorie intake (kcal) | 7215.3 ± 193.5 c | 8749.8 ± 616.5 a | 8543.2 ± 798.4 ab | 7813.4 ± 514.0 bc |
| Feed efficiency (%) | 9.0 ± 0.6 c | 15.8 ± 0.9 a | 14.0 ± 1.0 b | 13.5 ± 0.9 b |
| Visceral Fats | NC | HF | BCL | BCH |
|---|---|---|---|---|
| Total visceral fat | 5.3 ± 0.7 c | 12.1 ± 1.5 a | 11.2 ± 1.6 a | 9.2 ± 0.9 b |
| Perirenal fat | 2.3 ± 0.1 c | 5.5 ± 0.7 a | 4.8 ± 0.6 ab | 4.1 ± 0.6 b |
| Mesenteric fat | 1.4 ± 0.3 b | 3.2 ± 0.7 a | 3.0 ± 0.8 a | 2.4 ± 0.4 a |
| Epididymal fat | 1.7 ± 0.3 c | 3.5 ± 0.4 a | 3.4 ± 0.5 a | 2.6 ± 0.4 b |
| Liver total lipids (mg/g) | 156.3 ± 8.7 c | 294.8 ± 50.4 a | 257.5 ± 30.9 ab | 234.0 ± 30.1 b |
| SCFAs | NC | HF | BCL | BCH |
|---|---|---|---|---|
| Total SCFAs (μmol/g) | 266.5 ± 28.9 a | 193.1 ± 37.5 b | 266.3 ± 29.9 a | 274.0 ± 38.6 a |
| Acetic acid (μmol/g) | 186.5 ± 26.2 a | 119.2 ± 41.4 b | 177.2 ± 33.0 a | 182.0 ± 32.2 a |
| Propanoic acid (μmol/g) | 59.0 ± 21.0 | 40.6 ± 16.9 | 43.4 ± 18.6 | 44.0 ± 7.7 |
| Butyric acid (μmol/g) | 21.1 ± 3.9 c | 33.4± 12.8 b | 45.6 ± 6.7 a | 48.0 ± 7.9 a |
| NC | HF | BCL | BCH | |
|---|---|---|---|---|
| ACE index | 257.73 ± 25.75 a | 345.43 ± 21.87 b | 353.30 ± 16.49 b | 272.65 ± 45.43 a |
| Chao1 index | 254.47 ± 25.49 a | 350.19 ± 24.12 b | 354.30 ± 19.33 b | 274.27 ± 45.61 a |
| Shannon index | 3.97 ± 0.97 a | 5.80 ± 0.28 bc | 6.06 ± 0.35 c | 5.09 ± 0.35 b |
| Simpson index | 0.84 ± 0.11 a | 0.96 ± 0.01 b | 0.97 ± 0.02 b | 0.94 ± 0.01 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chan, Z.; Saw, C.-Y.; Wu, Y.-C.; Lau, D.-K.; Chen, W.-J.; Chu, H.-F.; Liu, C.-Y.; Tseng, A.-L.; Chau, C.-F. Potential Efficacy of Bacillus coagulans BACO-17 to Modulate Gut Microbiota in Rats Fed High-Fat Diet. Processes 2022, 10, 2692. https://0-doi-org.brum.beds.ac.uk/10.3390/pr10122692
Chan Z, Saw C-Y, Wu Y-C, Lau D-K, Chen W-J, Chu H-F, Liu C-Y, Tseng A-L, Chau C-F. Potential Efficacy of Bacillus coagulans BACO-17 to Modulate Gut Microbiota in Rats Fed High-Fat Diet. Processes. 2022; 10(12):2692. https://0-doi-org.brum.beds.ac.uk/10.3390/pr10122692
Chicago/Turabian StyleChan, Zu, Chiou-Yeong Saw, Yi-Ching Wu, Der-Kai Lau, Wei-Jen Chen, Hui-Fang Chu, Cheng-Yen Liu, Ai-Lun Tseng, and Chi-Fai Chau. 2022. "Potential Efficacy of Bacillus coagulans BACO-17 to Modulate Gut Microbiota in Rats Fed High-Fat Diet" Processes 10, no. 12: 2692. https://0-doi-org.brum.beds.ac.uk/10.3390/pr10122692