
Metabolomic Profiling, Antioxidant and Antimicrobial Activity of Bidens pilosa

 ,
, 
 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical and Reagents
2.2. Plants Material
2.3. Untargeted LC-MS/MS-Based Metabolomics
2.4. Determination of the Antioxidant Activity
2.5. HPLC–DAD-MS Determination of Phenolic Compounds
2.6. Antimicrobial Tests
2.7. Bioinformatics
3. Results
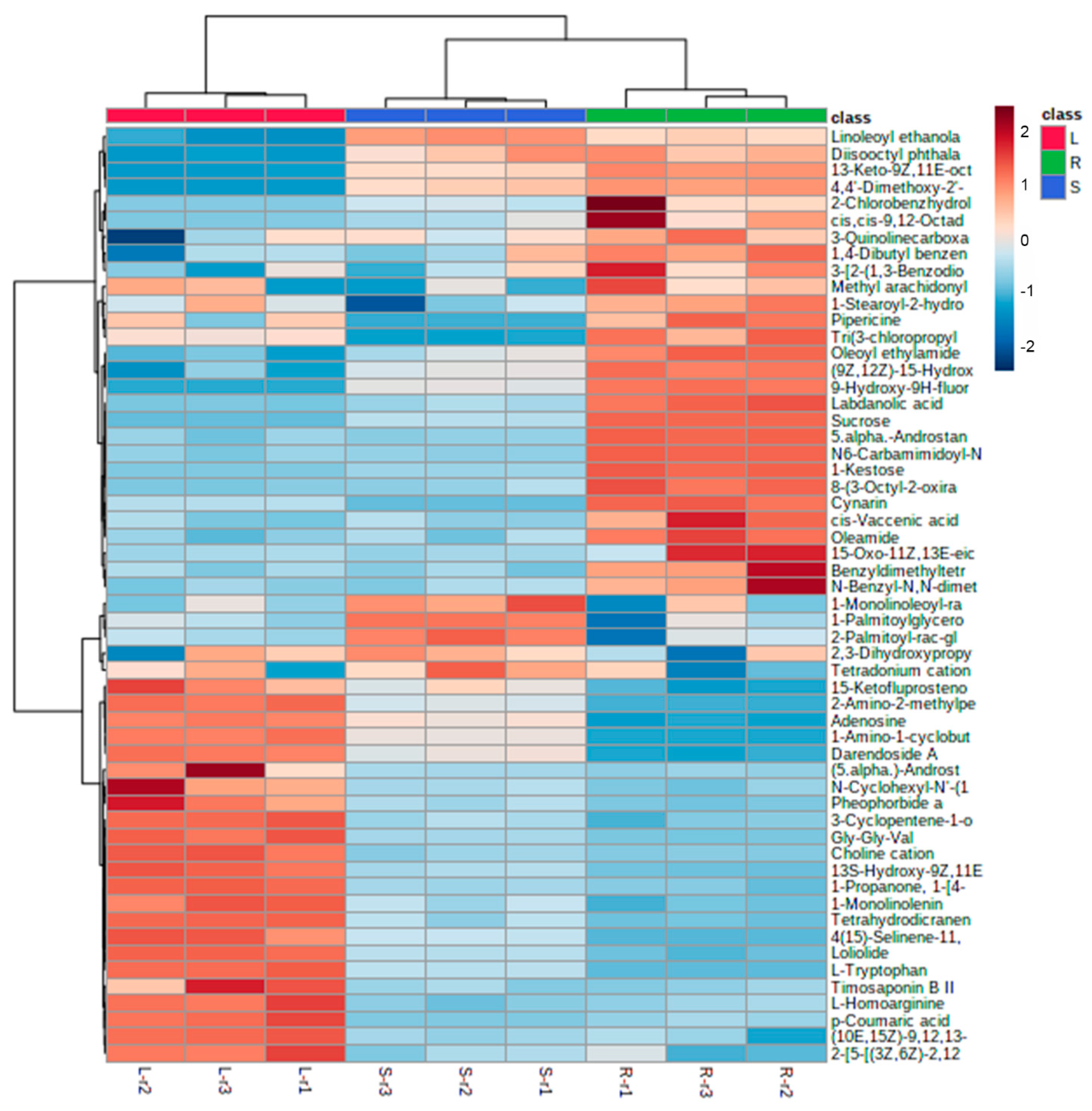
3.1. Untargeted LC-MS/MS-Based Metabolomics
3.2. Antimicrobial Effects
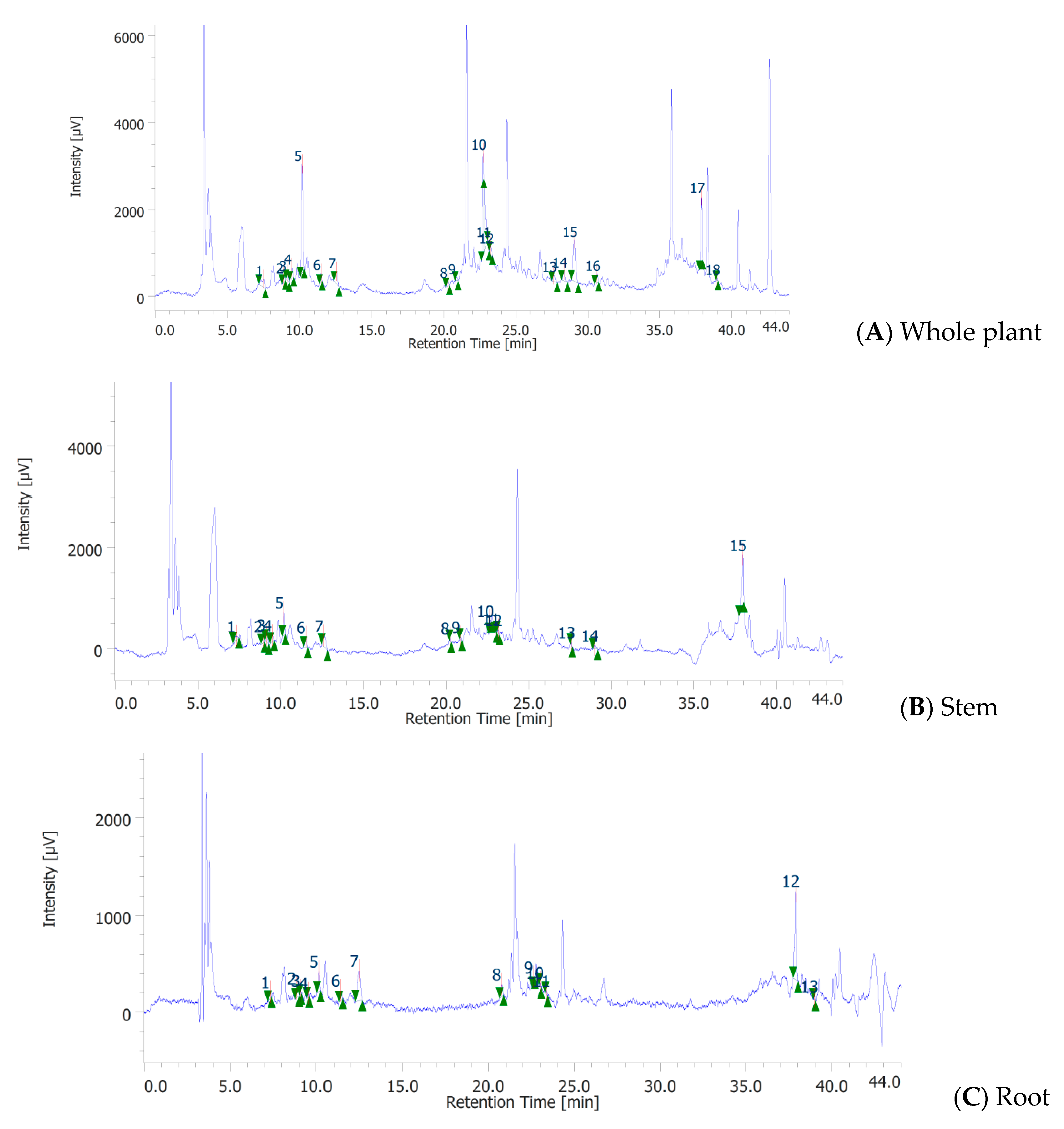
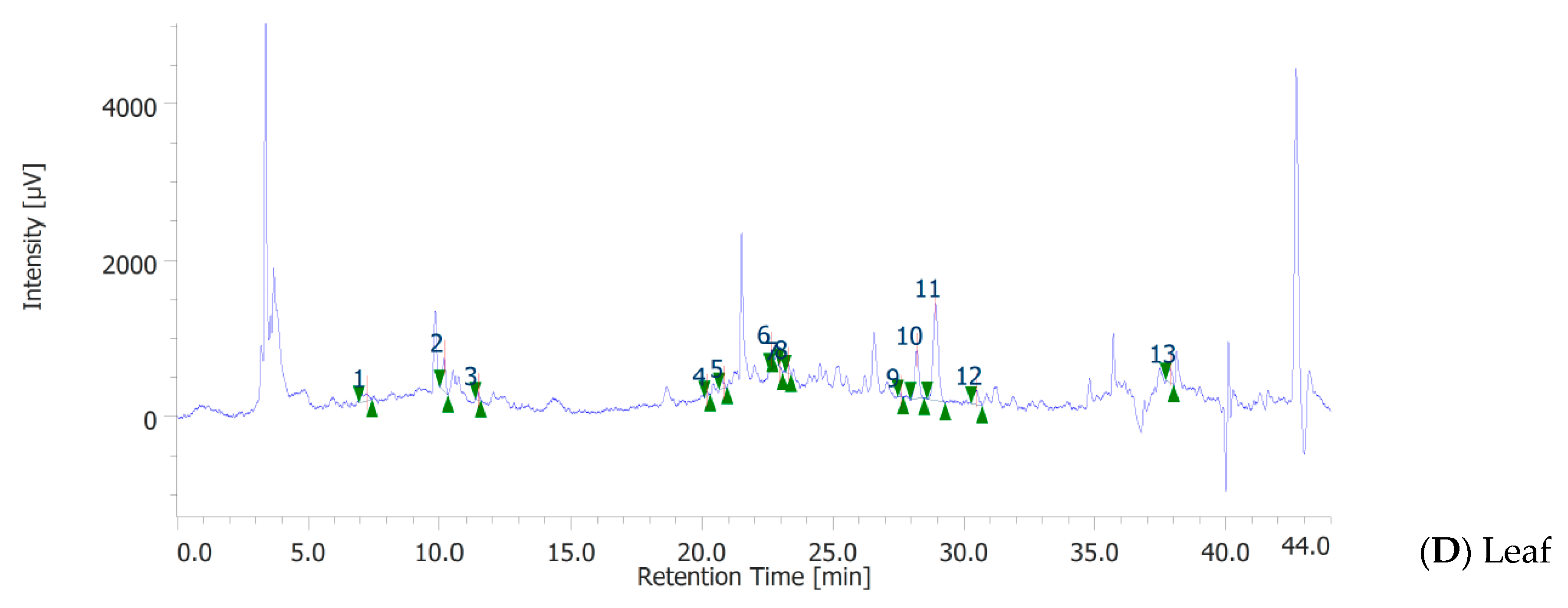
3.3. Phenolic Profile
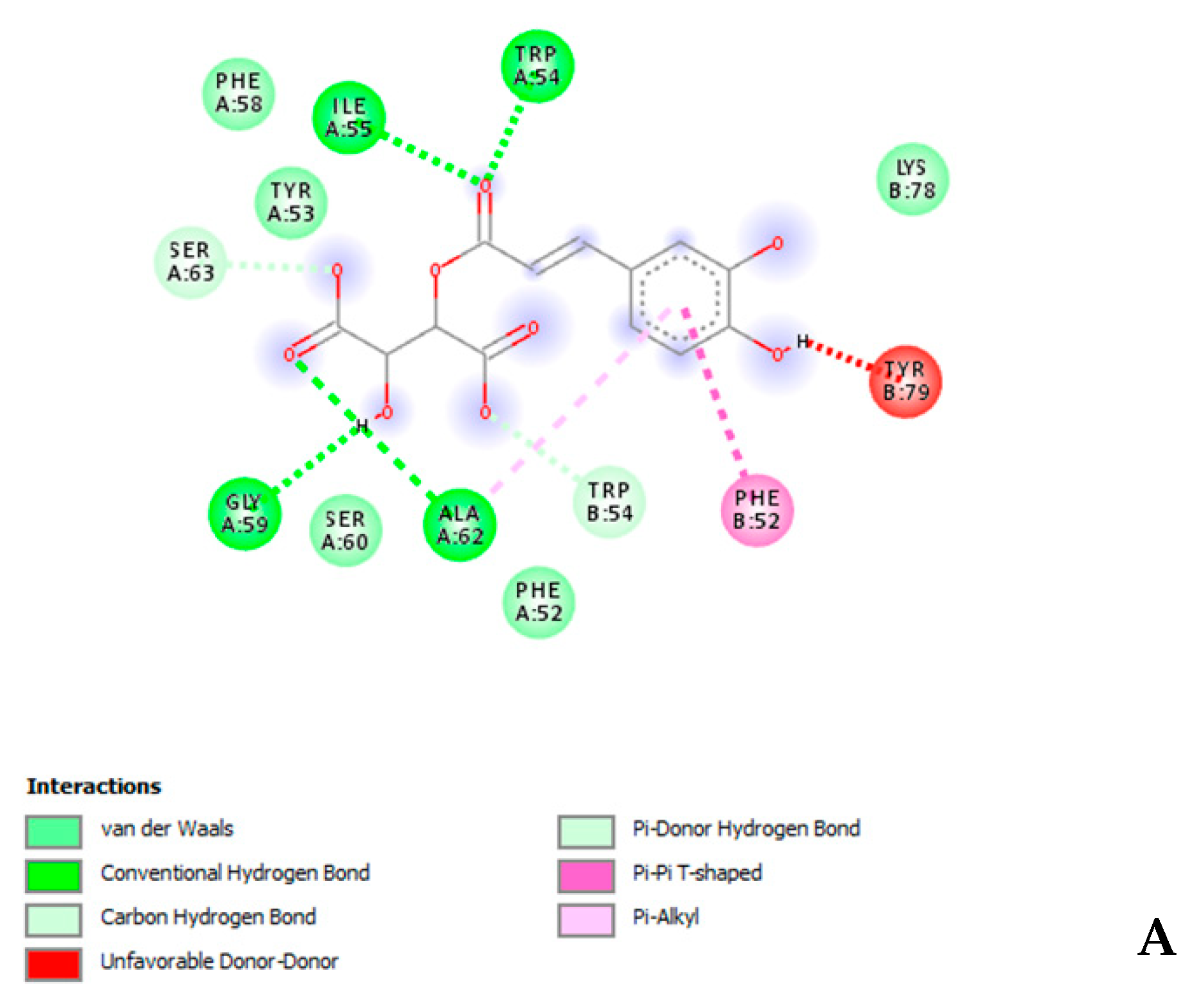
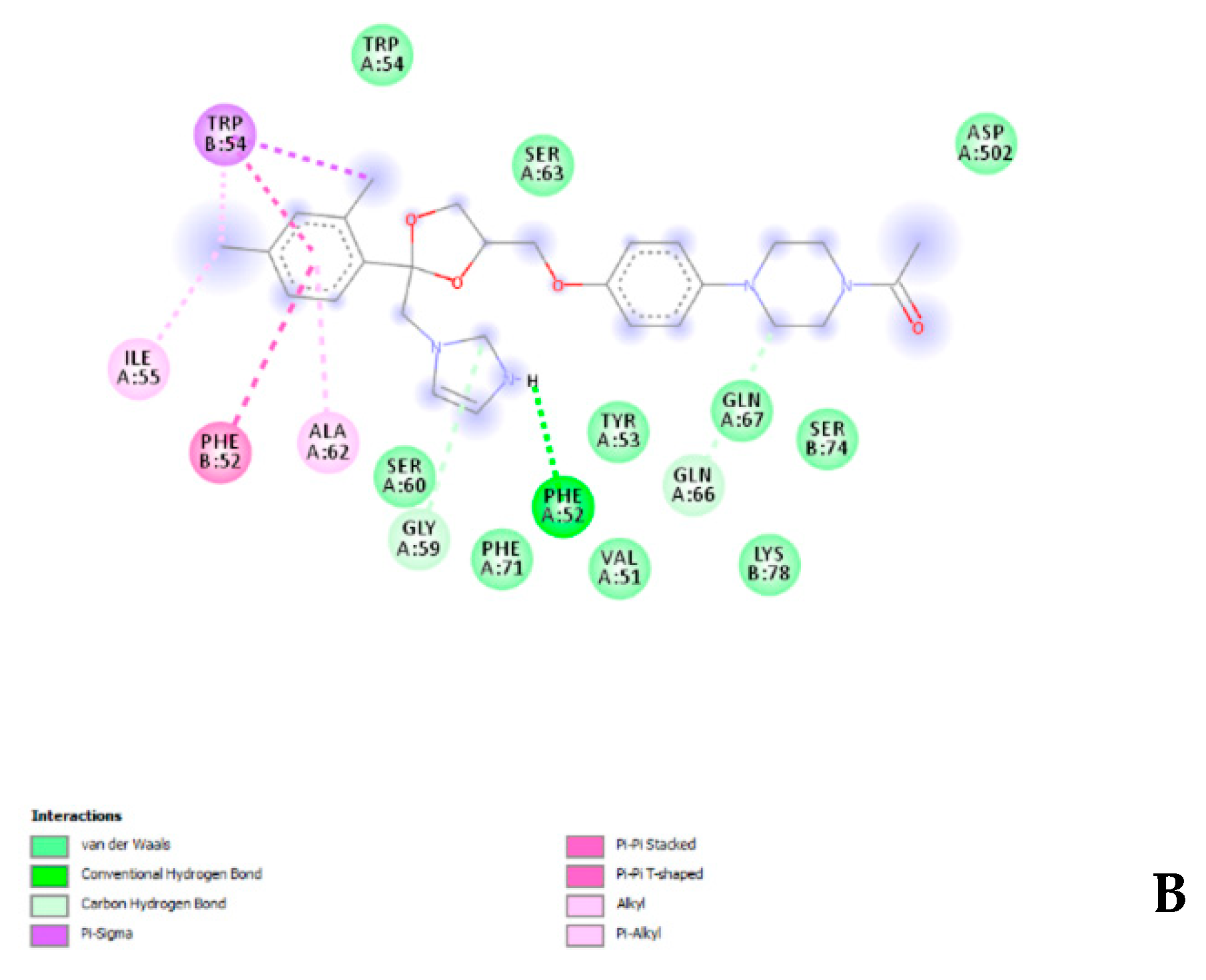
3.4. In Silico Experiments
3.5. Intrinsic Scavenging/Reducing Properties
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferrante, C.; Angelini, P.; Venanzoni, R.; Angeles Flores, G.; Tirillini, B.; Recinella, L.; Chiavaroli, A.; Brunetti, L.; Leone, S.; Di Simone, S.C.; et al. Antimicrobial, antioxidant, and antiproliferative effects of Coronilla minima: An unexplored botanical species. Antibiotics 2020, 9, 611. [Google Scholar] [CrossRef]
- Mahomoodally, M.F.; Jugreet, S.; Sinan, K.I.; Zengin, G.; Ak, G.; Ceylan, R.; Jekő, J.; Cziáky, Z.; Angelini, P.; Angeles Flores, G.; et al. Pharmacological potential and chemical characterization of Bridelia ferruginea Benth. A native tropical African medicinal plant. Antibiotics 2021, 10, 223. [Google Scholar] [CrossRef]
- Chiavaroli, C.; Sinan, K.I.; Zengin, G.; Mahomoodally, M.F.; Sadeer, N.B.; Etienne, O.K.; Cziáky, Z.; Jeko, J.; Glamocilja, J.; Sokovic, M.; et al. Identification of chemical profiles and biological properties of Rhizophora racemosa G. Mey. extracts obtained by different methods and solvents. Antioxidants 2020, 9, 533. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, C.; Chiavaroli, A.; Angelini, P.; Venanzoni, R.; Angeles Flores, G.; Brunetti, L.; Petrucci, M.; Politi, M.; Menghini, L.; Leone, S.; et al. Phenolic content and antimicrobial and anti-Inflammatory effects of Solidago virga-aurea, Phyllanthus niruri, Epilobium angustifolium, Peumus boldus, and Ononis spinosa extracts. Antibiotics 2020, 9, 783. [Google Scholar] [CrossRef] [PubMed]
- Xuan, T.D.; Khanh, T.D. Chemistry and pharmacology of Bidens pilosa: An overview. J. Pharm. Investig. 2016, 46, 91–132. [Google Scholar] [CrossRef] [PubMed]
- Bartolome, A.P.; Villaseñor, I.M.; Yang, W.C. Bidens pilosa L. (Asteraceae): Botanical properties, traditional uses, phytochemistry, and pharmacology. Evid. Based Complement. Alternat. Med. 2013, 2013, 340215. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.L.; Chen, S.C.; Chang, N.W.; Chang, J.M.; Lee, M.L.; Tsai, P.C.; Fu, H.H.; Kao, W.W.; Chiang, H.C.; Wang, H.H.; et al. Protection from oxidative damage using Bidens pilosa extracts in normal human erythrocytes. Food Chem. Toxicol. 2006, 44, 1513–1521. [Google Scholar] [CrossRef]
- Brandão, M.G.; Krettli, A.U.; Soares, L.S.; Nery, C.G.; Marinuzzi, H.C. Antimalarial activity of extracts and fractions from Bidens pilosa and other Bidens species (Asteraceae) correlated with the presence of acetylene and flavonoid compounds. J. Ethnopharmacol. 1997, 57, 131–138. [Google Scholar] [CrossRef]
- Kumari, P.; Misra, K.; Sisodia, B.S.; Faridi, U.; Srivastava, S.; Luqman, S.; Darokar, M.P.; Negi, A.S.; Gupta, M.M.; Singh, S.C.; et al. A promising anticancer and antimalarial component from the leaves of Bidens pilosa. Planta Med. 2009, 75, 59–61. [Google Scholar] [CrossRef] [Green Version]
- Kviecinski, M.R.; Felipe, K.B.; Correia, J.F.; Ferreira, E.A.; Rossi, M.H.; de Moura Gatti, F.; Filho, D.W.; Pedrosa, R.C. Brazilian Bidens pilosa Linné yields fraction containing quercetin-derived flavonoid with free radical scavenger activity and hepatoprotective effects. Libyan J. Med. 2011, 6. [Google Scholar] [CrossRef]
- Ubillas, R.P.; Mendez, C.D.; Jolad, S.D.; Luo, J.; King, S.R.; Carlson, T.J.; Fort, D.M. Antihyperglycemic acetylenic glucosides from Bidens pilosa. Planta Med. 2000, 66, 82–83. [Google Scholar] [CrossRef]
- Geissberger, P.; Séquin, U. Constituents of Bidens pilosa L.: Do the components found so far explain the use of this plant in traditional medicine? Acta Trop. 1991, 48, 251–261. [Google Scholar] [CrossRef]
- Khan, M.R.; Kihara, M.; Omoloso, A.D. Antimicrobial activity of Bidens pilosa, Bischofia javanica, Elmerillia papuana and Sigesbekia orientalis. Fitoterapia 2001, 72, 662–665. [Google Scholar] [CrossRef]
- Motsei, M.L.; Lindsey, K.L.; Van Staden, J.; Jägerm, A.K. Screening of traditionally used South African plants for antifungal activity against Candida albicans. J. Ethnopharmacol. 2003, 86, 235–241. [Google Scholar] [CrossRef]
- Abajo, C.; Boffill, M.A.; Campo, J.D.; Mendez, M.A.; Gonzalez, Y.; Mitjans, M.; Vinardell, M.P. In vitro study of the antioxidant and immunomodulatory activity of aqueous infusion of Bidens pilosa. J. Ethnopharmacol. 2004, 93, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Rojas, J.J.; Ochoa, V.J.; Ocampo, S.A.; Muñoz, J.F. Screening for antimicrobial activity of ten medicinal plants used in Colombian folkloric medicine: A possible alternative in the treatment of non-nosocomial infections. BMC Complement. Altern. Med. 2006, 6, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deba, F.; Xuan, T.D.; Yasuda, M.; Tawata, S. Chemical composition and antioxidant, antibacterial and antifungal activities of the essential oils from Bidens pilosa Linn. var. radiata. Food Control 2008, 19, 346–352. [Google Scholar] [CrossRef]
- Ashafa, A.O.T.; Afolayan, A.J. Screening the root extracts from Biden pilosa L. var. radiata (Asteraceae) for antimicrobial potentials. J. Med. Plant Res. 2009, 3, 568–572. [Google Scholar]
- Silva, J.J.; Cerdeira, C.D.; Chavasco, J.M.; Cintra, A.B.P.; Silva, C.B.P.; Mendonça, A.N.; Ishikawa, T.; Boriollo, M.F.G.; Chavasco, J.K. In vitro screening antibacterial activity of Bidens pilosa Linné and Annona crassiflora Mart. against oxacillin resistant Staphylococcus aureus (ORSA) from the aerial environment at the dental clinic. Rev. Inst. Med. Trop. Sao Paulo 2014, 56, 333–340. [Google Scholar] [CrossRef]
- Shandukani, P.D.; Tshidino, S.C.; Masoko, P.; Moganedi, K.M. Antibacterial activity and in situ efficacy of Bidens pilosa Linn and Dichrostachys cinerea Wight et Arn extracts against common diarrhoea-causing waterborne bacteria. BMC Complement. Altern. Med. 2018, 18, 171. [Google Scholar] [CrossRef]
- Chiang, Y.M.; Chuang, D.Y.; Wang, S.Y.; Kuo, Y.H.; Tsai, P.W.; Shyur, L.F. Metabolite profiling and chemopreventive bioactivity of plant extracts from Bidens pilosa. J. Ethnopharmacol. 2004, 95, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Muchuweti, M.; Mupure, C.; Ndhlala, A.; Murenje, T.; Benhura, M.A.N. Screening of antioxidant and radical scavenging activity of Vigna ungiculata, Bidens pilosa and Cleome gynandra. Am. J. Food Technol. 2007, 2, 161–168. [Google Scholar] [CrossRef]
- Deba, F.; Xuan, T.D.; Yasuda, M.; Tawata, S. Herbicidal and fungicidal activities and identification of potential phytotoxins from Bidens pilosa L. var. radiata Scherff. Weed Biol. Manag. 2007, 7, 77–83. [Google Scholar] [CrossRef]
- Nthulane, N.P.; Mosebi, S.; Tshikalange, T.E.; Nyila, M.A.; Mankga, L.T. Antimicrobial and anti-inflammatory activities of selected medicinal plants against pathogens causing sexually transmitted infections. J. Herbmed. Pharmacol. 2020, 9, 130–137. [Google Scholar] [CrossRef]
- Di Giacomo, V.; Recinella, L.; Chiavaroli, A.; Orlando, G.; Cataldi, A.; Rapino, M.; Di Valerio, V.; Politi, M.; Antolini, M.D.; Acquaviva, A.; et al. Metabolomic profile and antioxidant/anti-Inflammatory effects of industrial hemp water extract in fibroblasts, keratinocytes and isolated mouse skin specimens. Antioxidants 2021, 10, 44. [Google Scholar] [CrossRef]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically; Approved Standard, 10th ed.; CLSI document M07-A10; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- Covino, S.; D’Ellena, E.; Tirillini, B.; Angeles Flores, G.; Arcangeli, A.; Bistocchi, G.; Venanzoni, R.; Angelini, P. Characterization of biological activities of methanol extract of Fuscoporia torulosa (Basidiomycetes) from Italy. Int. J. Med. Mushrooms 2019, 21, 1051–1063. [Google Scholar] [CrossRef]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi, 3rd ed.; CLSI standard M38; Clinical Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts. Approved Standard; Document M38; Clinical Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Pagiotti, R.; Angelini, P.; Rubini, A.; Tirillini, B.; Granetti, B.; Venanzoni, R. Identification and characterisation of human pathogenic filamentous fungi and susceptibility to Thymus schimperi essential oil. Mycoses 2011, 54, e364–e376. [Google Scholar] [CrossRef]
- Angelini, P.; Venanzoni, R.; Angeles Flores, G.; Tirillini, B.; Orlando, G.; Recinella, L.; Chiavaroli, A.; Brunetti, L.; Leone, S.; Di Simone, S.C.; et al. Evaluation of antioxidant, antimicrobial and tyrosinase inhibitory activities of extracts from Tricholosporum goniospermum, an edible wild mushroom. Antibiotics 2020, 9, 513. [Google Scholar] [CrossRef]
- Ferrante, C.; Recinella, L.; Ronci, M.; Menghini, L.; Brunetti, L.; Chiavaroli, A.; Leone, S.; Di Iorio, L.; Carradori, S.; Tirillini, B.; et al. Multiple pharmacognostic characterization on hemp commercial cultivars: Focus on inflorescence water extract activity. Food Chem. Toxicol. 2019, 125, 452–461. [Google Scholar] [CrossRef]
- Bottari, N.B.; Lopes, L.Q.; Pizzuti, K.; Filippi Dos Santos Alves, C.; Corrêa, M.S.; Bolzan, L.P.; Zago, A.; de Almeida Vaucher, R.; Boligon, A.A.; Giongo, J.L.; et al. Antimicrobial activity and phytochemical characterization of Carya illinoensis. Microb. Pathog. 2017, 104, 190–195. [Google Scholar] [CrossRef]
- Waidyanatha, S.; Pierfelice, J.; Cristy, T.; Mutlu, E.; Burback, B.; Rider, C.V.; Ryan, K. A strategy for test article selection and phytochemical characterization of Echinacea purpurea extract for safety testing. Food Chem. Toxicol. 2020, 137, 111125. [Google Scholar] [CrossRef]
- Ramabulana, A.-T.; Steenkamp, P.A.; Madala, N.E.; Dubery, I.A. Application of plant growth regulators modulates the profile of chlorogenic acids in cultured Bidens pilosa cells. Plants 2021, 10, 437. [Google Scholar] [CrossRef]
- Śliwa, K.; Sikora, E.; Ogonowski, J.; Oszmiański, J.; Kolniak-Ostek, J. A micelle mediated extraction as a new method of obtaining the infusion of Bidens tripartita. Acta Biochim. Pol. 2016, 63, 543–548. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef]
- Gülçin, I.; Topal, F.; Çakmakçı, R.; Bilsel, M.; Gören, A.C.; Erdogan, U. Pomological features, nutritional quality, polyphenol content analysis, and antioxidant properties of domesticated and 3 wild ecotype forms of raspberries (Rubus idaeus L.). J. Food Sci. 2011, 76, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Naczk, M.; Shahidi, F. Phenolics in cereals, fruits and vegetables: Occurrence, extraction and analysis. J. Pharm. Biomed. Anal. 2006, 41, 1523–1542, Erratum in: 2007, 43, 798. [Google Scholar] [CrossRef]
- Chatatikun, M.; Supjaroen, P.; Promlat, P.; Chantarangkul, C.; Waranuntakul, S.; Nawarat, J.; Tangpong, J.; Chiabchalard, A. Antioxidant and tyrosinase inhibitor properties of an aqueous extract of Garcinia atrovirdis Griff. Ex. T. Anderson fruit pericarps. Pharmacogn. J. 2020, 12, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Angelini, P.; Tirillini, B.; Bistocchi, G.; Arcangeli, A.; Rubini, A.; Pellegrino, R.M.; Fabiani, R.; Cruciani, G.; Venanzoni, R.; Rosignoli, P. Overview of the biological activities of a methanol extract from wild red belt conk, Fomitopsis pinicola (Agaricomycetes), fruiting bodies from Central Italy. Int. J. Med. Mushrooms 2018, 20, 1047–1063. [Google Scholar] [CrossRef]
- Tanyeli, A.; Ekinci Akdemir, F.N.; Eraslan, E.; Güler, M.C.; Nacar, T. Anti-oxidant and anti-inflamatuar effectiveness of caftaric acid on gastric ulcer induced by indomethacin in rats. Gen. Physiol Biophys. 2019, 38, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, N.; Kanekura, T.; Higashi, Y.; Kanzaki, T. Bidens pilosa suppresses interleukin-1beta-induced cyclooxygenase-2 expression through the inhibition of mitogen activated protein kinases phosphorylation in normal human dermal fibroblasts. J. Dermatol. 2006, 33, 676–683. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | S-r1 | S-r2 | S-r3 | R-r1 | R-r2 | R-r3 | L-r1 | L-r2 | L-r3 | SRL-r1 | SRL-r2 | SRL-r3 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Label | Stem | Stem | Stem | Root | Root | Root | Leaf | Leaf | Leaf | Whole Plant | Whole Plant | Whole Plant |
| (10E,15Z)-9,12,13-Trihydroxyoctadeca-10,15-dienoic acid | 102,173 | 96,783 | 102,660 | 107,873 | 71,433 | 99,821 | 196,627 | 188,188 | 189,856 | 321,988 | 331,819 | 318,475 |
| (5.alpha.)-Androstane-3,11,17-trione | 34,194 | 36,441 | 44,320 | 0 | 8046 | 21,702 | 213,508 | 389,303 | 663,509 | 242,485 | 211,420 | 272,362 |
| (9Z,12Z)-15-Hydroxyoctadeca-9,12-dienoic acid | 1065,623 | 105,5848 | 998,072 | 1,701,017 | 1,663,171 | 1,620,107 | 511,408 | 411,803 | 781,119 | 772,966 | 691,404 | 827,335 |
| 1-Amino-1-cyclobutanecarboxylic acid | 2669,482 | 2,636,512 | 2,660,328 | 143,652 | 137,926 | 133,015 | 5,333,338 | 5,134,590 | 5,057,910 | 1,530,695 | 1,489,768 | 1,507,354 |
| 1-Kestose | 38,408 | 36,278 | 38,266 | 405,333 | 395,093 | 385,401 | 2708 | 2676 | 3990 | 95,238 | 96,636 | 93,696 |
| 1-Monolinolenin | 117,654 | 76,238 | 115,568 | 9897 | 37,743 | 42,994 | 359,371 | 313,651 | 367,023 | 181,659 | 181,033 | 124,774 |
| 1-Monolinoleoyl-rac-glycerol | 272,465 | 212,759 | 229,670 | 20,577 | 76,455 | 187,741 | 91,642 | 76,617 | 145,129 | 62,188 | 69,493 | 58,052 |
| 1-Palmitoylglycerol | 1181,775 | 1,196,756 | 1,197,374 | 793,127 | 958,065 | 1,029,851 | 938,824 | 1,007,960 | 980,759 | 882,266 | 887,517 | 894,507 |
| 1-Propanone,1-[4-(5′-chloro-3,5-dimethyl [2,4′-bipyridin]-2′-yl)-1-piperazinyl]-3-(methylsulfonyl)- | 27,306 | 25,892 | 27,243 | 18,541 | 9878 | 18,272 | 113,474 | 115,616 | 117,522 | 131,924 | 121,542 | 121,718 |
| 1-Stearoyl-2-hydroxy-sn-glycero-3-phosphate | 97,333 | 93,382 | 83,501 | 104,747 | 108,420 | 105,850 | 98,269 | 97,664 | 105,060 | 99,102 | 95,465 | 98,623 |
| 1,4-Dibutylbenzene-1,4-dicarboxylate | 416,022 | 371,401 | 360,682 | 434,349 | 440,723 | 423,912 | 375,740 | 326,158 | 374,673 | 416,988 | 433,662 | 403,848 |
| 13-Keto-9Z,11E-octadecadienoic acid | 849,783 | 806,623 | 815,040 | 1,095,797 | 1,059,482 | 1,052,804 | 290,096 | 265,444 | 273,350 | 423,990 | 445,814 | 390,688 |
| 13S-Hydroxy-9Z,11E,15Z-octadecatrienoic acid | 552,453 | 537,008 | 496,079 | 325,373 | 308,474 | 310,760 | 1,425,663 | 1,557,364 | 1,508,194 | 933,262 | 825,946 | 908,150 |
| 15-Ketofluprostenol isopropyl ester | 84,501 | 89,978 | 82,093 | 69,068 | 65,897 | 63,703 | 94,096 | 109,892 | 101,781 | 98,409 | 105,042 | 104,047 |
| 15-Oxo-11Z,13E-eicosadienoic acid | 25,400 | 30,232 | 15,037 | 65,219 | 376,202 | 368,532 | 37,077 | 23,581 | 35,402 | 40,086 | 30,585 | 18,520 |
| 2-[5-[(3Z,6Z)-2,12-Dihydroxydodeca-3,6-dienyl]oxolan-2-yl]acetic acid | 68,976 | 68,620 | 56,467 | 81,527 | 49,314 | 45,317 | 150,965 | 134,456 | 132,756 | 97,041 | 95,508 | 89,950 |
| 2-Amino-2-methylpentanoic acid | 1,520,393 | 1,553,978 | 1,496,055 | 827,158 | 798,054 | 810,260 | 2,691,101 | 2,655,615 | 2,582,067 | 1,721,792 | 1,718,619 | 1,695,509 |
| 2-Chlorobenzhydrol | 47,514 | 57,401 | 55,174 | 232,441 | 90,092 | 86,206 | 21,397 | 22,254 | 21,276 | 36,777 | 33,981 | 37,482 |
| 2-Palmitoyl-rac-glycerol | 987,286 | 1,009,506 | 985,447 | 728,865 | 862,561 | 876,106 | 834,020 | 857,752 | 840,223 | 732,102 | 749,222 | 776,668 |
| 2,3-Dihydroxypropyl octadecanoate | 1,133,703 | 1,196,710 | 1,244,961 | 1,030,349 | 1,164,072 | 835,097 | 1,154,587 | 868,611 | 1,207,623 | 843,702 | 579,951 | 779,486 |
| 3-[2-(1,3-Benzodioxol-5-yl)-2-oxoethyl]-5-bromo-3-hydroxy-1,3-dihydro-2H-indol-2-one | 227,015 | 215,612 | 203,206 | 252,015 | 239,255 | 225,506 | 221,613 | 209,372 | 200,299 | 250,098 | 254,710 | 238,036 |
| 3-Cyclopentene-1-octanoic acid, 2-(3-hydroxy-1-penten-1-yl)-5-oxo- | 100,681 | 107,678 | 93,923 | 62,210 | 85,529 | 82,842 | 240,640 | 231,413 | 230,760 | 209,629 | 175,091 | 174,184 |
| 3-Quinolinecarboxamide, 1,4-dihydro-6-(1-methylethyl)-4-oxo-1-pentyl-N-tricyclo[3.3.1.1(3,7)]dec-1-yl- | 836,070 | 736,462 | 839,506 | 978,107 | 896,483 | 1,084,069 | 843,779 | 273,146 | 671,986 | 886,706 | 548,460 | 734,240 |
| 4,4′-Dimethoxy-2′-hydroxychalcone | 643,936 | 619,740 | 577,619 | 751,830 | 751,240 | 727,338 | 198,064 | 189,351 | 197,834 | 706,887 | 682,076 | 696,318 |
| 4(15)-Selinene-11,12-diol | 60,495 | 62,398 | 61,881 | 6961 | 7336 | 5863 | 168,662 | 207,043 | 204,807 | 105,752 | 118,421 | 81,926 |
| 5.alpha.-Androstane-3,17-dione | 232,166 | 218,264 | 177,346 | 1,647,262 | 1,626,859 | 1,602,939 | 271,209 | 250,080 | 75,003 | 132,367 | 126,227 | 105,284 |
| 8-(3-Octyl-2-oxiranyl)octanoic acid | 343,285 | 201,933 | 158,994 | 1,477,105 | 1,379,746 | 1,304,038 | 155,294 | 102,545 | 94,800 | 94,103 | 87,381 | 124,007 |
| 9-Hydroxy-9H-fluorene-9-carboxylic acid ethyl ester | 121,171 | 126,353 | 124,089 | 272,125 | 268,880 | 279,196 | 817 | 299 | 740 | 82,251 | 82,078 | 82,875 |
| Adenosine | 124,413 | 120,181 | 125,404 | 56,006 | 59,374 | 61,803 | 171,186 | 172,298 | 175,864 | 93,050 | 87,825 | 92,119 |
| Benzyldimethyltetradecylammonium cation | 3,751,915 | 7,049,053 | 4,888,025 | 2 × 107 | 3.1 × 107 | 2 × 107 | 7,066,741 | 7,550,522 | 4,195,071 | 3,096,660 | 2,012,705 | 5,112,836 |
| Choline cation | 42,960 | 41,793 | 28,619 | 27,102 | 26,229 | 30,563 | 184,548 | 204,572 | 206,850 | 29,087 | 31,226 | 49,225 |
| cis-Vaccenic acid | 297,462 | 296,264 | 468,595 | 1,128,680 | 1,491,160 | 1,804,998 | 212,669 | 448,366 | 225,255 | 100,788 | 126,485 | 163,398 |
| cis,cis-9,12-Octadecadien-1-ol | 3,955,832 | 2,099,838 | 1,737,622 | 1.5 × 107 | 8,594,468 | 5,123,672 | 710,151 | 474,279 | 560,286 | 667,796 | 763,629 | 368,557 |
| Cynarin | 5467 | 4750 | 4372 | 124,950 | 118,904 | 129,559 | 31,993 | 31,242 | 31,062 | 39,844 | 36,948 | 40,683 |
| Darendoside A | 246,997 | 243,141 | 234,194 | 156,516 | 162,866 | 154,800 | 322,945 | 334,038 | 329,086 | 301,644 | 294,323 | 305,039 |
| Diisooctyl phthalate | 655,023 | 539,591 | 460,410 | 662,511 | 587,209 | 534,374 | 130,106 | 129,056 | 145,171 | 125,839 | 114,733 | 113,067 |
| Gly-Gly-Val | 590,655 | 579,052 | 581,744 | 340,701 | 347,372 | 323,871 | 2,281,278 | 2,223,467 | 2,070,998 | 917,095 | 867,045 | 896,749 |
| L-Homoarginine | 29,419 | 8015 | 33,727 | 35,426 | 56,567 | 50,880 | 321,122 | 273,969 | 265,960 | 115,573 | 142,021 | 120,612 |
| L-Tryptophan | 109,556 | 107,202 | 110,760 | 41,130 | 40,510 | 41,398 | 320,162 | 308,335 | 307,264 | 84,341 | 88,041 | 87,022 |
| Labdanolic acid | 47,859 | 59,733 | 37,730 | 258,351 | 293,240 | 278,953 | 12,472 | 13,866 | 16,874 | 26,643 | 21,161 | 42,621 |
| Linoleoyl ethanolamide | 151,265 | 152,477 | 146,410 | 116,457 | 116,544 | 123,219 | 32,107 | 46,621 | 33,168 | 77,323 | 94,694 | 68,613 |
| Loliolide | 141,912 | 139,791 | 146,221 | 60,562 | 57,027 | 35,675 | 435,314 | 448,573 | 448,272 | 274,405 | 254,800 | 263,034 |
| Methyl arachidonyl fluorophosphonate | 185,247 | 192,752 | 183,865 | 204,057 | 196,979 | 194,467 | 183,821 | 198,621 | 197,452 | 182,045 | 181,197 | 178,476 |
| N-Benzyl-N,N-dimethyl-1-hexadecanaminium cation | 3,372,469 | 3,200,520 | 1,997,818 | 7,575,669 | 1.3 × 107 | 8,243,737 | 2,263,083 | 1,815,563 | 2,871,648 | 375,748 | 582,177 | 882,638 |
| N-Cyclohexyl-N′-(1-naphthyl)urea | 84,932 | 71,905 | 69,777 | 40,831 | 60,361 | 32,076 | 214,537 | 368,038 | 224,414 | 168,117 | 127,194 | 170,491 |
| N6-Carbamimidoyl-N2-((4E,6E,12E,14E)-3-hydroxy-2-(hydroxymethyl)-8,10,16-trimethyloctadeca-4,6,12,14-tetraenoyl)lysine | 224,512 | 207,881 | 265,592 | 2,147,753 | 2,106,907 | 2,110,218 | 306,836 | 266,825 | 97,710 | 189,847 | 148,730 | 175,346 |
| Oleamide | 187,073 | 148,732 | 182,692 | 312,739 | 319,209 | 347,815 | 163,145 | 170,533 | 141,877 | 212,284 | 217,719 | 192,874 |
| Oleoyl ethylamide | 288,452 | 278,168 | 247,724 | 381,802 | 404,893 | 409,015 | 181,339 | 206,324 | 223,119 | 866,575 | 626,771 | 825,341 |
| p-Coumaric acid | 642 | 671 | 827 | 6247 | 7505 | 10,671 | 95,179 | 81,212 | 83,525 | 48,922 | 44,264 | 39,818 |
| Pheophorbide a | 882,828 | 636,397 | 802,593 | 315,815 | 317,935 | 237,413 | 2,603,775 | 4,186,462 | 3,212,948 | 1,264,546 | 1,153,771 | 1,085,994 |
| Pipericine | 140,872 | 139,382 | 135,285 | 305,766 | 368,281 | 383,514 | 291,684 | 296,772 | 169,931 | 1,184,817 | 1,198,274 | 1,130,158 |
| Sucrose | 171,039 | 175,897 | 181,013 | 664,452 | 656,404 | 655,640 | 30,340 | 28,754 | 32,234 | 254,888 | 246,343 | 248,963 |
| Tetrahydrodicranenone B | 62,007 | 30,813 | 63,402 | 14,760 | 12,860 | 18,913 | 224,458 | 221,431 | 222,159 | 79,141 | 109,472 | 75,175 |
| Timosaponin B II | 4125 | 33,780 | 20,202 | 4324 | 25,703 | 12,869 | 243,569 | 138,488 | 282,852 | 32,716 | 23,914 | 11,535 |
| Tri(3-chloropropyl) phosphate | 160,488 | 159,000 | 156,995 | 329,548 | 338,360 | 288,679 | 255,460 | 253,231 | 247,410 | 340,901 | 345,598 | 352,026 |
| f.Value | p.Value | log10(p) | FDR | Fisher’s LSD | |
|---|---|---|---|---|---|
| amino acid | 810.05 | 2.8488 × 10−10 | 9.5453 | 9.6858 × 10−9 | Leaf—Plant; Leaf—Root; Leaf—Stem; Plant—Root; Stem—Plant; Stem—Root |
| diterpenoid | 483.51 | 2.2226 × 10−9 | 8.6531 | 3.3113 × 10−8 | Root—Leaf; Stem—Leaf; Root—Plant; Stem—Plant; Root—Stem |
| oligosaccharide | 420.24 | 3.8807 × 10−9 | 8.4111 | 3.3113 × 10−8 | Plant—Leaf; Root—Leaf; Stem—Leaf; Root—Plant; Plant—Stem; Root—Stem |
| disaccharide | 419.83 | 3.8957 × 10−9 | 8.4094 | 3.3113 × 10−8 | Plant—Leaf; Root—Leaf; Stem—Leaf; Root—Plant; Root—Stem |
| cinnamate derivative | 387.96 | 5.3299 × 10−9 | 8.2733 | 3.6244 × 10−8 | Root—Leaf; Leaf—Stem; Root—Plant; Plant—Stem; Root—Stem |
| peptide | 339.51 | 9.0488 × 10−9 | 8.0434 | 4.4266 × 10−8 | Leaf—Plant; Leaf—Root; Leaf—Stem; Plant—Root; Stem—Root |
| benzofurano | 333.66 | 9.6956 × 10−9 | 8.0134 | 4.4266 × 10−8 | Leaf—Plant; Leaf—Root; Leaf—Stem; Plant—Root; Plant—Stem; Stem—Root |
| hydroxycinnamic acid | 327.68 | 1.0416 × 10−8 | 7.9823 | 4.4266 × 10−8 | Leaf—Plant; Leaf—Root; Leaf—Stem; Plant—Root; Plant—Stem |
| nucleoside | 283.3 | 1.8539 × 10−8 | 7.7319 | 7.0037 × 10−8 | Leaf—Plant; Leaf—Root; Leaf—Stem; Plant—Root; Stem—Plant; Stem—Root |
| chalcone derivate | 208.61 | 6.2137 × 10−8 | 7.2067 | 2008 × 10−4 | Plant—Leaf; Root—Leaf; Stem—Leaf; Stem—Plant; Stem—Root |
| amine | 206.27 | 6.4966 × 10−8 | 7.1873 | 2.008 × 10−4 | Leaf—Plant; Leaf—Root; Leaf—Stem; Plant—Root; Stem—Plant; Stem—Root |
| fatty acid | 176.56 | 1.199 × 10−4 | 6.9212 | 3.3971 × 10−7 | Leaf—Plant; Root—Leaf; Stem—Leaf; Root—Plant; Stem—Plant; Root—Stem |
| quaternary ammonium compound | 165.01 | 1.5641 × 10−7 | 6.8057 | 4.0907 × 10−7 | Leaf—Plant; Leaf—Root; Leaf—Stem; Stem—Root |
| carbamide | 149.31 | 2.3163 × 10−7 | 6.6352 | 5.6252 × 10−7 | Root—Leaf; Root—Plant; Root—Stem |
| steroid | 128.67 | 4.1484 × 10−7 | 6.3821 | 9.4031 × 10−7 | Leaf—Plant; Root—Leaf; Leaf—Stem; Root—Plant; Root—Stem |
| phenolic derivative | 122.17 | 5.0797 × 10−7 | 6.2942 | 1.0794 × 10−6 | Leaf—Plant; Leaf—Root; Plant—Root; Stem—Root |
| N-acyl amine | 117.35 | 5.9443 × 10−7 | 6.2259 | 1.1889 × 10−6 | Plant—Leaf; Plant—Root; Plant—Stem |
| Cyclopentane | 72.681 | 3.806 × 10−3 | 5.4195 | 7.189 × 10−3 | Leaf—Plant; Leaf—Root; Leaf—Stem; Plant—Root; Stem—Root |
| Monoglyceride | 65.417 | 5.7002 × 10−6 | 5.2441 | 1.02 × 10−5 | Leaf—Plant; Leaf—Root; Stem—Leaf; Stem—Plant; Stem—Root |
| phthalate derivate | 55.477 | 1.0687 × 10−5 | 4.9711 | 1.8168 × 10−5 | Root—Leaf; Stem—Leaf; Root—Plant; Stem—Plant; Stem—Root |
| diterpene | 53.245 | 1.2488 × 10−5 | 4.9035 | 2.0219 × 10−5 | Leaf—Plant; Leaf—Root; Leaf—Stem; Plant—Root; Stem—Root |
| furanone derivate | 27.794 | 0.00013939 | 3.8558 | 0.00021543 | Leaf—Plant; Leaf—Root; Leaf—Stem; Plant—Root; Stem—Root |
| Saponin | 23.322 | 0.00026134 | 3.5828 | 0.00036454 | Leaf—Plant; Leaf—Root; Leaf—Stem |
| porphyrins | 23.314 | 0.00026165 | 3.5823 | 0.00036454 | Leaf—Plant; Leaf—Root; Leaf—Stem |
| Aliphatic ester | 23.156 | 0.00026805 | 3.5718 | 0.00036454 | Leaf—Plant; Stem—Leaf; Stem—Plant; Stem—Root |
| aromatic ester | 21.937 | 0.00032451 | 3.4888 | 0.00042436 | Leaf—Root; Plant—Root; Stem—Root |
| organofluorine derivate | 21.655 | 0.00033966 | 3.469 | 0.00042771 | Root—Leaf; Root—Plant; Stem—Plant; Root—Stem |
| amide | 20.619 | 0.00040342 | 3.3942 | 0.00048987 | Root—Leaf; Root—Plant; Root—Stem |
| glycerophospholipid | 14.443 | 0.0013585 | 2.867 | 0.0015927 | Stem—Leaf; Stem—Plant; Stem—Root |
| terephthalate derivate | 9.818 | 0.0046632 | 2.3313 | 0.0052849 | Stem—Leaf; Stem—Plant; Stem—Root |
| bromo derivate | 8.1485 | 0.008147 | 2.089 | 0.0089355 | Stem—Leaf; Stem—Root |
| aliphatic alcohol | 6.3184 | 0.016665 | 1.7782 | 0.017707 | Root—Leaf; Root—Plant; Root—Stem |
| organochloride compound | 5.4374 | 0.024748 | 1.6065 | 0.025498 | Plant—Stem; Root—Stem |
| Minimum Inhibitory Concentration (MIC) | |||||
|---|---|---|---|---|---|
| Extract Typology | n-hex (mg mL−1) | EtOAc (mg mL−1) | MeOH (mg mL−1) | Ciprofloxacin (µg mL−1) | |
| Bacterial strain | Plant parts | ||||
| E. coli (ATCC 10536) | roots | <0.031 | <0.031 | <0.031 | <0.12 |
| leaves | <0.031 | <0.031 | <0.031 | ||
| stems | <0.031 | <0.031 | <0.031 | ||
| whole | <0.031 | <0.031 | <0.031 | ||
| E. coli (PeruMycA 2) | roots | 0.049 (0.031–0.062) | 0.078 (0.062–0.125) | 0.157 (0.125–0.25) | 1.23 (0.98–1.95) |
| leaves | 0.049 (0.031–0.062) | 0.039 (0.031–0.062) | 0.157 (0.125–0.25) | ||
| stems | 0.039 (0.031–0.62) | 0.157 (0.125–0.25) | 0.157 (0.125–0.25) | ||
| whole | <0.031 | 0.157 (0.125–0.25) | 0.157 (0.125–0.25) | ||
| E. coli (PeruMycA 3) | roots | 0.099 (0.062–0.125) | 0.157 (0.125–0.25) | 0.157 (0.125–0.25) | 0.62 (0.49–0.98) |
| leaves | 0.198 (0.125–0.25) | 0.039 (0.031–0.062) | 0.198 (0.125–0.25) | ||
| stems | 0.078 (0.062–0.125) | 0.315 (0.25–0.5) | 0.157 (0.125–0.25) | ||
| whole | 0.039 (0.031–0.62) | 0.157 (0.125–0.25) | 0.157 (0.125–0.25) | ||
| B. cereus (PeruMycA 4) | roots | <0.031 | <0.031 | <0.031 | <0.12 |
| leaves | <0.031 | <0.031 | <0.031 | ||
| stems | <0.031 | <0.031 | <0.031 | ||
| whole | <0.031 | <0.031 | <0.031 | ||
| P. aeruginosa (ATCC15442) | roots | 0.039 (0.031–0.062) | 0.078 (0.062–0.125) | <0.031 | <0.12 |
| leaves | 0.049 (0.031–0.062) | 0.039 (0.031–0.062) | <0.031 | ||
| stems | 0.039 (0.031–0.062) | 0.039 (0.031–0.062) | <0.031 | ||
| whole | 0.039 (0.031–0.62) | 0.078 (0.062–0.125) | <0.031 | ||
| B. subtilis (PeruMyc 6) | roots | 0.315 (0.5–0.25) | 0.198 (0.125–0.25) | 0.078 (0.062–0.125) | <0.12 |
| leaves | 0.396 (0.25–0.5) | 0.396 (0.25–05) | 0.078 (0.062–0.125) | ||
| stems | 0.315 (0.5–0.25) | 0.198 (0.125–0.25) | <0.031 | ||
| whole | 0.315 (0.25–0.5) | 0.098 (0.062–0.125) | <0.031 | ||
| S. typhy (PeruMyc 7) | roots | 0.049 (0.031–0.062) | 0.157 (0.125–0.25) | 0.157 (0.125.0.25) | 0.49 |
| leaves | 0.198 (0.125–0.25) | 0.198 (0.125–0.25) | 0.078 (0.062–0.125) | ||
| stems | 0.198 (0.125–0.25) | 0.039 (0.031–0.062) | 0.049 (0.031–0.062) | ||
| whole | 0.157 (0.125–0.25) | 0.198 (0.125–0.25) | 0.049 (0.031–0.062) | ||
| S. aureus (ATCC 6538) | roots | 0.039 (0.031–0.062) | 0.039 (0.031–0.062) | 0.039 (0.031–0.062) | 0.98 |
| leaves | 0.049 (0.031–0.062) | 0.039 (0.031–0.062) | 0.039 (0.031–0.062) | ||
| stems | 0.039 (0.031–0.062) | 0.039 (0.031–0.062) | 0.039 (0.031–0.062) | ||
| whole | 0.039 (0.031–0.062) | 0.039 (0.031–0.062) | 0.039 (0.031–0.062) | ||
| Extract Typology | Minimum Inhibitory Concentration (MIC) | ||||
|---|---|---|---|---|---|
| n-hex (mg mL−1) | EtOAc (mg mL−1) | MeOH (mg mL−1) | Fluconazole (µg mL−1) | ||
| Yeast strain | Plant parts | ||||
| C. albicans (YEPGA 6183) | roots | 0.39 (0.25–0.5) | 0.198 (0.125–0.250) | 0.157 (0.125–0.250) | 2 |
| leaves | 0.314 (0.125–0.25) | 0.315 (0.25–0.5) | 0.198 (0.125–0.250) | ||
| stems | 0.396 (0.25–0.5) | 0.315 (0.25–0.5) | 0.396 (0.25–0.5) | ||
| whole | 0.198 (0.125–025) | 0.315 (0.25–0.5) | 0.315 (0.25–0.5) | ||
| C. tropicalis (YEPGA 6184) | roots | 0.051 (0.031–0.065) | 0.099 (0.0625–0.125) | <0.031 | 2 |
| leaves | 0.314 (0.25–0.5) | 0.157 (0.125–0.250) | 0.198 (0.125–0.250) | ||
| stems | 0.198 (0.125–0.25) | 0.198 (0.125–0.250) | 0.099 (0.0625–0.125) | ||
| whole | 0.198 (0.125–0.5) | 0.099 (0.0625–0.125) | 0.198 (0.125–0.250) | ||
| C. albicans (YEPGA 6379) | roots | 0.314 (0.25–0.5) | 0.099 (0.0625–0.125) | 0.157 (0.125–0.250) | 1 |
| leaves | 0.396 (0.25–0.5) | 0.198 (0.125–0.250) | 0.198 (0.125–0.250) | ||
| stems | 0.314 (0.250–0.5) | 0.315 (0.25–0.5) | 0.396 (0.25–0.5) | ||
| whole | 0.314 (0.125–0.5) | 0.315 (0.25–0.5) | 0.315 (0.25–0.5) | ||
| C. parapsilosis (YEPGA 6551) | roots | <0.031 | <0.031 | <0.031 | 4 |
| leaves | 0.157 (0.125–0.25) | <0.031 | <0.031 | ||
| stems | 0.198 (0.125–025) | <0.031 | <0.031 | ||
| whole | <0.031 | <0.031 | <0.031 | ||
| Extract Typology | Minimum Inhibitory Concentration (MIC) | ||||
|---|---|---|---|---|---|
| n-hx (mg mL−1) | EtOAc (mg mL−1) | MeOH (µg mL−1) | Griseofulvin (µg mL−1) | ||
| Fungal strain | Plant parts | ||||
| T. rubrum (CCF4933) | roots | 0.198 (0.125–0.25) | <0.031 | <0.031 | 1.26 (1–2) |
| leaves | 0.396 (0.25–0.5) | <0.031 | 0.049 (0.031–0.062) | ||
| stems | 0.315 (0.25–0.5) | 0.049 (0.031–0.062) | 0.157 (0.125–0.25) | ||
| whole | 0.396 (0.25–0.5) | <0.031 | <0.031 | ||
| T. mentagrofites (CCF 4823) | roots | 0.198 (0.125–0.25) | <0.031 | <0.031 | 1 |
| leaves | 0.049 (0.031–0.062) | <0.031 | <0.031 | ||
| stems | 0.315 (0.25–0.5) | 0.049 (0.031–0.062) | 0.157 (0.125–0.25) | ||
| whole | 0.396 (0.25–0.5) | <0.031 | <0.031 | ||
| T.rubrum (CCF4879) | roots | 0.198 (0.125–0.25) | <0.031 | <0.031 | 2 |
| leaves | 0.198 (0.125–2.5 | <0.031 | 0.049 (0.031–0.062) | ||
| stems | 0.315 (0.25–0.5) | <0.031 | 0.157 (0.125–0.25) | ||
| whole | 0.396 (0.25–0.5) | <0.031 | <0.031 | ||
| T. tonsurans (CCF4834) | roots | 0.198 (0.125–0.25) | <0.031 | <0.031 | 0.125 |
| leaves | 0.049 (0.031–0.062) | <0.031 | 198 (0.125–25)) | ||
| stems | 0.315 (0.25–0.5) | <0.031 | 0.157 (0.125–0.25) | ||
| whole | 0.396 (0.25–0.5) | <0.031 | <0.031 | ||
| A. crocatum (CCF5300) | roots | 0.198 (0.125–0.25) | <0.031 | <0.031 | >8 |
| leaves | 0.396 (0.25–0.5) | <0.031 | 0.078 (0.062–0.125) | ||
| stems | 0.315 (0.25–0.5) | 0.049 (0.031–0.062) | 0.157 (0.125–0.25) | ||
| whole | 0.396 (0.25–0.5) | <0.031 | <0.031 | ||
| A. gypseum (CCF6261) | roots | 0.315 (0.25–0.5) | <0.031 | <0.031 | 1.587 (1–2) |
| leaves | 0.396 (0.25–0.5) | <0.031 | 0.157 (0.125–0.25) | ||
| stems | 0.198 (0.125–0.25) | 0.315 (0.25–0.5) | 0.078 (0.062–0.125) | ||
| whole | 0.198 (0.125–0.25) | <0.031 | <0.031 | ||
| T. erinacei (CCF5930) | roots | 0.315 (0.25–0.5) | <0.031 | <0.031 | 0.25 |
| leaves | 0.396 (0.25–0.5) | <0.031 | 0.157 (0.125–0.25) | ||
| stems | 0.198 (0.125–0.25) | 0.157 (0.125–0.250) | 0.157 (0.125–0.25) | ||
| whole | 0.198 (0.125–0.25) | <0.031 | <0.031 | ||
| A. quadrifidum (CCF5792) | roots | 0.315 (0.25–0.5) | <0.031 | <0.031 | >8 |
| leaves | 0.396 (0.25–0.5) | <0.031 | 0.157 (0.125–0.25) | ||
| stems | 0.198 (0.125–0.25) | 0.157 (0.125–0.250) | 0.157 (0.125–0.25) | ||
| whole | 0.396 (0.25–0.5) | <0.031 | <0.031 | ||
| Plant Material | DPPH | ABTS | FRAP | β-Carotene/Linoleic Acid Assay |
|---|---|---|---|---|
| roots | 15.2 ± 1.22 | 25.3 ± 2.16 | 15.3 ± 1.17 | 21.1 ± 1.74 |
| stems | 101.4 ± 8.23 | 89.1 ± 7.81 | 10.1 ± 0.87 | 44.7 ± 3.86 |
| leaves | 9.9 ± 0.84 | 15.4 ± 1.36 | 73.2 ± 6.5 | 27.4 ± 2.32 |
| plants | 3.7 ± 2.38 | 34.3 ± 3.05 | 17.7 ± 1.49 | 37.4 ± 3.78 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angelini, P.; Matei, F.; Flores, G.A.; Pellegrino, R.M.; Vuguziga, L.; Venanzoni, R.; Tirillini, B.; Emiliani, C.; Orlando, G.; Menghini, L.; et al. Metabolomic Profiling, Antioxidant and Antimicrobial Activity of Bidens pilosa. Processes 2021, 9, 903. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9060903
Angelini P, Matei F, Flores GA, Pellegrino RM, Vuguziga L, Venanzoni R, Tirillini B, Emiliani C, Orlando G, Menghini L, et al. Metabolomic Profiling, Antioxidant and Antimicrobial Activity of Bidens pilosa. Processes. 2021; 9(6):903. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9060903
Chicago/Turabian StyleAngelini, Paola, Florentina Matei, Giancarlo Angeles Flores, Roberto Maria Pellegrino, Lydie Vuguziga, Roberto Venanzoni, Bruno Tirillini, Carla Emiliani, Giustino Orlando, Luigi Menghini, and et al. 2021. "Metabolomic Profiling, Antioxidant and Antimicrobial Activity of Bidens pilosa" Processes 9, no. 6: 903. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9060903