
Expression of Secreted Neutrophil Gelatinase-Associated Lipocalin in 293T Cell Using the Inducible Dual-Function System

 , , , and
, , , and 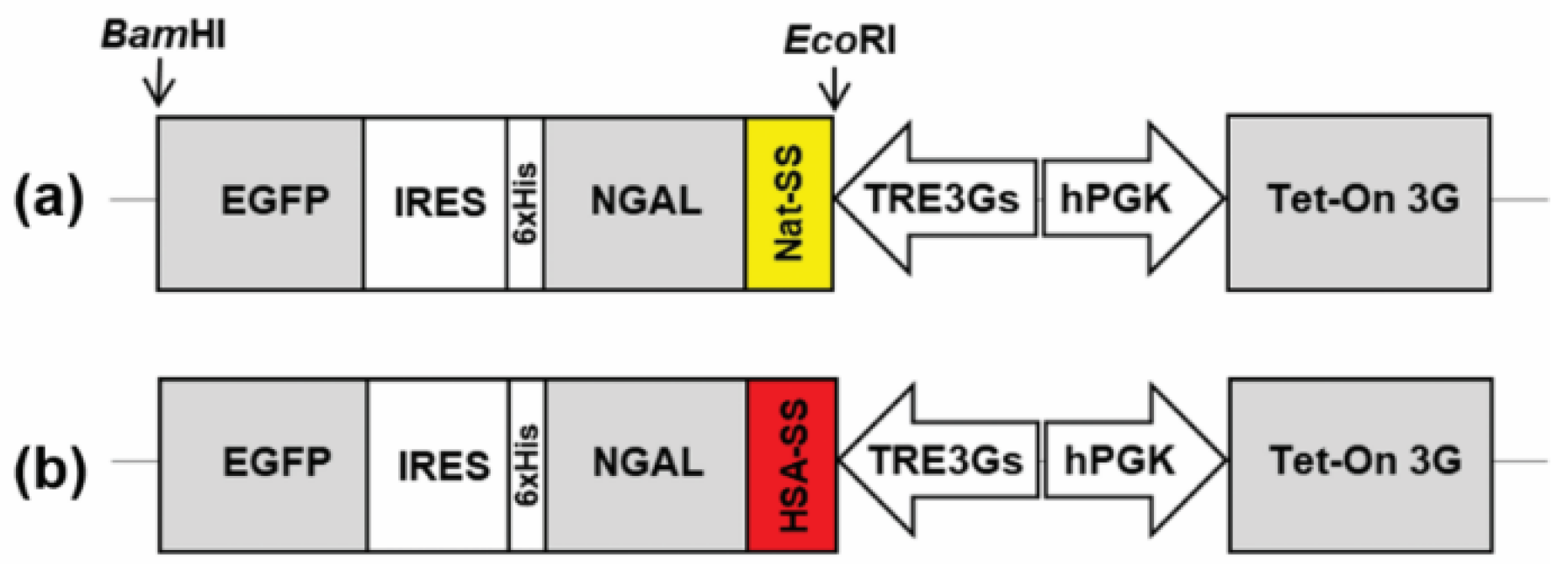{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Design and Construction of NGAL-Expressing Tet-On Vectors
2.2. Cell Transfection and Supernatant Collection
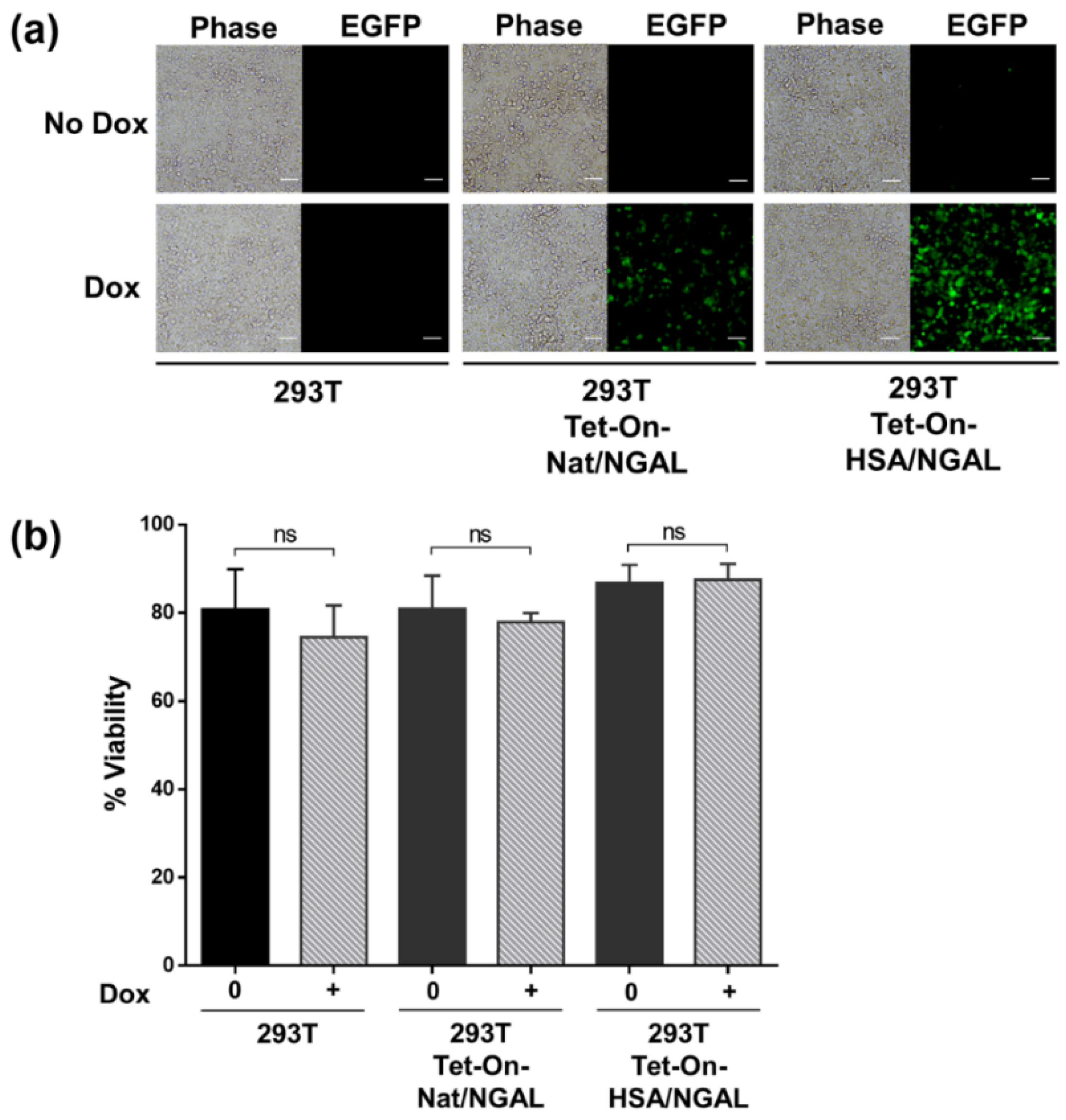
2.3. Cell Viability
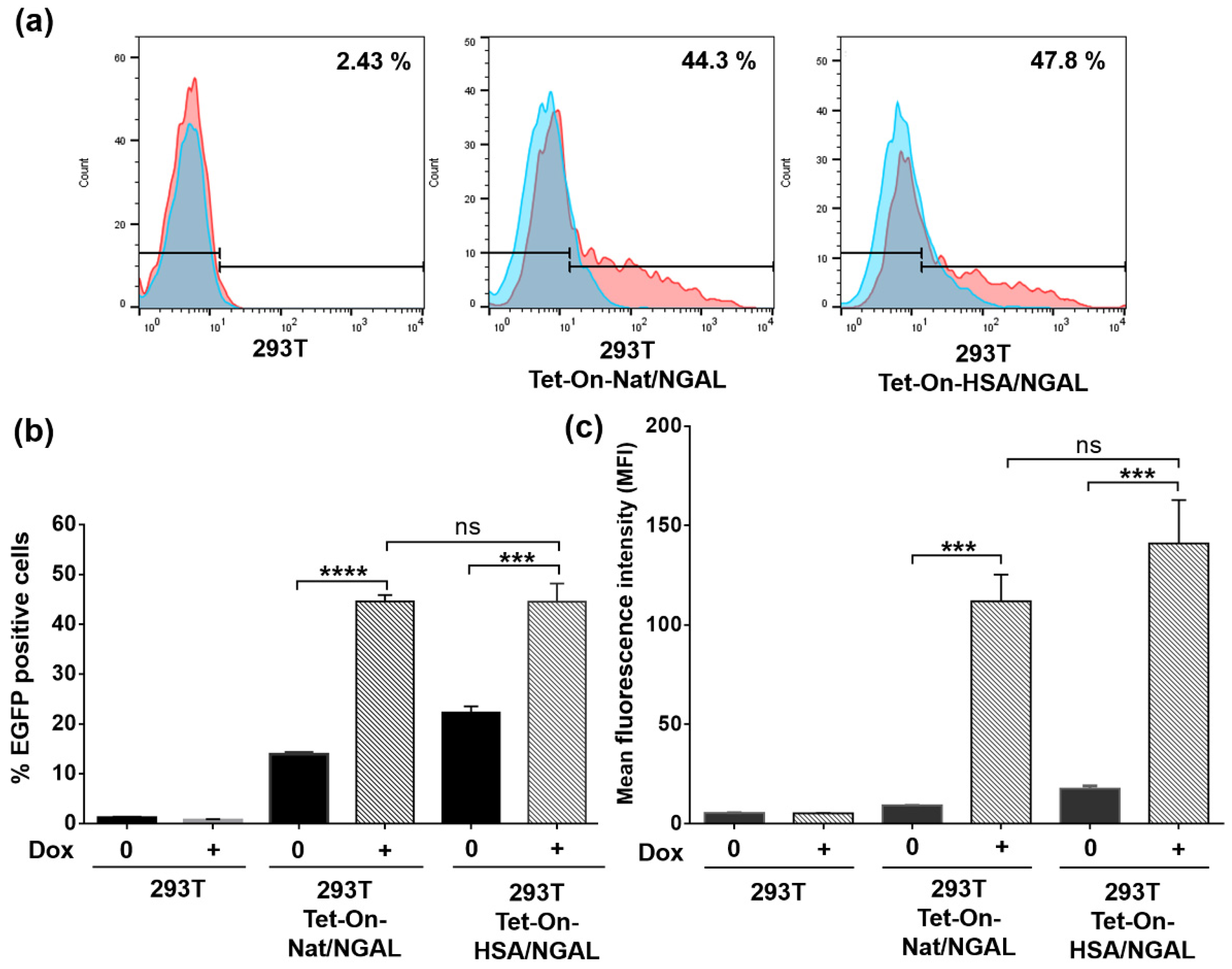
2.4. Evaluation of EGFP Expression by Flow Cytometry
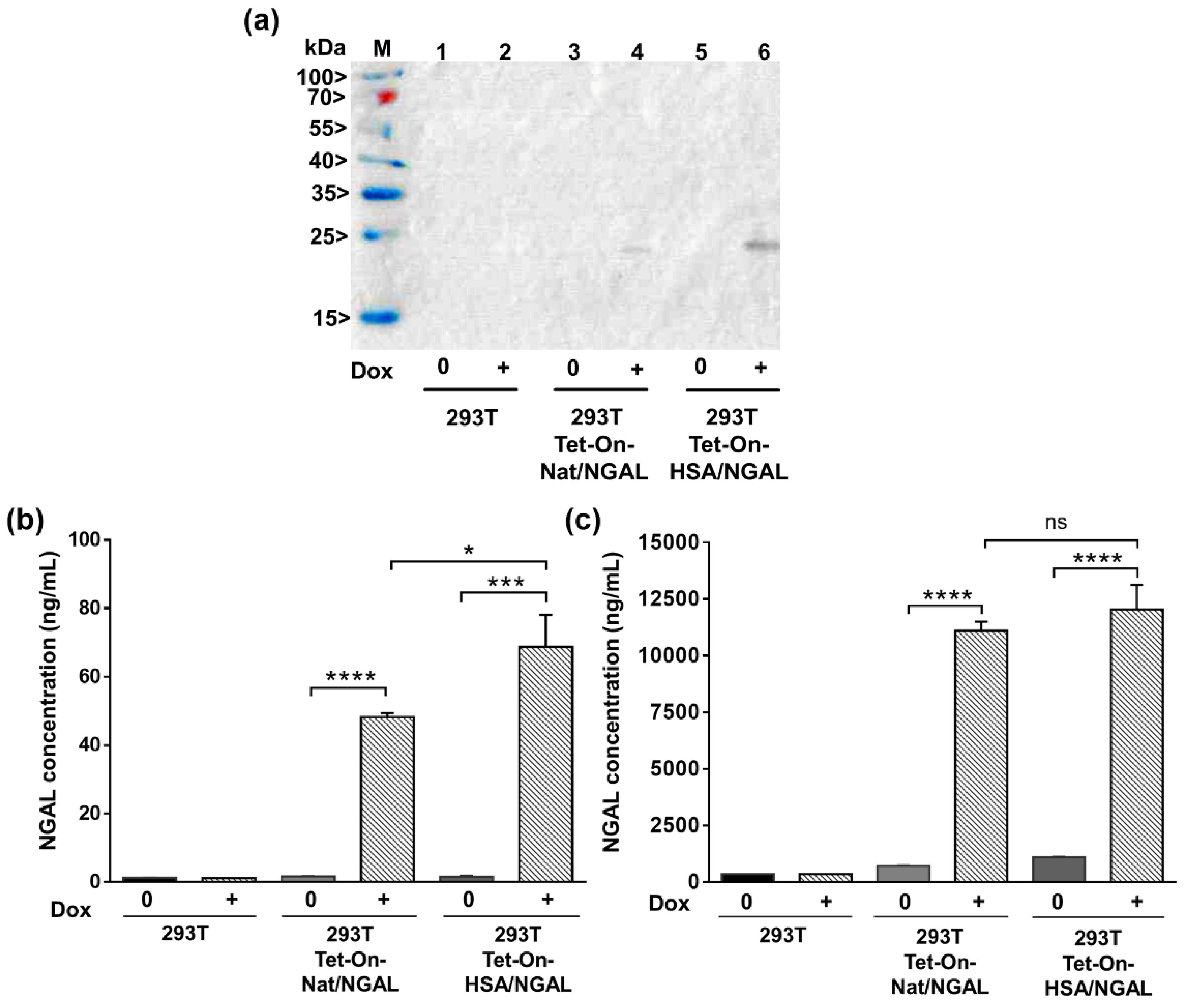
2.5. Detection of Soluble NGAL by Western Immunoblotting
2.6. Determination of NGAL Concentration
2.7. Statistical Analysis
3. Results
3.1. Construction of a Dox-Inducible Vector for Dual Gene Expression
3.2. Dox-Induced EGFP Expression in Transfected 293T Cells
3.3. Level of EGFP Expression in Transfected Cells
3.4. Quantification of Secreted NGAL in the Supernatant
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mishra, J.; Dent, C.; Tarabishi, R.; Mitsnefes, M.M.; Ma, Q.; Kelly, C.; Ruff, S.M.; Zahedi, K.; Shao, M.; Bean, J.; et al. Neutrophil gelatinase-associated lipocalin (NGAL) as a biomarker for acute renal injury after cardiac surgery. Lancet 2005, 365, 1231–1238. [Google Scholar] [CrossRef]
- Slocum, J.L.; Heung, M.; Pennathur, S. Marking renal injury: Can we move beyond serum creatinine? Transl. Res. 2012, 159, 277–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellomo, R.; Ronco, C.; Kellum, J.A.; Mehta, R.L.; Palevsky, P.; Acute Dialysis Quality Initiative Workgroup. Acute renal failure—definition, outcome measures, animal models, fluid therapy and information technology needs: The Second International Consensus Conference of the Acute Dialysis Quality Initiative (ADQI) Group. Crit. Care. 2004, 8, R204–R212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolignano, D.; Donato, V.; Coppolino, G.; Campo, S.; Buemi, A.; Lacquaniti, A.; Buemi, M. Neutrophil gelatinase-associated lipocalin (NGAL) as a marker of kidney damage. Am. J. Kidney Dis. 2008, 52, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Kjeldsen, L.; Johnsen, A.H.; Sengelov, H.; Borregaard, N. Isolation and primary structure of NGAL, a novel protein associated with human neutrophil gelatinase. J. Biol. Chem. 1993, 268, 10425–10432. [Google Scholar] [CrossRef]
- Nickolas, T.L.; O’Rourke, M.J.; Yang, J.; Sise, M.E.; Canetta, P.A.; Barasch, N.; Buchen, C.; Khan, F.; Mori, K.; Giglio, J.; et al. Sensitivity and specificity of a single emergency department measurement of urinary neutrophil gelatinase-associated lipocalin for diagnosing acute kidney injury. Ann. Intern. Med. 2008, 148, 810–819. [Google Scholar] [CrossRef]
- Bennett, M.; Dent, C.L.; Ma, Q.; Dastrala, S.; Grenier, F.; Workman, R.; Syed, H.; Ali, S.; Barasch, J.; Devarajan, P. Urine NGAL predicts severity of acute kidney injury after cardiac surgery: A prospective study. Clin. J. Am. Soc. Nephrol. 2008, 3, 665–673. [Google Scholar] [CrossRef] [Green Version]
- Krzeminska, E.; Wyczalkowska-Tomasik, A.; Korytowska, N.; Paczek, L. Comparison of Two Methods for Determination of NGAL Levels in Urine: ELISA and CMIA. J. Clin. Lab. Anal. 2016, 30, 956–960. [Google Scholar] [CrossRef] [Green Version]
- Roudkenar, M.H.; Halabian, R.; Roushandeh, A.M.; Nourani, M.R.; Masroori, N.; Ebrahimi, M.; Nikogoftar, M.; Rouhbakhsh, M.; Bahmani, P.; Najafabadi, A.J.; et al. Lipocalin 2 regulation by thermal stresses: Protective role of Lcn2/NGAL against cold and heat stresses. Exp. Cell Res. 2009, 315, 3140–3151. [Google Scholar] [CrossRef]
- Bundgaard, J.R.; Sengelov, H.; Borregaard, N.; Kjeldsen, L. Molecular cloning and expression of a cDNA encoding NGAL: A lipocalin expressed in human neutrophils. Biochem. Biophys. Res. Commun. 1994, 202, 1468–1475. [Google Scholar] [CrossRef]
- Bakhshandeh, Z.; Halabian, R.; Imani Fooladi, A.A.; Jahanian-Najafabadi, A.; Jalili, M.A.; Roudkenar, M.H. Recombinant human lipocalin 2 acts as an antibacterial agent to prevent platelet contamination. Hematology 2014, 19, 487–492. [Google Scholar] [CrossRef]
- Dumont, J.; Euwart, D.; Mei, B.; Estes, S.; Kshirsagar, R. Human cell lines for biopharmaceutical manufacturing: History, status, and future perspectives. Crit. Rev. Biotechnol. 2016, 36, 1110–1122. [Google Scholar] [CrossRef] [Green Version]
- Bosques, C.J.; Collins, B.E.; Meador, J.W., III; Sarvaiya, H.; Murphy, J.L.; Dellorusso, G.; Bulik, D.A.; Hsu, I.H.; Washburn, N.; Sipsey, S.F.; et al. Chinese hamster ovary cells can produce galactose-α-1,3-galactose antigens on proteins. Nat. Biotechnol. 2010, 28, 1153–1156. [Google Scholar] [CrossRef] [Green Version]
- Loignon, M.; Perret, S.; Kelly, J.; Boulais, D.; Cass, B.; Bisson, L.; Afkhamizarreh, F.; Durocher, Y. Stable high volumetric production of glycosylated human recombinant IFNalpha2b in HEK293 cells. BMC Biotechnol. 2008, 8, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iurlaro, R.; Munoz-Pinedo, C. Cell death induced by endoplasmic reticulum stress. FEBS J. 2016, 283, 2640–2652. [Google Scholar] [CrossRef] [Green Version]
- Chevallier, V.; Andersen, M.R.; Malphettes, L. Oxidative stress-alleviating strategies to improve recombinant protein production in CHO cells. Biotechnol. Bioeng. 2020, 117, 1172–1186. [Google Scholar] [CrossRef] [Green Version]
- Shuen, W.H.; Kan, R.; Yu, Z.; Lung, H.L.; Lung, M.L. Novel lentiviral-inducible transgene expression systems and versatile single-plasmid reporters for in vitro and in vivo cancer biology studies. Cancer Gene Ther. 2015, 22, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Das, A.T.; Tenenbaum, L.; Berkhout, B. Tet-On Systems For Doxycycline-inducible Gene Expression. Curr. Gene Ther. 2016, 16, 156–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, M.; Yuan, P.; Vavilala, D.; Fox, M. Optimization of Protein Expression in Mammalian Cells. Curr. Protoc. Protein Sci. 2019, 95, e77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darzacq, X.; Singer, R.H.; Shav-Tal, Y. Dynamics of transcription and mRNA export. Curr. Opin. Cell Biol. 2005, 17, 332–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiering, S.; Whitelaw, E.; Martin, D.I. To be or not to be active: The stochastic nature of enhancer action. BioEssays 2000, 22, 381–387. [Google Scholar] [CrossRef]
- Barnes, L.M.; Bentley, C.M.; Dickson, A.J. Molecular definition of predictive indicators of stable protein expression in recombinant NS0 myeloma cells. Biotechnol. Bioeng. 2004, 85, 115–121. [Google Scholar] [CrossRef] [PubMed]
- You, M.; Yang, Y.; Zhong, C.; Chen, F.; Wang, X.; Jia, T.; Chen, Y.; Zhou, B.; Mi, Q.; Zhao, Q.; et al. Efficient mAb production in CHO cells with optimized signal peptide, codon, and UTR. Appl. Microbiol. Biotechnol. 2018, 102, 5953–5964. [Google Scholar] [CrossRef]
- Chen, W.; Zhao, X.; Zhang, M.; Yuan, Y.; Ge, L.; Tang, B.; Xu, X.; Cao, L.; Guo, H. High-efficiency secretory expression of human neutrophil gelatinase-associated lipocalin from mammalian cell lines with human serum albumin signal peptide. Protein Expr. Purif. 2016, 118, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Moonmuang, S.; Saoin, S.; Chupradit, K.; Sakkhachornphop, S.; Israsena, N.; Rungsiwiwut, R.; Tayapiwatana, C. Modulated expression of the HIV-1 2LTR zinc finger efficiently interferes with the HIV integration process. Biosci. Rep. 2018, 38, BSR20181109. [Google Scholar] [CrossRef] [Green Version]
- Loew, R.; Heinz, N.; Hampf, M.; Bujard, H.; Gossen, M. Improved Tet-responsive promoters with minimized background expression. BMC Biotechnol. 2010, 10, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, H.; Maldonado-Agurto, R.; Dickson, A.J. The endoplasmic reticulum and unfolded protein response in the control of mammalian recombinant protein production. Biotechnol. Lett. 2014, 36, 1581–1593. [Google Scholar] [CrossRef]
- Tigges, M.; Fussenegger, M. Xbp1-based engineering of secretory capacity enhances the productivity of Chinese hamster ovary cells. Metab. Eng. 2006, 8, 264–272. [Google Scholar] [CrossRef]
- Nelson, A.M.; Zhao, W.; Gilliland, K.L.; Zaenglein, A.L.; Liu, W.; Thiboutot, D.M. Neutrophil gelatinase-associated lipocalin mediates 13-cis retinoic acid-induced apoptosis of human sebaceous gland cells. J. Clin. Investig. 2008, 118, 1468–1478. [Google Scholar] [CrossRef] [Green Version]
- Devireddy, L.R.; Gazin, C.; Zhu, X.; Green, M.R. A cell-surface receptor for lipocalin 24p3 selectively mediates apoptosis and iron uptake. Cell 2005, 123, 1293–1305. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.J.; Oh, B.M.; Kim, J.T.; Lim, J.; Park, S.Y.; Hwang, Y.S.; Baek, K.E.; Kim, B.Y.; Choi, I.; Lee, H.G. Efficient Interleukin-21 Production by Optimization of Codon and Signal Peptide in Chinese Hamster Ovarian Cells. J. Microbiol. Biotechnol. 2019, 29, 304–310. [Google Scholar] [CrossRef]
- Kober, L.; Zehe, C.; Bode, J. Optimized signal peptides for the development of high expressing CHO cell lines. Biotechnol. Bioeng. 2013, 110, 1164–1173. [Google Scholar] [CrossRef]
- Attallah, C.; Etcheverrigaray, M.; Kratje, R.; Oggero, M. A highly efficient modified human serum albumin signal peptide to secrete proteins in cells derived from different mammalian species. Protein Expr. Purif. 2017, 132, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Costello, A.; Lao, N.T.; Gallagher, C.; Capella Roca, B.; Julius, L.A.N.; Suda, S.; Ducree, J.; King, D.; Wagner, R.; Barron, N.; et al. Leaky Expression of the TET-On System Hinders Control of Endogenous miRNA Abundance. Biotechnol. J. 2019, 14, 1800219. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Vink, M.; Klaver, B.; Berkhout, B.; Das, A.T. Optimization of the Tet-On system for regulated gene expression through viral evolution. Gene Ther. 2006, 13, 1382–1390. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Suzuki, Y.; Nagasaki, S.C.; Okuno, H.; Imayoshi, I. Light Control of the Tet Gene Expression System in Mammalian Cells. Cell Rep. 2018, 25, 487–500.e6. [Google Scholar] [CrossRef] [Green Version]
- Ghassemi, F.; Madadgar, O.; Roohvand, F.; Rasekhian, M.; Etemadzadeh, M.H.; Boroujeni, G.R.N.; Langroudi, A.G.; Azadmanesh, K. Translational efficiency of BVDV IRES and EMCV IRES for T7 RNA polymerase driven cytoplasmic expression in mammalian cell lines. Mol. Biol. 2017, 51, 283–292. [Google Scholar] [CrossRef]
- Renaud-Gabardos, E.; Hantelys, F.; Morfoisse, F.; Chaufour, X.; Garmy-Susini, B.; Prats, A.C. Internal ribosome entry site-based vectors for combined gene therapy. World J. Exp. Med. 2015, 5, 11–20. [Google Scholar] [CrossRef]
- Wiznerowicz, M.; Fong, A.Z.; Mackiewicz, A.; Hawley, R.G. Double-copy bicistronic retroviral vector platform for gene therapy and tissue engineering: Application to melanoma vaccine development. Gene Ther. 1997, 4, 1061–1068. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.M.; Chen, C.J.; Shih, S.R. Regulation Mechanisms of Viral IRES-Driven Translation. Trends Microbiol. 2017, 25, 546–561. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saoin, S.; Boonkrai, C.; Pisitkun, T.; Kloypan, C.; Nangola, S. Expression of Secreted Neutrophil Gelatinase-Associated Lipocalin in 293T Cell Using the Inducible Dual-Function System. Processes 2021, 9, 855. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9050855
Saoin S, Boonkrai C, Pisitkun T, Kloypan C, Nangola S. Expression of Secreted Neutrophil Gelatinase-Associated Lipocalin in 293T Cell Using the Inducible Dual-Function System. Processes. 2021; 9(5):855. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9050855
Chicago/Turabian StyleSaoin, Somphot, Chatikorn Boonkrai, Trairak Pisitkun, Chiraphat Kloypan, and Sawitree Nangola. 2021. "Expression of Secreted Neutrophil Gelatinase-Associated Lipocalin in 293T Cell Using the Inducible Dual-Function System" Processes 9, no. 5: 855. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9050855