1. Introduction
Microalgae are often considered as a promising alternative for the production of renewable energy, particularly as a potential source of biofuels, in an attempt to replace fossil fuels with renewable alternatives; and/or as a method of capturing carbon dioxide, towards a more sustainable World [
1,
2,
3]. The biofuels obtained from microalgae can be found in liquid form (bioethanol, biodiesel, vegetal oil) or gaseous form (biohydrogen or biosyngas) [
4,
5]. Other potentialities reside in providing proteins and other nutrients for animal feed and human food supplements [
1,
3,
6,
7] as well as other high-value molecules of pharmaceutical interest [
1,
7]. In this way, microalgae biomass and oil constitute a promising renewable feedstock for the emerging chemical technology and biotechnology industries. Therefore, there is a strong interest in developing dynamic models to better understand the microalgae-based processes and to develop control schemes that allow the optimal operation of these processes [
8,
9].
Nonetheless, in the way to continue the exploration of optimal operating conditions, some physiological properties of the microalgae related to their capacity to process light should be considered, using the current understanding of the photosynthetic process, along with our ability to efficiently exploit its properties. To improve culturing systems, the development of mathematical models capable of predicting photosynthetic productivity under dynamic conditions is crucial. Especially important is the adaptation process, due to the light-harvesting capacity of the photon flux, known as photoacclimation. Microalgae cultures can suffer a loss in biomass productivity due to a lack of illumination resulting from the darkening effect in the photobioreactor, or due to photoinhibition in the case of an excessive illumination. However, this loss can be alleviated by regulation of the light irradiance.
Globally speaking, the photosynthetic yield is affected by two key processes, which alter the impact of light conditions and nutrient supply over the culture. Photoinhibition is the first of them, which causes a decrease in the photosynthesis rate due to an excess of light, and later induces damage over some proteins in the photosynthetic apparatus [
10]. Photoacclimation is the second, and can be defined as the process by which microalgae adapt their pigment composition to the photon flux density, also altering the photosynthesis rate.
A pioneer work can be found in [
11,
12,
13], where the photoinhibition process is modeled at the level of the photosynthetic units inside the microalgae. The effect of photoinhibition is described through the photo-oxidative inactivation of these units, led by an excess of photon flux over the one required for photosynthesis, which denatures key proteins in the electron transfer chain [
14,
15]. This concept was used to explain the hysteresis effect found in P-I curves (photosynthesis rate vs. photon flux density), because photosynthetic rates measured with a succession of increasing irradiance levels are often higher than the rates measured with a series of decreasing irradiance.
For the production of microalgae in photobioreactors, the chemostat mode, i.e., equal inlet and outlet flows seems to be more profitable, because it allows prolonged stable operation, possibly at maximum productivity [
16]. Microalgae photobioreactors are complex systems, mainly due to their nonlinear and time-varying behavior. In early literature, a model proposed for outdoor cultures [
17] simulates biomass production, pH, growth rate, oxygen evolution, and carbon dioxide fixation rate.
Microalgae modeling usually involves three basic states, accounting for biomass, substrate internal quota, and a substrate external concentration, respectively.
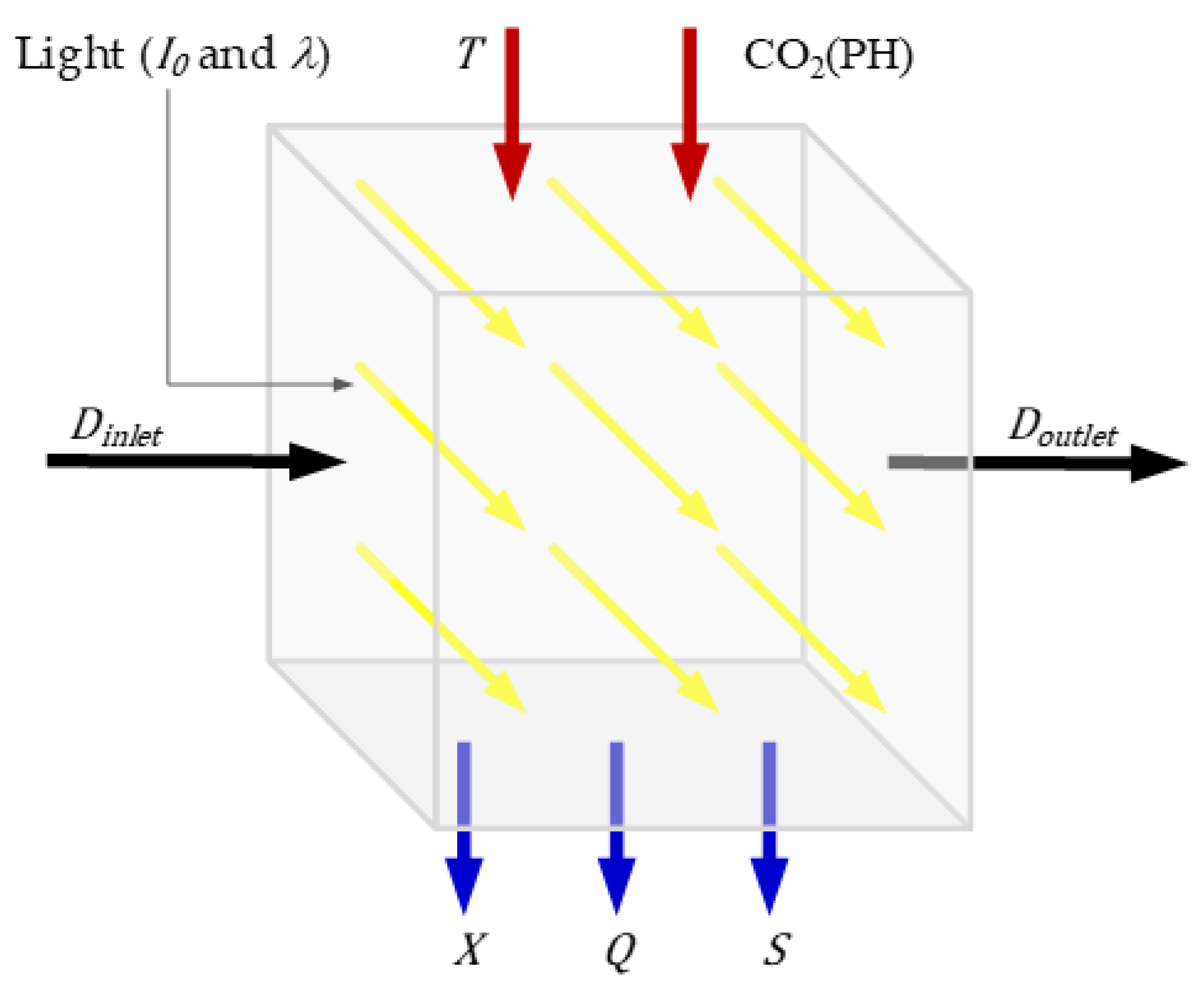
Figure 1 illustrates some of the most common variables involved in a microalgae photobioreactor, either as inputs or outputs. An example of this kind of model can be found in [
18], where a parameter identification is performed over batch cultures of
Dunaliella tertiolecta strain. In addition, modeling of neutral lipids and carbohydrates quotas were introduced in [
19] with a five-state model. A more complex mathematical representation considering eight state variables, in view of nitrogen and phosphorus as substrate limited sources, and comprising sugar and lipids concentration quotas was reported by [
20]. Moreover, this model was used in an optimization problem to minimize the culture costs accounting for light, aeration, and cooling processes [
21]. On the other hand, some research works followed a black-box modeling approach. In [
22] an input-output adaptive identification based on artificial neural networks (ANN) was proposed for biomass control by adjusting the photon flux density (In [
23], it is recommended to use the term
photon flux density instead of
light intensity—widely use in the literature, as it is technically more suitable. Further, it is advised to use the units [
] for this magnitude, instead of [
], as the first one belongs to
Le Système International (SI) of units). In [
24], a Hammerstein–Wiener representation is used to develop an extremum seeking strategy.
One of the most frequently cited models is the one developed in [
25], which accounts for the photoinhibition and shading effects inside the cultures. This model describes the dynamics of the three basic states (
,
, and
), but incorporates a novel approach with a fourth conceptual variable named
photoacclimation photon flux density,
, that represents the photon flux density at which the microalgae are currently physiologically acclimated. This model is presented with an analytical integration based on the average growth rate throughout the whole culture in [
26]. The model of [
25] is in a convenient form to be exploited for process control, particularly when the expression of the photoinhibition phenomenon is simplified and changed for a Haldane-inhibitory formulation. In [
10] an equation is introduced to simulate the dynamics of chlorophyll, removing in this way the intermediate variable of photoacclimation photon flux density to represent the photoacclimation state of the culture. Instead, an expression for the chlorophyll concentration is formulated based on the incident light irradiance. Other modeling studies investigate the effect of other variables that affect microalgae growth, especially temperature [
27].
In this study, we will focus attention on the model originally proposed in [
25,
26] and develop a parameter identification study for cultures of the microalgae
Scenedesmus obliquus in batch photo-bioreactors. Our motivation is that, even if the models of [
25,
26] are largely accepted, their validation with experimental data is limited to a few case studies. In the current work, measurements of the biomass, intracellular substrate quota, extracellular substrate, and Chlorophyll concentrations are collected from nine batch experiments differing by their initial conditions and light exposition conditions. The parameters of a dynamic model are then estimated using a weighted least squares approach, and the uncertainty in the parameters is estimated based on the Fisher Information Matrix (FIM). Monte Carlo studies allow the model cross-validation against datasets not used in the identification procedure. Further, the dynamic model is exploited in a simulation study demonstrating the potential benefits of a Nonlinear Model Predictive Control (NMPC) for the optimization of the biomass production in a continuous photo-bioreactor, acting simultaneously on the dilution rate and the light intensity.
2. Model Description
In this section, we discuss a dynamic model describing photo-acclimation and photo-inhibition, originally proposed in [
25,
26]. This model uses the classical Droop expression [
28] for substrate (
S) uptake, internal quota (
Q) evolution, and biomass (
X) growth as basic building block and introduces a state variable called
photon flux density of photoacclimation,
, which represents the intensity at which the culture is photoacclimated at a specific physiological level, rather than the actual photon flux density to which it is exposed.
To model the time evolution of this additional variable, expressions based on the Beer–Lambert law for the absorbance of light in the culture medium are proposed. An average light irradiation is computed, which is the photon flux density that affects the culture mass on average,
. This parameter drives the dynamics of the acclimation phenomenon through a recall factor proportional to
. A full model development description can be found in [
25,
26]. This model has been implemented in several occasions [
20,
21,
24,
27,
29,
30,
31,
32], sometimes including simplifications.
The mass balance model of a photobioreactor for the main species in liquid and gaseous phases (including, substrates, products, and biomass concentration) is given by the general model representation [
33].
Taking the mathematical description of photo-acclimation into account and using appropriate kinetic expressions for
and
, the mass balances for
,
and
in a continuous stirred-tank reactor (CSTR) can be described as:
where
and
denote, respectively, the dilution rate and the respiration rate; and
, the nutrient concentration in the feed. This model assumes Monod kinetics—at low and moderate media concentration, Andrews inhibition kinetics—at high media concentration, Droop model for internal nutrient cell quota, and light-limited/photo-initiated Haldane inhibition kinetics.
The remaining model parameters are calculated according to the following expressions. In Nomenclature,
Table A1 contains a full description of the model nomenclature.
Average Photon Flux Density 3. Materials and Methods
The microalgae strain used in this work is
Scenedesmus obliquus (also known as
Tetradesmus obliquus), a freshwater microalgae, first reported in [
34]. The inoculum was obtained from the Culture Collection of Algae and Protozoa (CCAP 276/3A). The microalgae cultures are run into VWR
® 5 L borosilicate beakers, and their variables are monitored by measuring the biomass, nitrogen quota, substrate concentration, and chlorophyll (
) concentration with the techniques described in the following subsections.
3.1. Variable Measurement
3.1.1. Biomass Measurement
The standard off-line method for quantifying biomass concentration of the culture is the dry weight mass measurement [
35]. The procedure requires several steps for cell separation, washing, and drying to a constant weight. The main drawback, however, is the lack of distinction between living and dead cellular material when taking the measurement.
Operation Procedure
Place cellulose acetate membrane filters (Whatman® (Sigma-Aldrich, St. Louis, MI, USA)) inside an oven at 105 °C and leave them overnight.
Retrieve the dried filters and allow them to cool down in a desiccator for 8 h. Weigh the chosen filter and record the value .
Extract a sample from the culture. Filter the sample using a vacuum pump; this leaves the biomass separated on the membrane aside from the medium.
Dry the filter together with the algal biomass in a furnace at 105 °C until a constant weight is achieved and then cool in a desiccator for 20 min.
Weight the sample on an analytical balance and record the value .
Determine the biomass weight as the difference between and , with the corresponding factor adjustment.
In this study, a Sartorius® AZ214 M-Power Analytical Balance (Sartorius, Goettingen, Germany) was used, with a readability of ±0.0001 g and a repeatability of ±0.0003 g; while the volume of the samples was taken with a pipet of ±0.01 mL precision.
3.1.2. Quota Measurement
The nitrogen determination was carried out with a Shimadzu® TOC-VCSN & TNM-1 (Shimadzu, Kyoto, Japan) instrument, which provides measurements over a wide range from 0.1 mg/L to 4000 mg/L of nitrogen.
Operation Procedure
Take the sample from the medium culture into a 15 mL vial.
Centrifuge it for 5 min at 5000 rpm. Remove the supernatant and gently shake the vial to dilute biomass. Use demineralized water to fill up to 15 mL.
Repeat the previous step to wash the biomass and ensure a proper removal of substrate in the medium.
Process the sample with the equipment to obtain the total nitrogen content ().
The intracellular quota
is then derived from total nitrogen measurements and biomass concentration
:
The measurement of nitrogen quota has a ±0.01 mg/L sensitivity.
3.1.3. Substrate Measurement
In order to quantify the nitrate (substrate) concentration in the medium, a spectrophotometry method is used [
36]. When dissolved in water, nitrate absorbs UV light under 250 nm. Sample turbidity from organic material disturbs the absorbance measurement. To cope with this interference, two different measurements must be taken at specific wavelengths: 210 nm (
) and 270 nm (
). For the first determination, a peak is observed which contains the maximum nitrate absorption plus the so-called interference, whereas for the second one, only the effect of turbidity is considered. Accordingly, one can compute:
where
corresponds to the nitrate concentration from the optical density measurement.
A Shimadzu
® UVmini-1240 spectrophotometer is used, with a sensitivity of ±0.1 for absorbance measurements. The relationship between absorbance and nitrate concentration is obtained by preparing several dilutions of a nitrate solution. The absorbance difference (
) is then linearly correlated to the nitrate concentration,
:
where
is a linear correlation coefficient. On-line measurements can also be implemented using a deuterium light source and a spectrometer, connected to a fiber optics probe. The light travels through the probe across the culture medium, where it is absorbed, and eventually reaches the spectrometer.
Operation Procedure
Collect a 5 mL sample from the reactor.
Centrifuge the sample over 5 min at 5000 rpm.
Take out 4 mL of supernatant and introduce it into a spectrophotometer cuvette.
Measure the absorbance with the spectrophotometer at 210 nm and 270 nm.
Determine the concentration of the substrate according to the correlation curve.
A centrifugation step is accomplished before the absorbance measurement to eliminate the microalgae so that they do not interfere with the measurement. The volume samples are taken with a pipet of ±0.01 mL sensitivity.
3.1.4. Chlorophyll Measurement
For this propose, the extraction method described in [
37] is followed, changing the filtering step for a centrifugation one to generate less turbidity and manage a smaller sample volume. Additionally, a step of mechanical grinding is used in order to improve the chlorophyll extraction from the microalgae, as reported in [
38,
39].
Operation Procedure
Collect a 10 mL sample from the reactor in a 15 mL test tube.
Centrifuge the sample over 5 min at 5000 rpm.
Take out 9 mL of supernatant, add 3 mL of acetone and mix.
Transfer the content to a 10 mL capsule of a ball mill.
Add glass microspheres under 0.50 mm into the capsule, and shake it at 30 Hz for 20 min.
Remove the capsule and transfer the content to a test tube.
Fill the tube up to 10 mL of content adding acetone. This way the full content will be 90% acetone and 10% water, with the pigments solved in it.
Centrifuge over for 5 min at 5000 rpm.
Take out 4 mL of supernatant and transfer it to a 10 mm glass cuvette for the spectrophotometer.
Measure the absorption at 630, 647, 664, and 750 nm.
To consider that the extraction is properly executed, the measurement at 750 nm should be less than 0.010, while the absorption peak at 664 nm should not be higher than 1.000. The correlation between the absorbance and the concentration of the pigments is made with the formulas from [
40] for extraction with 90% acetone solvent. This sample should be processed just after being taken. Otherwise, after the first centrifugation and supernatant extraction, it can be preserved at −18 °C for up to one month.
3.1.5. Irradiance Measurement
Quantum sensors measure photosynthetically active radiation (PAR)—also named as photosynthetic photon flux density (PPFD)—which is the photon flux density in the spectral range (wave band) of solar radiation from 400–700 nm that photosynthetic organisms are able to use in the process of photosynthesis. An Apogee® MQ-200 Quantum Sensor (Apogee, Santa Monica, CA, USA) () was used to measure the light received by the culture.
3.2. Cell Culture and Medium Preparation
All cultures are maintained in heat-sterilized (121 °C over 20 min) modified Bold’s Basal Medium (BBM, see
Table 1) at an initial pH value of 6.8 [
41]. Prior to photobioreactor inoculation, the cultures were kept in an exponential growth phase for at least one week in Erlenmeyer flasks with constant agitation (250 rpm) and continuous light supply—no dark cycle—with constant photon flux density in the range
.
3.3. Laboratory Scale Process
The photobioreactors used are cylindrical VWR® (VWR International, Radnor, PA, USA) borosilicate glass beakers of 5 L with 17 cm in diameter with covers made of acrylic glass. These covers allow access to inoculation, culture sampling, nutrient addition, gas evacuation, and carbon dioxide injection. In all the conducted experiments, the total sample volume extracted was under 10% of the initial culture volume, a condition to be met so as to neglect the volume changes. The temperature was maintained in the range 22–24 °C thanks to a water bath which was adjusted manually at regular time intervals.
In autotrophic conditions, microalgae need light for their photosynthetic growth as an energy source, carbon dioxide as carbon source, as well as mineral nutrients such as nitrogen, phosphate, and sulfate sources. The carbon dioxide transfer, dissolution, and consumption lead to pH changes and, therefore, probes for pH (HAO SHI® H-101 industrial online pH-meter) (HAO SHI, Taiwan) and carbon dioxide injections are installed to monitor and control the pH in the culture and to provide a carbon source to the microalgae. The pH is set at a constant value of 7.0. For mixing proposes, a magnetic bar of 70 mm long is positioned at the bottom of each reactor and moved by magnetic stirrers placed under the beaker at a constant speed of 250 rpm.
The effect of different wavelengths, especially the one of red/blue light with different ratios comparing against red, blue, and white light is reported in [
42]. It is pointed out that light sources with red/blue lights in different ratios improve the growth rates of the microalgae. Other reports such as [
43] support the same conclusion, aside from other effects on chlorophyll concentration or protein concentration changes. Hence, it is recommended to provide the microalgae with a wide variety of wavelengths, with emphasis on the red spectrum where the chlorophyll absorbs the light. The light source available for our experiments was a ROLEADRO
® (Shenzhen Houyi Energy Efficiency Co. Ltd., Shenzhen, China) Dual Channel Dimming 600 W full spectrum LED panel, composed of a combination of 7 different wavelengths mainly contained in the visible spectrum between 400–700 nm. (410, 450, 470, 440, 460, 580, 610, 630, 660, and 740 nm; 3000 k, 6500 k). This light source makes emphasis on the red and blue light bands, in an 8:1 ratio. The light panel is capable of generating up to
from a close distance.
3.4. Experimental Design
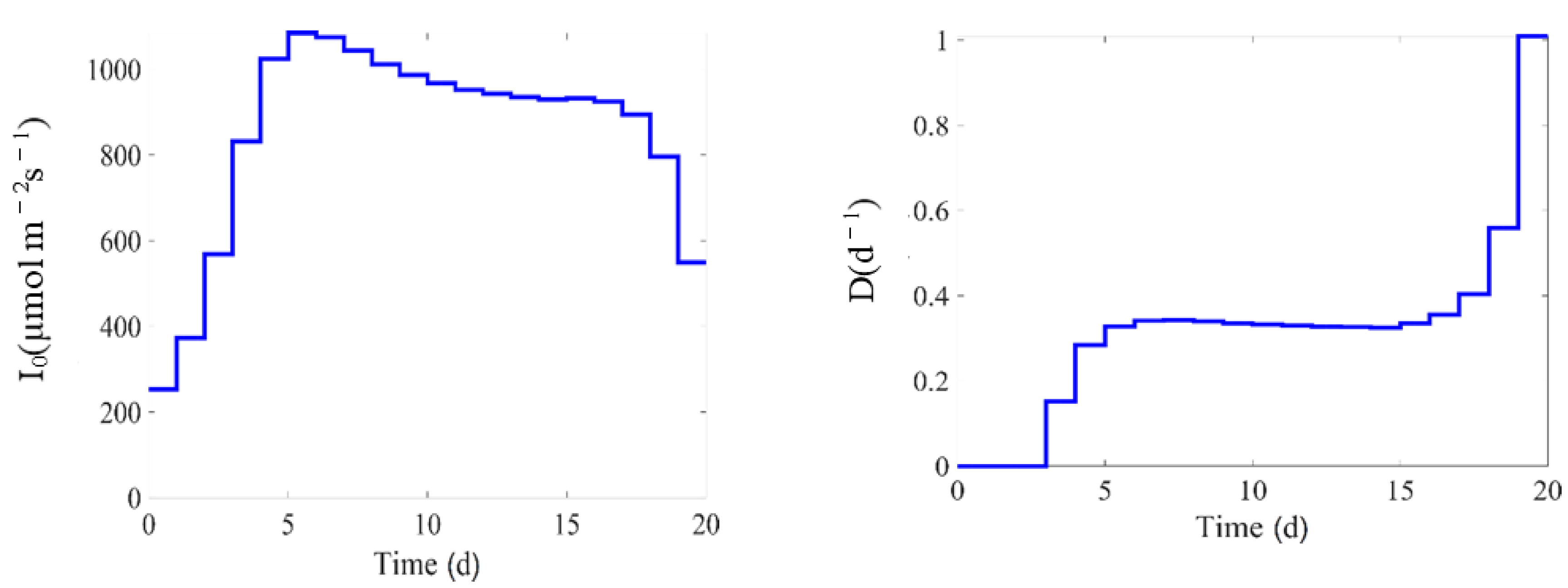
The experiments are designed to provide informative datasets for parameter identification. As a first approach, operational ranges are defined for the substrate concentration, the photon flux density, and the biomass concentration inside the bioreactors, so as to observe various phenomena related to substrate uptake, biomass growth, photo-acclimation, and inhibition. Based on previous experimental observations (from literature and our research group expertise), it is known that microalgae cultures can run from very small concentrations of biomass, to a couple of thousands of milligrams per liter, while the substrate concentration can range from as low as zero to around a hundred milligrams per liter. Hence, the substrate concentration is varied in the experiments between high levels where no limitation occurs, to very low levels where microalgae are in a nutrient-limited environment, and where the Droop kinetics is active. Chlorophyll is directly proportional to the level of the nitrogen quota and, at the same time, is highly dependent on the substrate concentration . Some runs are therefore conducted with high substrate concentrations, trying to decouple this effect and making more dependent over the acclimation photon flux density, . Last, the photon flux density operational range is explored at relatively low levels, especially over not so dense cultures, to collect data where microalgae do not express photoinhibition, and through the whole interval up to high levels, where photoinhibition can be detected and, at the same time, light penetration inside the dense culture is still possible.
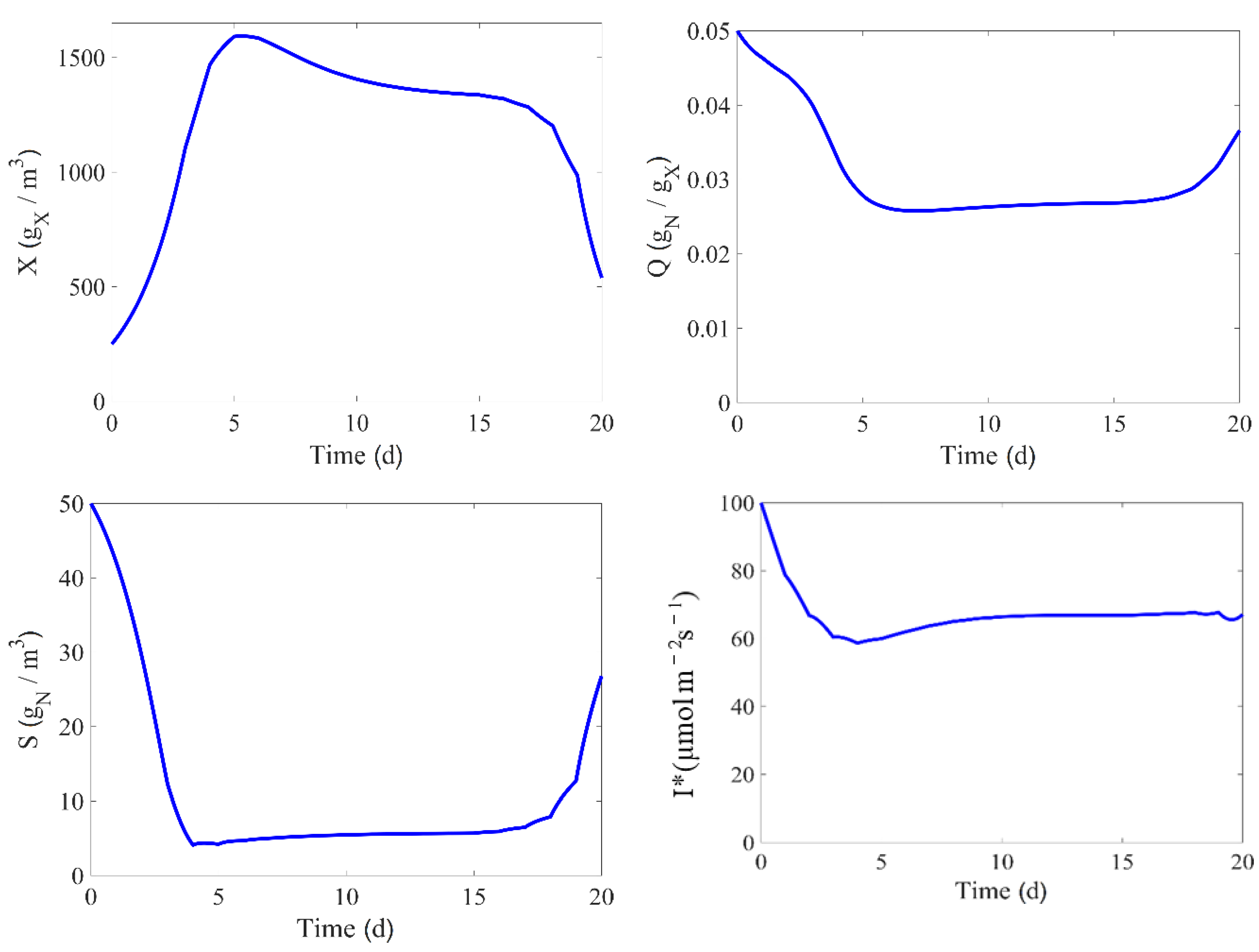
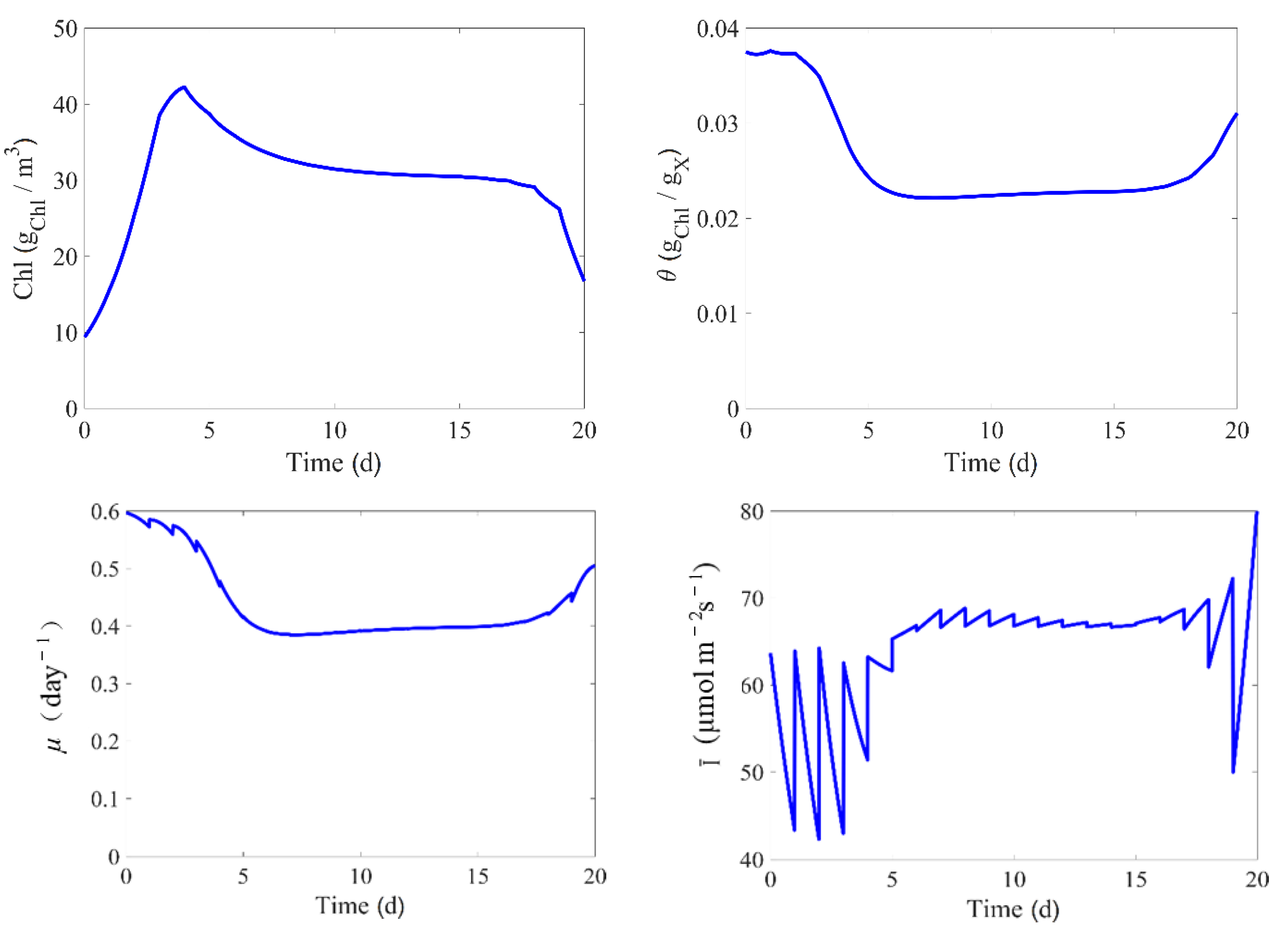
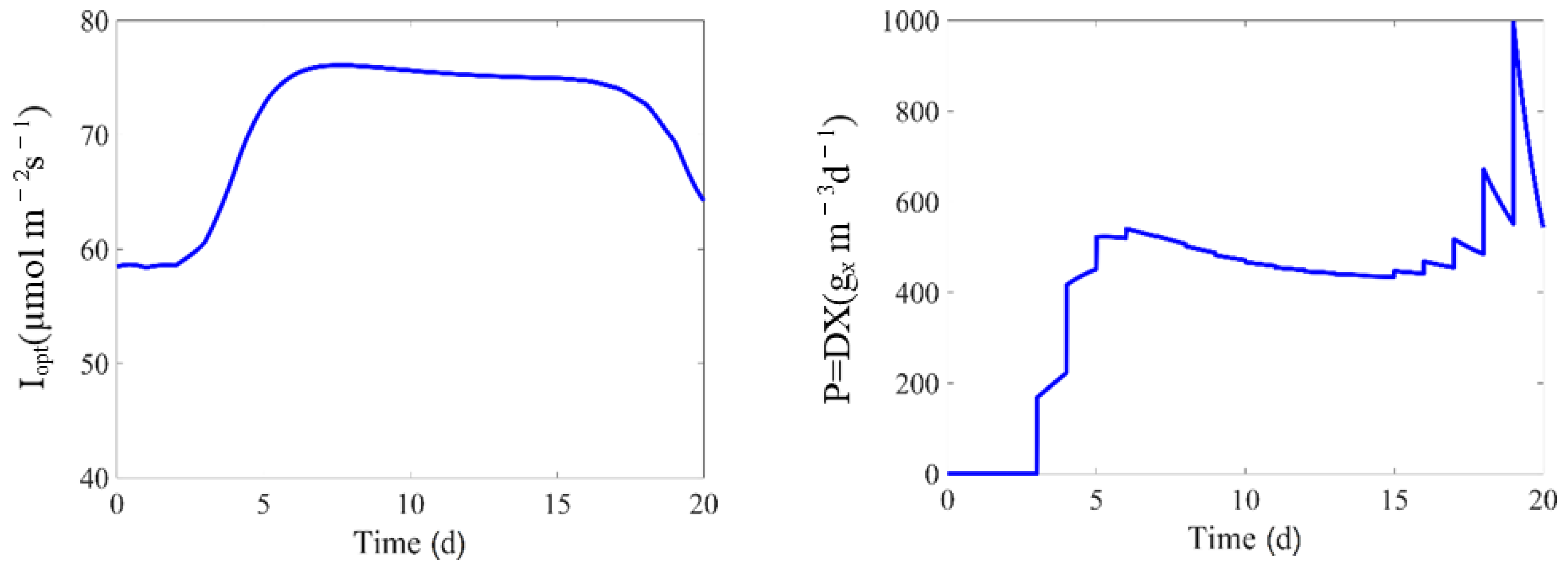
In some of the culture runs, the operational variables are adjusted in a way that approaches the optimal conditions for biomass growth. As discussed in [
44], this strategy tracks a performance path which is one of the ultimate goal of the model, and it is a good approach to excite the different kinetics and mechanisms along this path.
Table 2 shows the initial values of each state variable, the input variables values, and the cultivation time for each run.
5. Model Parameter Identification
For the model chosen in this work, parameters can be grouped into two categories: growth parameters (, , , , ,), and light parameters (, , , , , , , ). The first category actually contains parameters involved in many processes, like nutrient uptake kinetics, nutrient intracellular quota, respiration, and substrate-based growth kinetics, involving both factors, nutrient and light, respectively. The second category includes parameters relative to light absorption and scattering, and light acclimation. Roughly speaking, the identification of parameters in each category is favored by sets of data that excite one the most while diminishing the effects from the other category. Similarly, certain particular growth conditions are better for the determination of a certain parameter, e.g., internal quota lower and upper limits during starvation when nutrients are exhausted and during substrate repletion, respectively. The broad experimental conditions considered in this study contain a rich set of culture scenarios that excite the system in order to avoid potential parameter identifiability problems.
The model represented by Equations (2)–(5), can be expressed in a compact way as:
where
denotes the vector of state variables, and
is the vector of the
model parameters. Hence, using numerical integration, a solution of these nonlinear model equations provides a time profile of each state, which depends on the parameter set
.
where
stands for the output model prediction. Furthermore, the collected data at a time
can be represented in a vector form as:
where
represents the true value of the parameter vector, and
is the number of observation times or measurements. This last expression links the measured value
with the modeled result
. The idea is to try to match these two values through a multiparameter regression. The measurement errors are assumed to be independent, zero mean, and Gaussian.
Then, the identification of model parameters can be performed with a Weighted Least Squares (WLS) criterion, where the cost function
is formulated as:
and the weighting matrix can be selected as:
Note that the matrix
scales each variable for differences in physical units and orders of magnitude. An estimation of the covariance matrix of the measurement noise can be done as
where
with
the number of model variables,
the number of time samples, and
the number of model parameters. The estimation of the parameters is obtained by minimizing the cost function
.
Various methods can be used to solve this minimization problem, which is nonconvex. For instance, a two-step procedure can be used where a genetic algorithm (GA)—i.e., ga in MATLAB® (MathWorks, Natick, MA, USA)—is executed for the first step. With enough generations it allows locating a global minimum, avoiding falling into local minima. The second step can be achieved with the lsqnonlin function from MATLAB®. Alternatively, the minimization of the cost function could be carried out using some local minimization method combined with a multi-start strategy which allows locating some of the local minima, and hopefully determining the global one. In this case, it is required to define a restricted parameter space and a set of initial parameter guesses, which can be obtained through a Latin Hypercube Sampling.
The Fisher Information Matrix,
, is then calculated as follows:
where
are the sensitivity matrices.
The analytical computation of these matrices for the model under consideration in this study is not an easy task. However, the sensitivity matrices can be obtained as a by-product of the
lsqnonlin algorithm. The parameter covariance matrix is then approximated—assuming an unbiased estimator, as:
The standard deviation
of the parameter estimates
is obtained.
The 95% confidence intervals (
) for each parameter can be calculated as
. The coefficient of variation (
) is then given by:
The computation of the confidence intervals can be easily achieved with the MATLAB
® command
nlparci. An alternative procedure uses the outputs of
nlinfit as inputs to
nlparci. The parameters and their respective confidence intervals are listed in
Table 3.
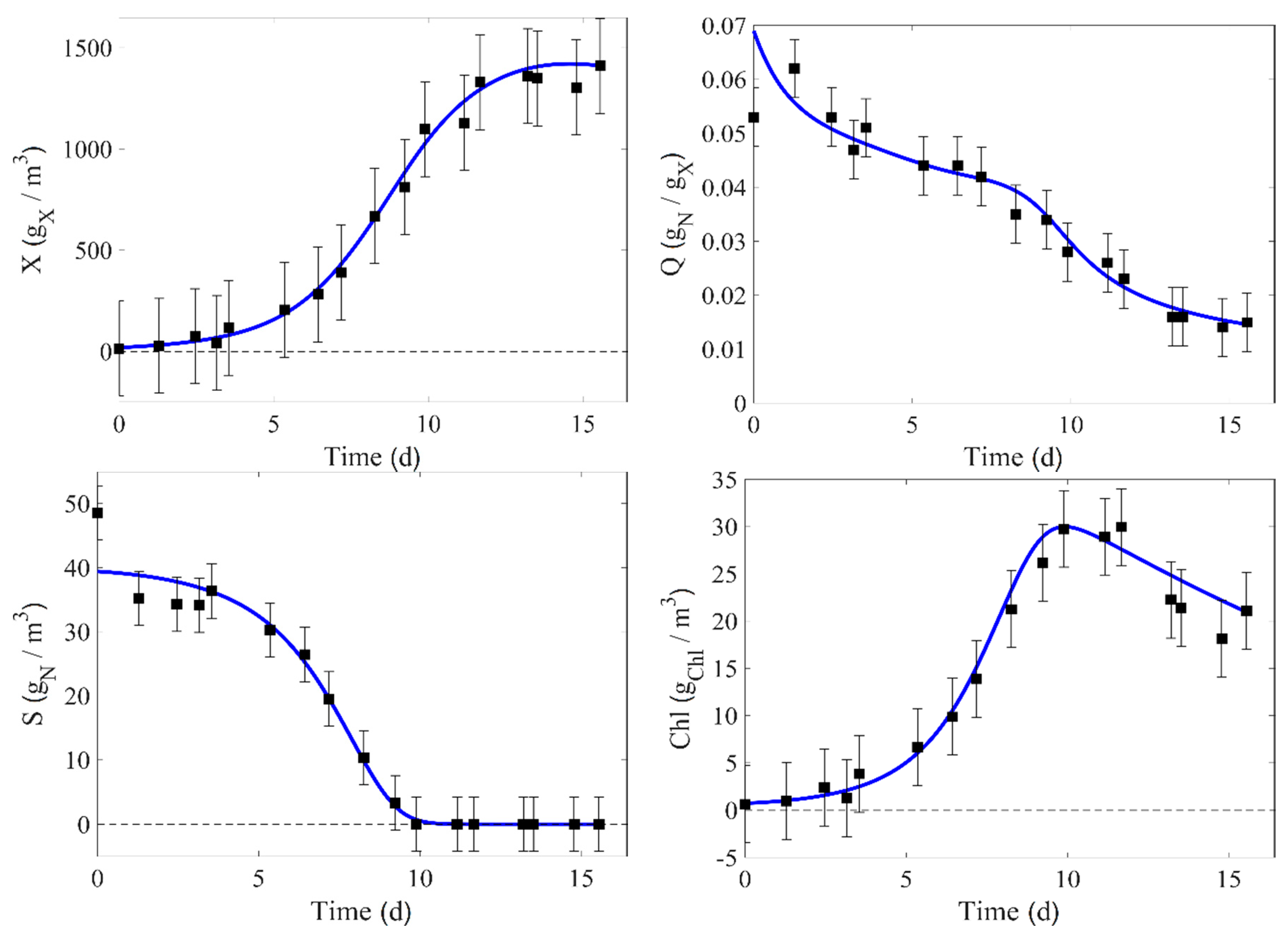
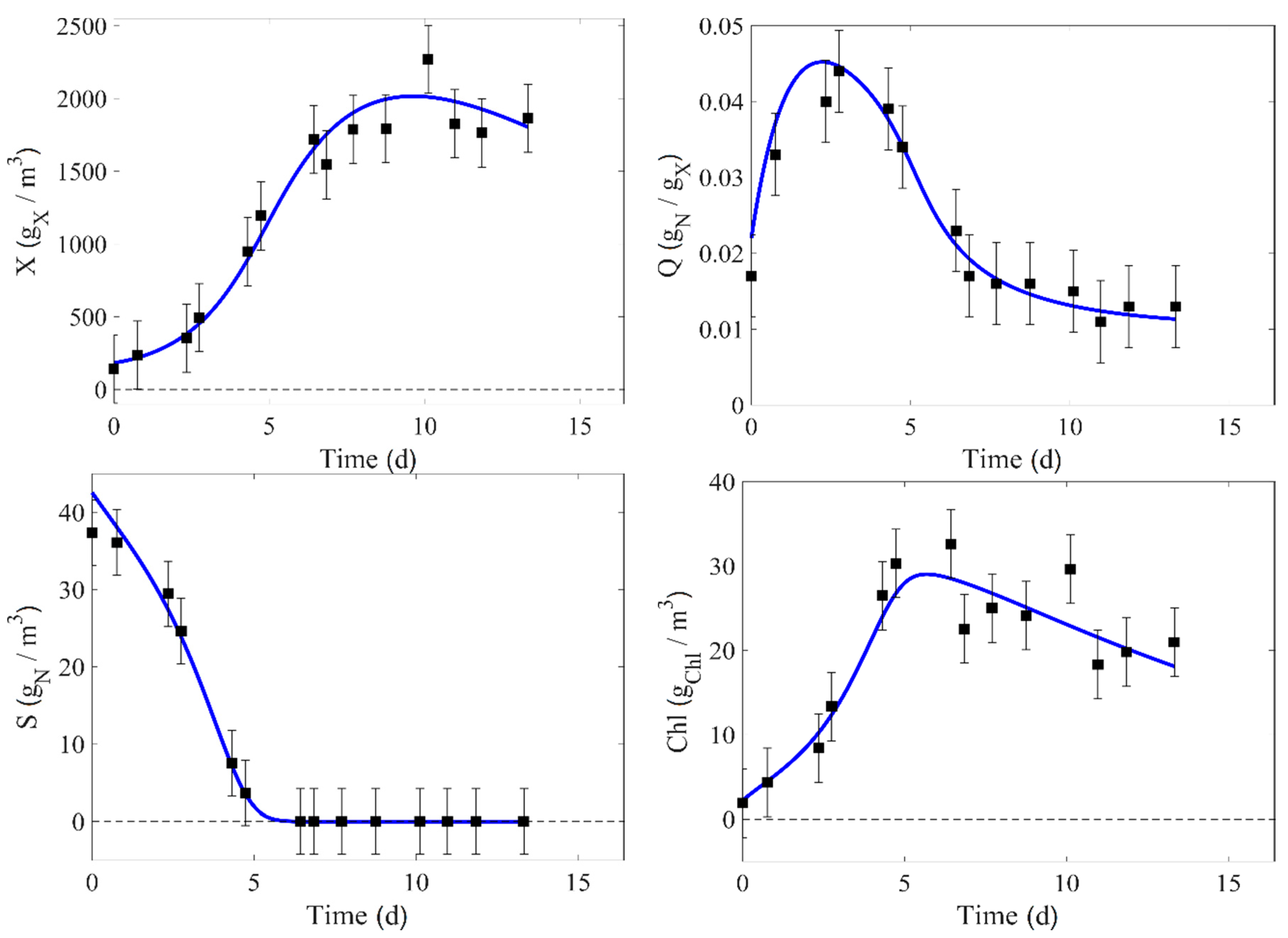
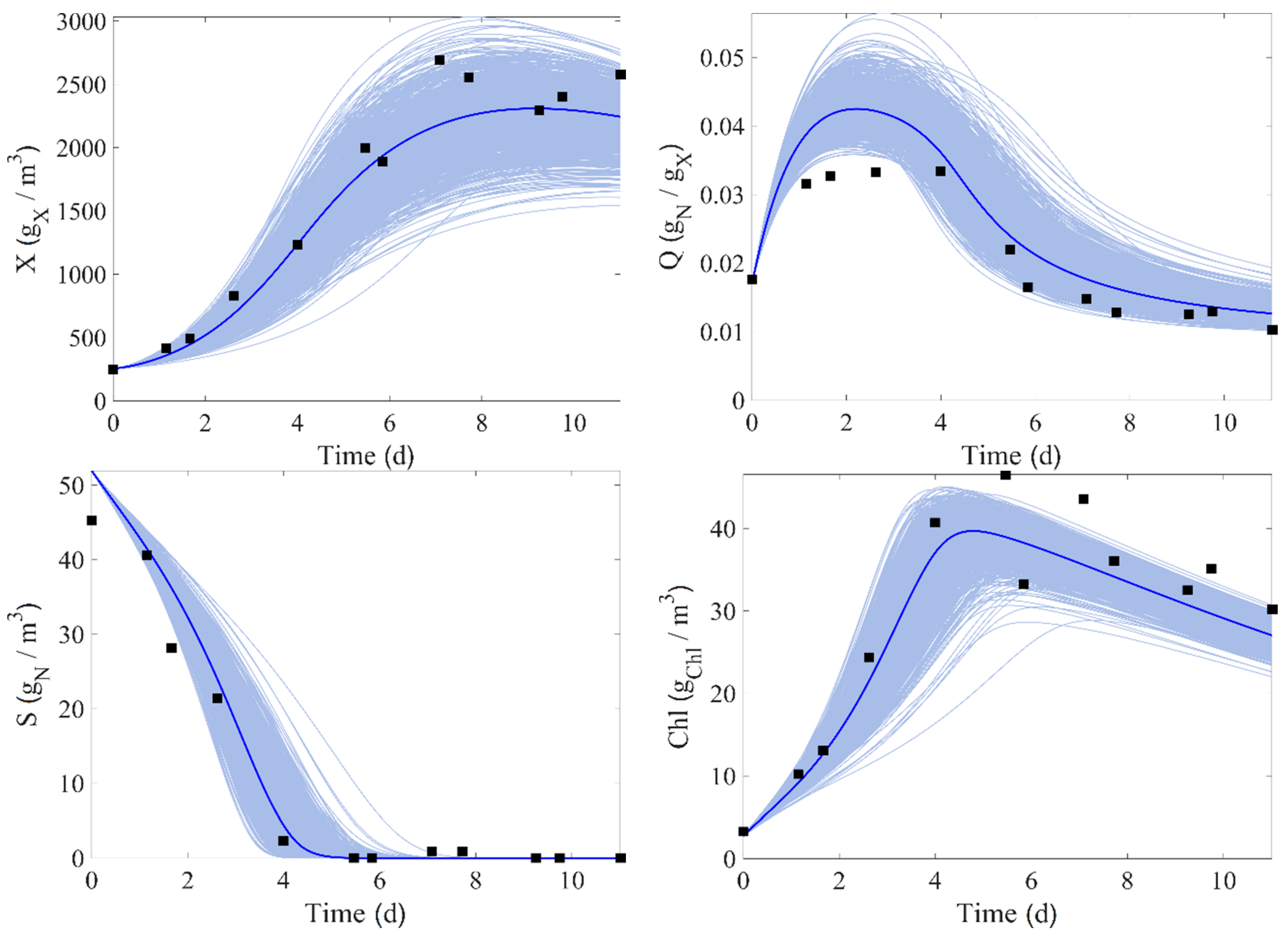
In this work, there is a total of nine experimental runs. Seven of them (runs 1–4, 6–7, and 9) are used for the identification process and direct validation. The experimental data of runs 4 and 9 are compared to the corresponding model prediction in
Figure 2 and
Figure 3, where the confidence intervals of the measurement at 95% are also displayed (these intervals are estimated from Equation (24)).
An important point is that not only the model parameters are estimated but also the initial conditions of the model state variables. Indeed, these initial values are measured as any other data point and they are therefore uncertain. Nonetheless, they play a critical role in the model integration and a good identification practice is to not completely trust the measured initial conditions as they are corrupted by measurement noise, and thus, to not force the model to exactly start from these points. Hence, initial conditions are estimated as part of the identification procedure. This gives the model more flexibility to fit the datasets. However, the combined identification of the model parameters and initial conditions (of each experiment) leads to a relatively large problem which can be delicate to solve at once. The procedure is therefore subdivided into two steps. It is initially assumed that the errors in the initial conditions are relatively small, and only the model parameters are estimated. In a second step, the model parameters are fixed to their estimated values and the initial conditions are estimated. This two-step process is performed iteratively until there is a convergence—usually achieved in a couple of iterations.
Table 4 contains the estimated initial conditions.
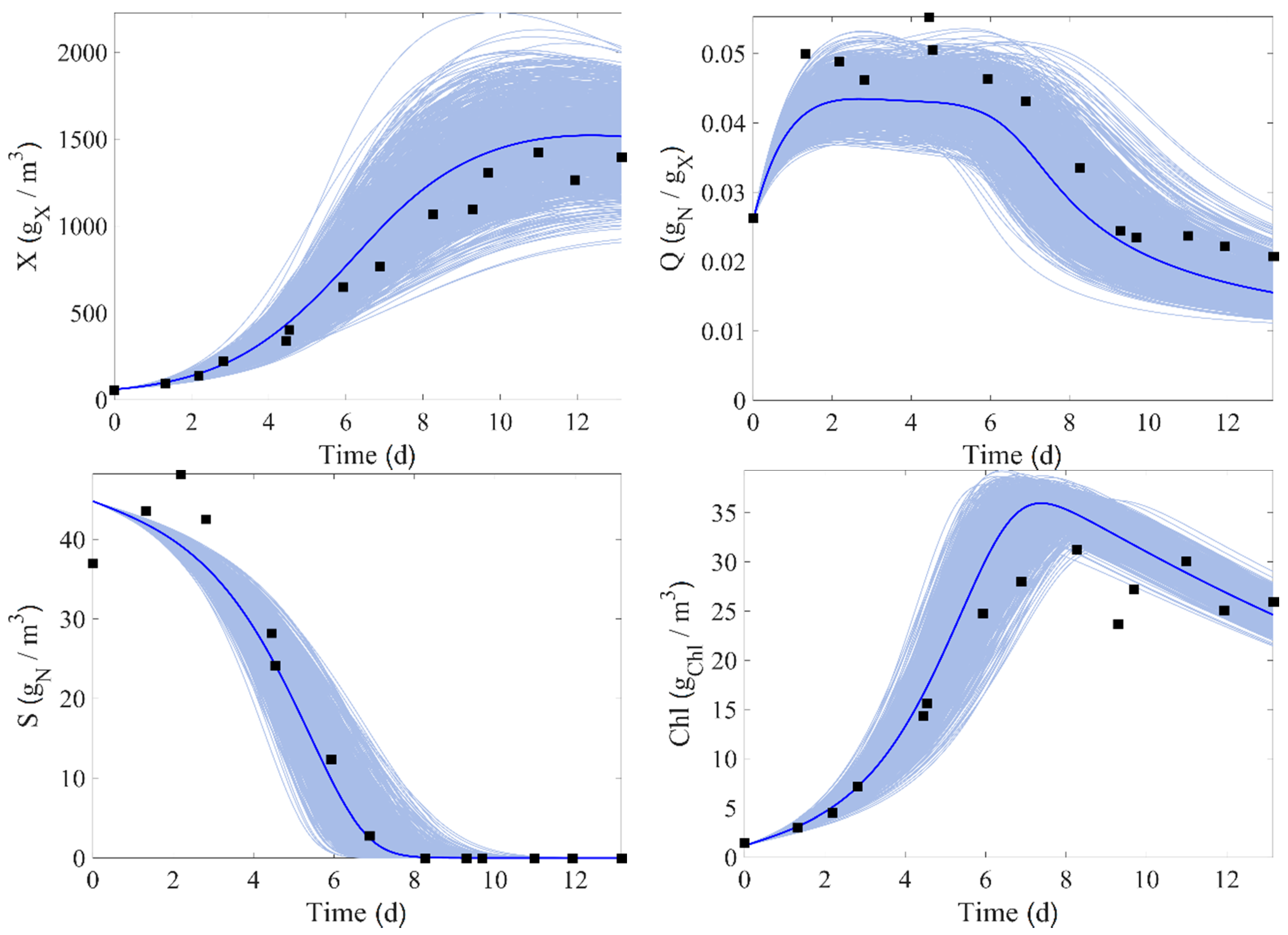
In order to check the model prediction under parameter uncertainty and to compare it in cross-validation, simulations are carried out using a Monte Carlo approach. The parameter set is generated with a Latin Hypercube Sampling (LHS), which can be achieved using the MATLAB
® commands
lhsdesign or
lhsnorm using the estimated mean and standard deviation of the parameters. For this purpose, runs 5 and 8 are considered and 1000 simulation runs are produced with sampled model parameters (the initial conditions are fixed in this process).
Figure 4 and
Figure 5 show the results, where the light blue areas represent the range of values taken by each state variables. It can be seen that the trend of the data is well followed by the model, and the experimental data are almost entirely inside the uncertainty intervals.
At this stage, it is interesting to compare our results against the parameters identified in previous works (see
Table 5). One of them is [
26], where the identification is performed for another strain,
Isochrysis galbana and a different version of the model, based on experimental data reported in [
45]. Another one is [
29], which is based on the model originally reported in [
25], with a few simplifications in some physiological variables equations, and experimental data for the same strain,
Scenedesmus obliquus cultivated in large cylindrical photobioreactors.
The first study identified the model parameters based on a dataset with a relatively narrow range of conditions corresponding to low biomass concentrations and low photon flux density, as it can be seen in [
45]. The second study has achieved the model identification using measurements of biomass, nitrogen quota, and substrate concentration, but chlorophyll was not available at the time, which implies identifiability problems. In this work, our objective was to provide a wider range of operating conditions and to include the measurement of chlorophyll, in an attempt to have a more precise determination of parameters involved in the acclimation and photoinhibition processes.
The reported values of in these previous identification studies are relatively small compared to the actual substrate concentrations, which is not the case in the present study. Considering that it represents the half-saturation coefficient of a Monod law, therefore, the kinetic factor approaches unity, making it irrelevant. The same happens with parameter . When comparing the operational values of the optical depth with , the value of is noticeably smaller, which makes the average photon flux density received throughout the reactor almost negligible for most operational conditions, which is probably not realistic. Taking a look at the coefficient , their values are in agreement in the three studies and notably high, which implies that chlorophyll has a strong shading effect in the culture, aside from the biomass darkening effect. The majority of higher impact parameters, like the maximum specific growth rate, , the maximum nitrogen intake rate, , the minimum and maximum nitrogen quota, and , the attenuation coefficient due to chlorophyll, , and the attenuation coefficient due to biomass, , they all have similar values within the same magnitude order.

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








