
Properties of Bacterial Cellulose Produced Using White and Red Grape Bagasse as a Nutrient Source


1
Faculty of Natural Sciences and Engineering, University of Ljubljana, Aškerčeva 12, 1000 Ljubljana, Slovenia
2
Agricultural Institute of Slovenia, Hacquetova ulica 17, 1000 Ljubljana, Slovenia
*
Author to whom correspondence should be addressed.
Processes 2021, 9(7), 1088; https://0-doi-org.brum.beds.ac.uk/10.3390/pr9071088
Submission received: 27 May 2021
/
Revised: 17 June 2021
/
Accepted: 22 June 2021
/
Published: 23 June 2021
(This article belongs to the Special Issue Preparation of Bacterial Cellulose and Its Biomedical Applications)
Abstract
:The purpose of the study is to investigate the possibility of using wine industry wastes, such as red and white grape bagasse, to produce bacterial cellulose (BC) instead of using a costly commercial medium. BC was produced using grape bagasse as a carbon source replacement and the sole nutrient in the medium. The BC films were evaluated for their productivity and water-holding capacity. The BC films were also investigated for their morphology using scanning electron microscopy (SEM), their viscoelastic properties using dynamic mechanical analysis (DMA), and their chemical composition using Fourier-transform infrared spectroscopy (FTIR). Although the use of grape bagasse as the sole nutrient was successful in the preparation of BC, the BC films had inferior viscoelastic properties to other produced BC films. White grape bagasse proved to be an excellent carbon substitute as the production of BC and its water-holding capacity were five times higher and the produced BC films were up to 72% more flexible than the bacterial cellulose produced using standard HS medium.
1. Introduction
Cellulose is a water-insoluble polysaccharide, mainly produced by plants and algae, that can be modified for higher water solubility (i.e., cellulose esters) and employed in numerous medical, environmental, and technological applications [1,2,3]. Cellulose is also produced by bacteria, where Komagatabacter xylinus (formerly named Gluconacetobacter xylinus) is commercially important [4,5]. Nowadays, the production of bacterial cellulose (BC) is increasing since it can be used as a wound dressing and drug delivery material in the production of artificial blood vessels, as scaffolding in tissue engineering, as an additive in healthy food (e.g., Nata-de-coco, fat replacer, artificial meat, rheology modifier), in cosmetics, as a reinforcement of high-quality papers, and for textiles [6,7,8,9]. The most commonly used medium for the growth of BC-producing bacteria is Hestrin-Schramm (HS medium) [10], where glucose is the carbon source, with yeast extract and peptone as sources of nitrogen [11,12]. Due to high production costs when using HS medium, the search for alternative, cheaper carbon and nutrient sources, such as food waste, is a current commercial strategy [12]. This strategy is also interesting from the eco-friendly and circular economy perspective. The majority of the research is focused on using natural carbon sources from fruit waste, i.e., citrus juices from whole oranges and grapefruits (waste from open markets), aqueous extracts from citrus processing waste, [7,13,14,15,16,17], rotten bananas [18], and also a natural nitrogen source from waste beer yeast that has been used instead of the conventional yeast extract used in HS medium [19]. It was found that using grapefruit and orange juices yielded higher bacterial cellulose concentrations than using lemon, aqueous extracts of grapefruit, and orange peels [14]. When comparing natural nutrient sources to HS medium, it was found that production of BC was higher when using pineapple and sugar cane [20], coffee cherry husks [21], potato peels [22], and rotten fruit [23] but lower when using dry olive mill waste [24]. The BC produced on nitric-acid-hydrolyzed potato peel waste had improved properties [22], such as higher crystallinity, higher thermal stability, and higher surface area, than BC produced in HS medium (HS-BC) alone. Similarly, higher crystallinity and higher BC production were obtained from rotten food (RF-BC) but not from the combination of rotten food and milk whey (RFW-BC) [23]. The latter combination, however, did increase the production, but the crystallinity was much lower (i.e., 15.17% to 59.76% for RFW-BC, 64.66% for HB-BC, and 70.69% for RF-BC).
The aim of our research is to evaluate the possibility of replacing the carbon and nutrient sources for BC production from standard HS medium with grape bagasse, a natural waste product from the grape wine industry. According to the United Nations Food and Agriculture Organization (FAO), the global production of grapes increases every year; it was more than 87 million tons in 2017 [25]. Grape bagasse represents almost 20% by weight of total grape input [26]. If the stalks are removed prior to grape processing, the residue consists of 40% seeds and 60% skin and pulp. Winery waste for grapes pressed with the stalks comprises 30% stalks, 30% seeds, and 40% skin and pulp, with slight differences depending on the grape variety. It has been calculated that during cultivation and harvesting, about 5 tons of solid waste are generated per hectare per year [27]. Therefore, the need to recycle, reuse, and recover valuable chemicals from winery waste becomes apparent. There has already been some research done on using wine waste as a source of carbon for BC production [17,28,29]. The cost of the carbon source represents the key factor determining total bacterial cellulose (BC) production costs [30]. Using waste as a source of nutrients for BC production, such as grape bagasse from the wine production industry, would significantly lower the BC production costs, presuming that the BC yield is comparable. Carreira et al. [17] substituted the carbon source in HS medium with several agroforestry industrial residues, including a white grape skin aqueous extract, and found that the production of BC with grape residues was much lower (0.10 g/L) than with the use of HS medium (2.70 g/L). The findings could be related to the absence of nitrogen and phosphorus in the raw material. However, the results of Vazquez et al. [28] also using white grape bagasse from regional wine production, showed different results. They found that the BC production using grape bagasse doubled compared to the yield on HS culture medium, but only when the medium was supplemented with corn steep liquor and the incubation time was extended up to 14 days. Increasing the incubation time to 25 days produced four times more BC when a combination of white grape bagasse and corn steep liquor in modified HS medium was used [29]. In the previously published research on BC production using winery waste, only white bagasse was used, always as a carbon substitute in HS medium, with the addition of other substances (i.e., corn steep). Research on using red grape bagasse for BC production has not yet been reported. Therefore, the purpose of our study is to evaluate red and white grape bagasse as a replacement carbon source in standard HS medium without any other added nutrient source and as a complete replacement for HS medium. The characteristics of bacterial cellulose films produced on different media are evaluated and compared for their water-holding capacity and structural, viscoelastic, and chemical properties.
2. Materials and Methods
2.1. Microorganism and Inoculum Preparation
The bacterial strain of Komagataeibacter xylinus DSM 6513 used in this work was purchased from Leibniz Institute DSMZ (German Collection of Microorganisms and Cell Cultures GmbH). The bacterial culture was grown on solid GOM medium (100 g/L glucose, yeast extract 10 g/L, 20 g/L CaCO3, 15 g/L agar, pH 6.8) and stored in 15% glycerol at –80 °C. The inoculum used in the experiments was prepared from a week-old culture (GOM, 28 °C). Prior to experiments, the bacterial colonies were suspended in sterile distilled water to a selected density (70% turbidity, BioLog, Hayward, CA, USA).
2.2. Cultivation Media and Conditions
Experiments were performed in variations of HS medium (Table 1) at 28 °C for 14 days. The experimental setup was performed under aseptic conditions.
2.3. Preparation of Grape Waste Bagasse Extracts
The white and red grape bagasse used in this work was obtained from an experimental cellar at the Agricultural Institute of Slovenia. White bagasse was from Souvignier Gris grapes and red bagasse from Cabernet Eidos grapes, both tolerant varieties against downy mildew (Plasmopara viticola (Berk. & M.A. Curtis) Berl. & de Toni) and powdery mildew (Uncinula necator (Schwein.) Burrill), sprayed with fungicides only three times during the growing season. In the case of white grapes, immediately after de-stemming and crushing, the mash was pressed with a 55 L water bladder press to a maximum pressure of 1.5 bar (Lancman VSX 55, Gomark d.o.o., Vransko, Slovenia). For red grapes, after de-stemming and crushing, 12-days of maceration with inoculated alcoholic fermentation followed, and, after that, pressing was carried out under the conditions described above.
Grape bagasse obtained after pressing of both varieties was homogenized separately in a blender according to the procedure described by Vazquez et al. [28]. In short, bagasse was boiled at 70 °C with distilled water to achieve proper density, then blended and sieved to obtain almost fluid mash. The measured pH of the white bagasse extract was 3.80 and the red extract 3.50.
2.4. BC Production and Harvest
Bacterial cellulose was produced in different culture media, described in Table 1. In all procedures, the media’s pH was adjusted to 6.0 and sterilized by autoclaving (121 °C, 15 min). For BC production, static incubations were performed in the cell culture plates with a 10 mL capacity per well (Nunclon Delta Surface, Thermo Fisher Scientific, Roskilde, Denmark) for 14 days at 28 °C. We performed each treatment in 6 replicates.
After the cultivation period, the produced BC was rinsed in distilled water and immersed in 0.1 M NaOH at 80 °C for 120 min under mild shaking conditions to remove any attached cells and impurities. The BC was rinsed in distilled water to remove NaOH and dried on a hydrophobic plate at 50 °C for 24 h.
2.5. Determination of Total Sugars in Grape Bagasse Extracts
The sugar concentration was determined using the Lucifer–Schoorl method [31].
2.6. Evaluation of BC Production
BC production was recorded as the dry weight of BC within the volume of medium, according to Equation (1) [28]:
where m is the dry weight of BC (g), and V is the volume (L) of the medium for BC production.
2.7. Water-Holding Capacity (WHC)
For the determination of WHC, wet BC samples were weighed immediately after treatment in NaOH. The samples were quickly dabbed on a paper and then weighed and dried at 50 °C for 24 h to completely remove water; then, we weighed the samples again. WHC was calculated by the following equation [19]:
where mw is the mass of water removed during drying (g), and mdryBC is the dry weight of bacterial cellulose (g).
2.8. Scanning Electron Microscopy (SEM)
For scanning electronic microscope observation, the air-dried BC membrane was mounted on a copper stub using double adhesive tape and coated with gold. SEM photographs were obtained by scanning electron microscope (JSM-6060LV, JEOL, Tokyo, Japan) at 10,000 times magnification, beam voltage 30 kV, working distance 27 mm, and beam spot size 30.
2.9. Dynamic Mechanical Analysis (DMA)
The viscoelastic properties of bacterial cellulose were investigated using a dynamic mechanical apparatus, DMA Q800 (TA Instruments, New Castle, DE, USA). The tension mode of deformation was performed with the amplitude of 10 μm and at the frequency of 10 Hz of oscillation. The samples were cut in rectangular shape with the following dimensions: approx. 8 mm (W) x approx. 30 mm (L) with a clamping length of approx. 8 mm. Dynamic frequency sweep tests were carried out from 0 to 250 °C in temperature, with a 3 °C/min heating rate. The storage (E’) and loss (E”) modulus, damping (tan δ), and glass transition temperature (Tg) as a function of temperature were determined.
2.10. Fourier-Transform Infrared Spectroscopy (FTIR)
BC samples were examined in a Spectrum 3 (Perkin Elmer, Akron, OH, USA) infrared spectrometer using the attenuated transmittance resonance (ATR) technique; 8 scans per sample were taken in the spectral range, from 4000 to 400 cm−1.
3. Results and Discussion
Bacterial cellulose (BC) was produced according to seven different procedures, i.e., using standard HS medium, modified HS medium with the exclusion of carbon or nitrogen sources, modified HS medium where carbon nutrients were partially replaced by white or red grape bagasse extracts, and the use of grape bagasse extract alone as a nutrient source. Exclusion of the nitrogen or carbon source from HS medium (samples HS-C and HS-N; Table 1) did not result in the formation of BC to be analyzed. When the carbon source was excluded from HS media, a very thin film (Figure 1a) that was very delicate and brittle formed after drying; it disintegrated immediately upon contact. Omitting the nitrogen source had a similar effect; the production of BC was insignificant (Figure 1b). In the latter case, only a hazy formation was visible, which could not be analyzed. Other procedures were successful in producing BC films. Photographs taken after incubation are shown in Figure 2.
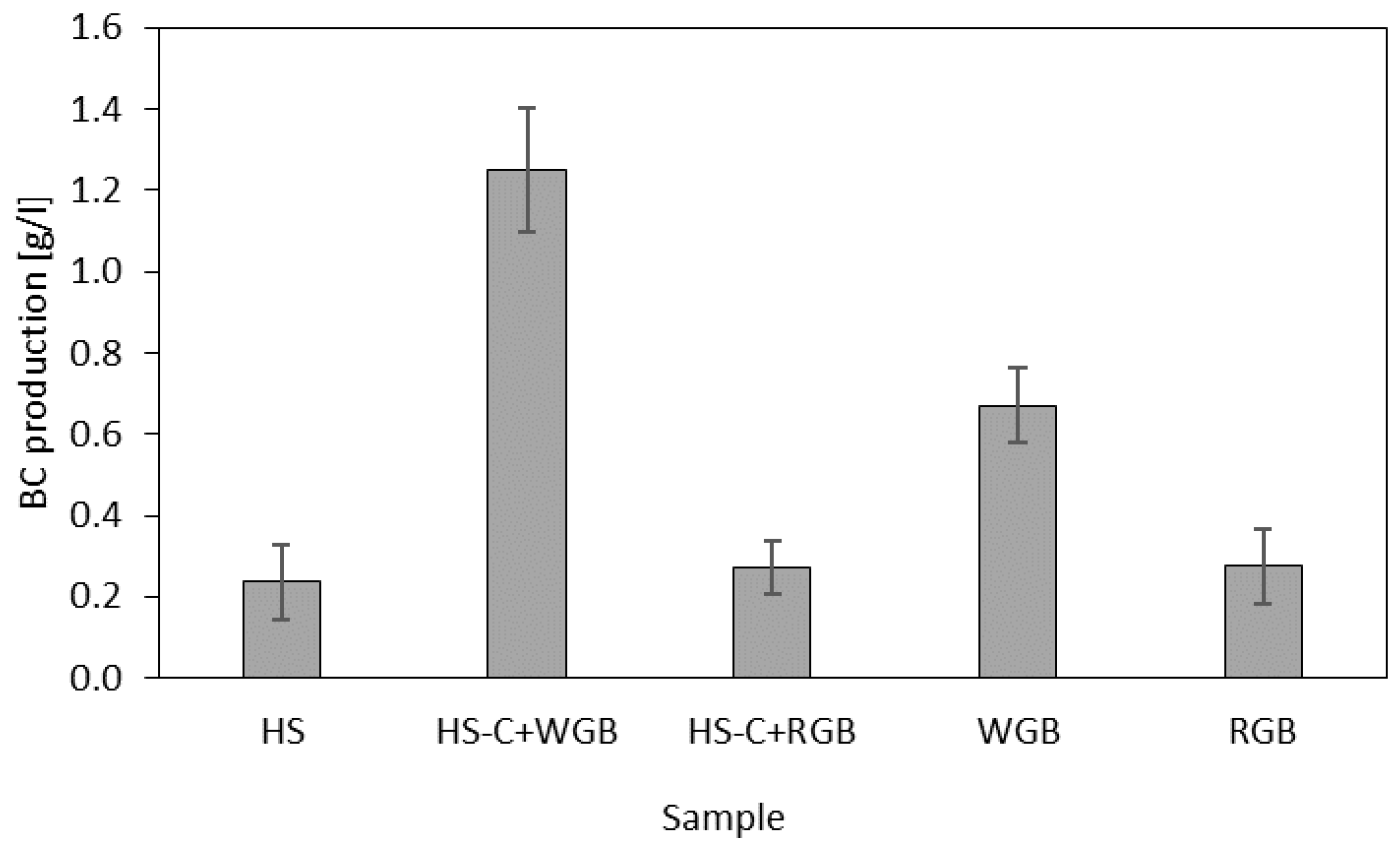
Figure 3 shows the results of bacterial cellulose production. For standard HS medium (sample HS), HS medium with red grape bagasse replacing the carbon source (sample HS -C + RGB), and red grape bagasse as the only nutrient source (sample RGB), the results were quantitatively very similar, with values of 0.24 ± 0.09, 0.27 ± 0.07, and 0.28 ± 0.09 g/L, respectively. In the case of HS-C + WGB, the production of bacterial cellulose was five times higher (1.25 ± 0.09 g/L) than in standard HS medium and almost three times higher when white bagasse was used as the only nutrient (0.67 ± 0.09 g/L; sample WGB). The results are in accordance with the previously published research of Vazquez et al. [28], where it was also found that BC production was higher in cultured HS medium containing white grape bagasse than in standard HS medium; however, their BC production was not as high as in our case. Our results when using white bagasse as the sole culture medium are in agreement with Cerruti et al. [29], who conducted an extensive study on BC production using wine industry residues. They found that using only white bagasse extract containing 60 g/L of sugar, at an incubation period of 20 days at 37 °C, produced 0.548 g/L of BC. Since there has been no research reported on BC production using red bagasse as the nutrient source, no comparisons to other papers could be made. The production of BC stops after the carbon source is depleted or in the case of other nutrient deficiencies [17]. The reason for such high productions of bacterial cellulose when white bagasse is used either as a replacement of carbon source or as the only nutrient source is the sugar content in the extract. The sugar in white and red bagasse extracts was determined by the Lucifer–Schoorl method [31], and the results showed that white bagasse contained 74.2 g/kg and red bagasse 1.5 g/kg of total sugar, while standard HS medium contains 20 g/kg of sugar. The content of total sugars in red grape bagasse is low because the sugars are consumed by yeasts during alcoholic fermentation in the maceration process [32]. The stimulation of BC production using red grape bagasse in culture media could be due to the presence of the flavonoids found in red grapes, which serve as a good carbon source for bacteria [33].
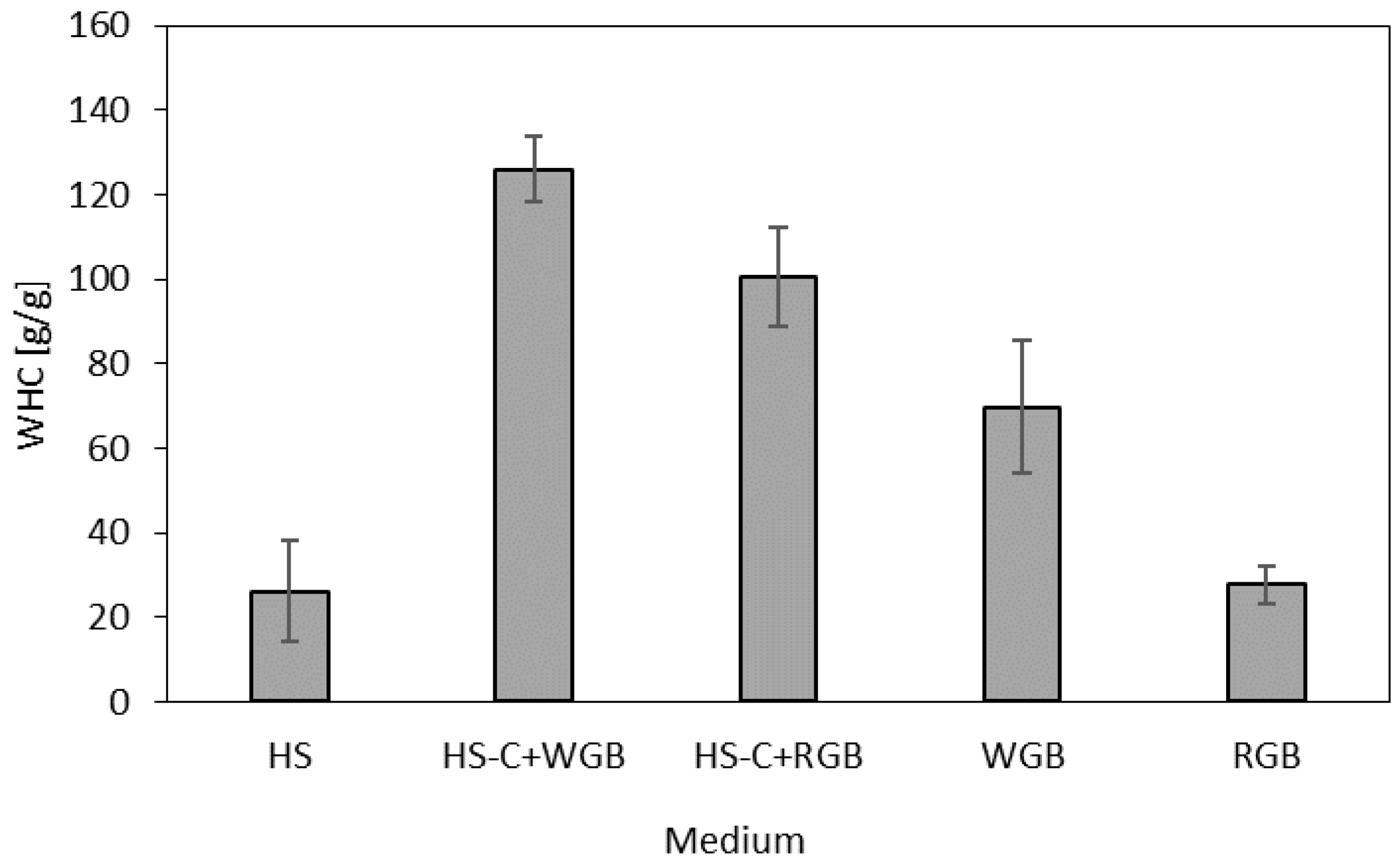
Water-holding capacity (WHC) is an important feature with an important role in different applications of BC films, i.e., for wound healing or as a cell culture substrate or carrier for active ingredients [4,34]. The results in Figure 4 show that WHC varies according to the media used to produce BC. The WHC of the produced BC films reached 126.04 ± 7.82, 100.62 ± 11.79, 69.70 ± 15.68, and 27.79 ± 4.48 g/g for HS-C + WGB, HS-C + RGB, WGB, and RGB, respectively, in comparison to 26.20 ± 12.02 of BC film produced using standard HS media. This indicates that the WHC of all differently prepared BC films was improved, and sample HS-C + WGB had almost five times higher capacity to hold water than sample HS. The WHC of BC depends on its porous structure, fibril arrangement, and surface area [35]. The most important factor for bacterial cellulose to bind water is the numerous hydroxyl groups of fibrils [36]. The hydroxyl groups also interact with each other, establishing intra- and intermolecular hydrogen bonds to form amorphous and crystalline domains, respectively.
The morphological structures of produced BC films were analyzed using scanning electron microscope (Figure 5). Regardless of the medium used to produce BC, the structures of BC films are formed by the intertwining of cellulose fibrils into a 3D structure. The BC film produced using standard HS medium shows a disoriented multilayered porous structure of intertwined fibrils (Figure 5a). The use of white grape bagasse as either a replacement carbon source (Figure 5b) or the only nutrient (Figure 5c) for BC production produces more compact, compressed fibrillar structures of BC films. The individual boundaries between the cellulose nanofibers are not distinct, especially in the case of sample WGB (Figure 5c). Here, we can notice that the surface morphology of the BC film fibrillar structure is the least complex among all samples. The BC films produced using white grape bagasse do have no visible pores on their surface. A completely different surface morphology of BC films appears when red grape bagasse is used in media for the production of BC (Figure 5d,e). The BC films have a very porous surface, with very distinct fibrils. In the case of sample HS-C + RGB (Figure 5d), the fibrils are longitudinally arranged, with exchanging sections of more compact and less compact (very porous) matrices of BC film. Sample RGB (Figure 5d) has a very similar complexity of mesh and multilayered structures as sample HS (Figure 5a). The fibrils are again randomly arranged, forming many spaces (pores) among them.
The samples of bacterial cellulose produced using standard HS medium (HS), HS medium with the replaced carbon source of white grape bagasse (HS-C + WGB), and medium where the nutrition source was only white grape bagasse (WGB) were analyzed by DMA. The samples produced with red grape bagasse were too brittle, and DMA analysis was not possible to perform on them. During the synthesis process of bacterial cellulose, protofibrils of the glucose chain are secreted through bacteria cell walls and aggregate together, forming nanofibril cellulose ribbons [37]. Ross et al. [37] also discussed that cellulose synthesis may be associated with specific sites of adhesion between the inner and outer membranes. Gardner et al. [38] emphasized that there are several proposed theories explaining adhesion phenomena. The bonding phenomenon is the sum of several mechanical, physical, and chemical forces that overlap and influence each other, and it is not possible to separate these forces; adhesion has been characterized as related to mechanical interlocking. The RGB-produced bacterial cellulose films probably have a low degree of polymerization that limits adhesion through interpenetrating networks or mechanical interlocking and are, consequently, more brittle.
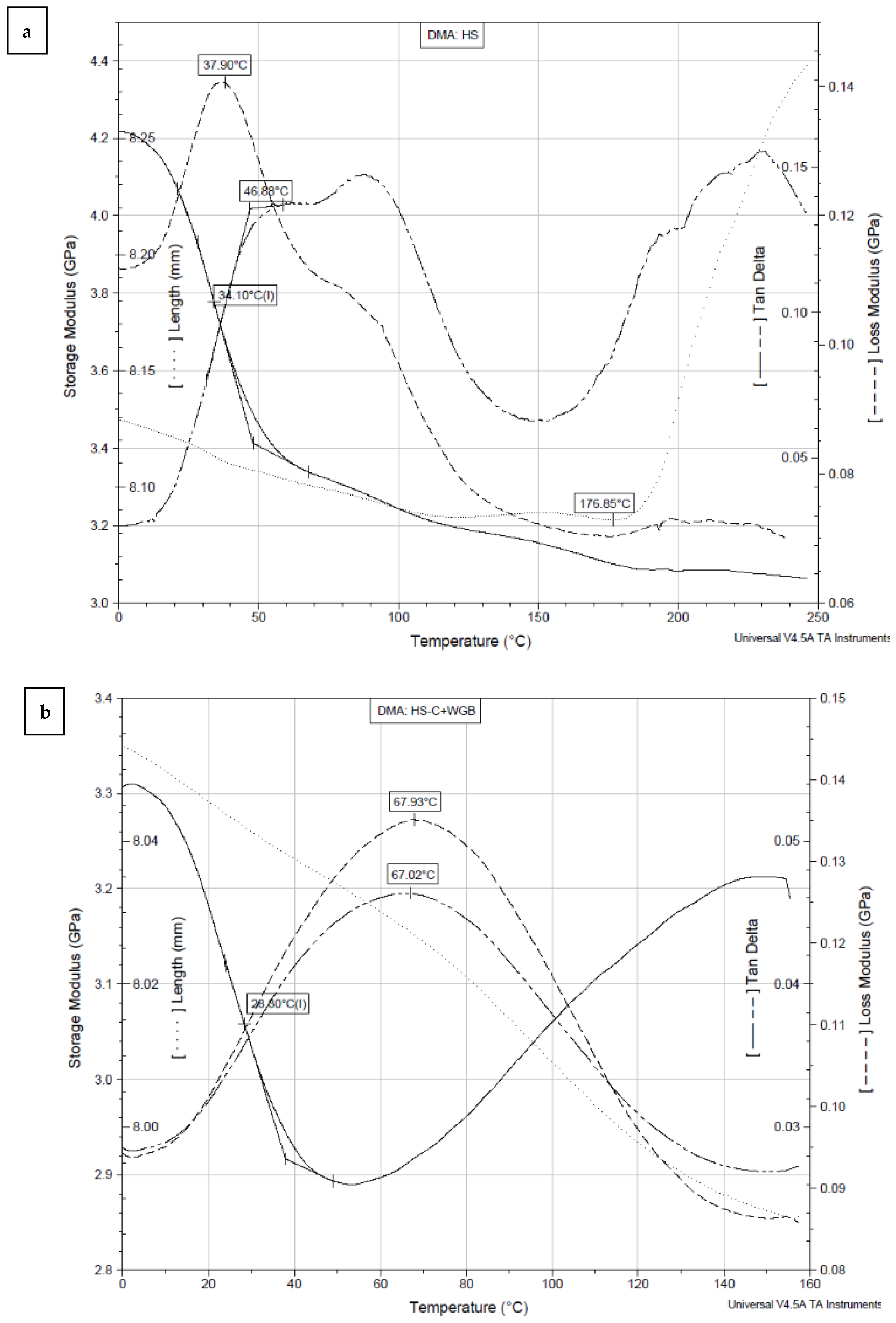
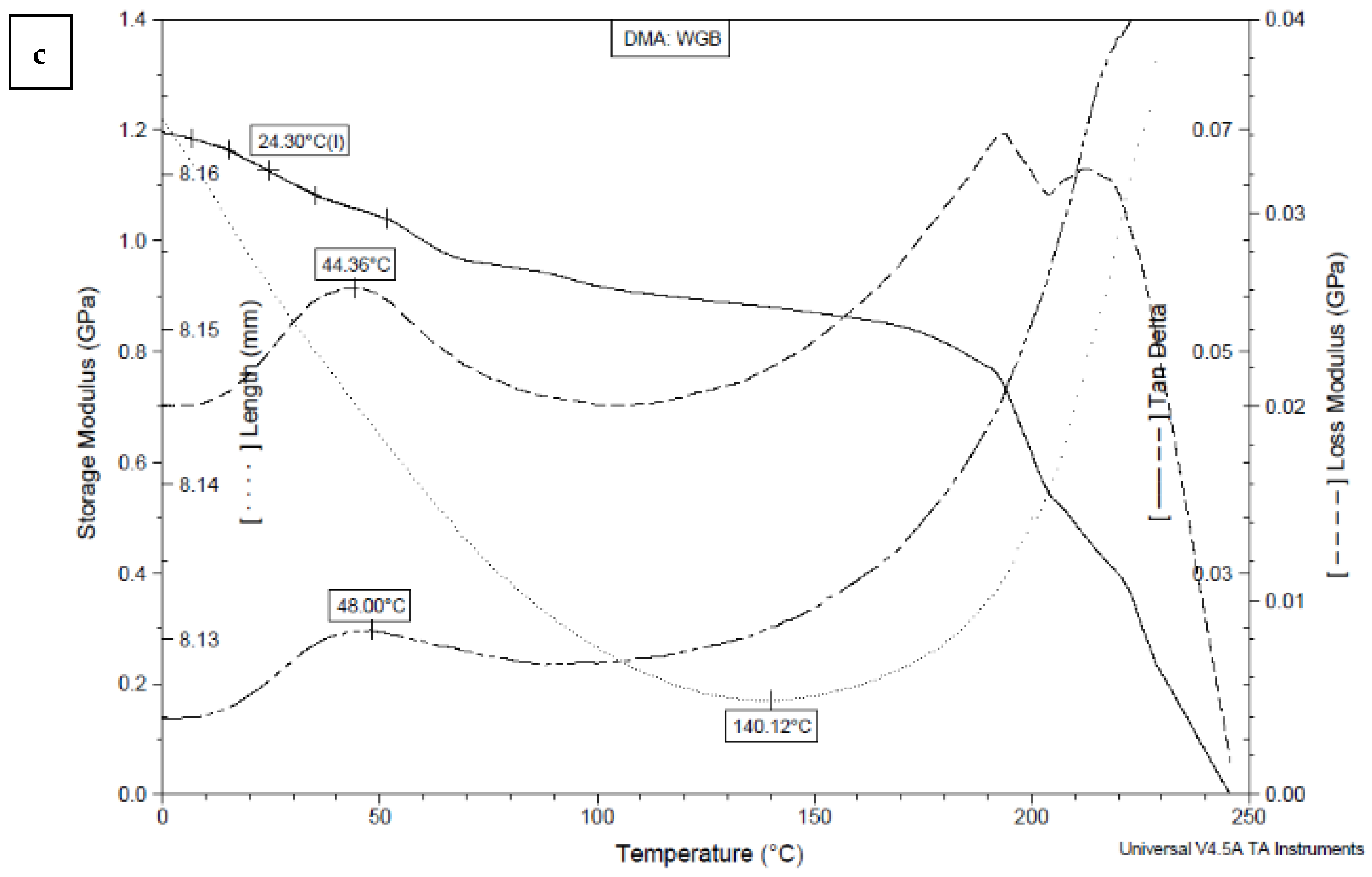
The influence of temperature on the viscoelastic properties (storage modulus, loss modulus, and tan delta) of BC films produced using HS medium and media with white grape bagasse is shown in Figure 6.
The relaxation transition of sample HS (Figure 6a) occurs in the temperature range between 10 and 140 °C. In the range from 0 to 70 °C, the elastic modulus E’ decreases rapidly (by 20.6%). In the whole heating range, the modulus values decrease from 4.22 to 3.06 GPa (27.3%). As the elasticity decreases, the mechanical loss modulus E’’ starts to increase due to the increased mobility of the structural elements; it represents the dissipation of thermal energy per deformation unit. The maximum heat dissipation of the sample is reached at 37.9 °C (Table 2). The tangent delta curve (tan δ), a loss factor representing the internal damping of the structure, peaks at 46.88 °C. The sample length spontaneously shrinks by 0.53% at 176.85 °C, followed by a stronger thermal expansion of the sample. The relaxation transition of the samples HS-C + WGB (Figure 6b) and WGB (Figure 6c) occurs between 10 and 140 °C and between 10 and 90 °C, respectively. The elasticity of sample HS-C + WGB (Figure 6b) decreases rapidly up to a temperature of 50 °C, similar to sample HS (Figure 6a), but then increases again up to a temperature of 160 °C (by 12.6%), unlike sample HS (Figure 6a). The mechanical loss modulus of sample HS-C + WGB (Figure 6b) peaks at 67.93 °C, and the loss factor peaks at 67.02 °C. The length of the sample decreases continuously throughout the temperature range, and the sample shrinks spontaneously by 0.82%. The elasticity of sample WGB (Figure 6c) decreases slowly up to 190 °C (by 9.6% up to 40 °C and by 35.1% up to 190 °C), then a faster decrease occurs until the elasticity reaches zero at 240 °C. The mechanical loss modulus of the WGB sample peaks at 44.36 °C, while the loss factor peaks at 48 °C. The sample length spontaneously shrinks by 0.47% up to 140.12 °C, followed by a sharp thermal expansion of the sample.
The values of the storage modulus (E’), loss modulus (E’’), and damping (tg δ) at 20 °C (room temperature) and glass transition temperatures (Tg) are presented in Table 2.
At the ambient temperature (20 °C), the storage modulus (E’) of sample HS is 4.09 GPa, which slightly decreases to 3.18 GPa when the carbon source is replaced with white grape bagasse (sample HS-C + WGB). The E’ is greatly reduced (to 1.14 GPa) when white grape bagasse is used as the only nutrient for the production of bacterial cellulose (sample WGB). The glass transition temperature (Tg) (determinated by the inflection point of the storage modulus) was highest for sample HS (Tg = 34.10 °C) and lower for samples where white grape bagasse was used either as a replacement for carbon or as the only nutrient for the production of BC. The Tg values of samples HS-C + WGB and WGB are 28.30 and 24.40 °C, respectively. The DMA results show that bacterial cellulose produced using white grape bagasse is less elastic (more flexible), while the bacterial cellulose produced using standard HS medium is more elastic and more rigid (stiffer). To explain the reasons for the changed elasticity, we must first take a closer look at the structure of the BC since the (dynamic) mechanical properties of any material strongly depend on them. On the molecular level, the chemical structure of BC is made of glucose monomers, i.e., a linear homopolymer of glucose monomers linked by β-(1→4) glycosidic linkage with the chemical formula (C6H10O5)n, similar to plant cellulose. However, contrary to it, the two successive monomers in BC are linked in such a way that the former glucose unit is rotated at 180° with reference to the preceding monomer. The degree of BC polymerization only ranges between 2000 and 6000 compared to plant cellulose, which lies in the range 13,000–14,000. The repeated glucose monomers form a continuous long unbranched polymer chain. Several cellulose chains in BC are held together through strong intra- and intermolecular hydrogen bonds that form a cellulose sheet. The hydrogen bonding between the sheets results in a highly crystalline structure, where all of the atoms are fixed in discrete positions with respect to one another, and the molecules of individual microfibrils are tightly packed, which gives the high mechanical strength and high thermal properties of BC [39]. The high crystallinity of BC is also the consequence of the lack of hemicellulose, lignin, or pectin, naturally found in plant cellulose [40]. On the micro-level, BC is an interconnected porous 3D network of random nanofibers that are made of cellulose microfibrils and microfibril bundles. The individual nanofibers range from 25 to 100 nm in diameter to several micrometers in length [41]. The nanofibers are loosely arranged and contain empty spaces (pores), making BC as a highly porous matrix [39]. Contrary to plant-based cellulose, BC consists of pure cellulose without non-cellulosic impurities (hemicellulose, lignin, pectin, and wax) [41]. Molecular chemistry, porosity, fiber arrangement, and orientation are, thus, the main parameters that define the mechanical and thermal properties of BC. As explained above, the change in the elastic behavior (storage–elastic modulus) of our samples is the overall result of the changes in crystallinity and fiber network entanglement. The stiffness of sample HS is thus the consequence of the higher crystallinity of its molecular structure, where molecular chains are arranged in a more perfect crystal lattice, and, at the same time, it is the consequence of close packing and the increasing connectivity between fibrils in the BC network. The greater connectivity is the result of many interactions, including hydrogen bonding, electrostatic, hydrophobic, van der Waals, and even entanglement. The close packing and strengthening restrict polymer chain mobility and increase the toughness of BC. The strong and stiff BC fibrils offer better resistance to the applied force and avoid changes in their structure [42]. The BC films prepared using white grape bagasse show a reduced elastic response to applied tension loads in the whole heated area, suggesting lower crystallinity due to the disturbed formation of 1,4-b-glucan chains and decreased connectivity, resulting in looser, i.e., a less-arranged 3D nanofibrous network structure. This is also evident from the SEM results (Figure 5), as the fibrillar interweaving of WGB samples is the least complex among all the samples. However, the films seem to have a more compact fibrillar structure than other samples and do not have visible pores on their surface. The consequence for lower crystallinity of WGB samples might also be in the content of impurities, as grape bagasse extract also contains non-cellulosic impurities such as minerals and hemicellulose. A similar finding was presented by Goelzer et al. [43], who used rice bark as a carbon cultivation source, where it was found that the crystallinity of BC films dropped from 56% (when using glucose as the carbon source) to 28% (when using rice bark as the carbon source). The polysaccharides disabled cellulose–cellulose binding during the formation of BC, resulting in a weaker BC network [44]. A lack of intense interpolymer chain interaction, especially hydrogen bonding that creates stronger interactions between the chains and microfibrils and enhances the thermal stability of BC, occurs. The less interconnected structure is, thus, more prone to the thermal mobility of the molecular segments (slippage), seeking their most comfortable state by bypassing the relaxation transition.
Since the storage modulus represents the mechanical energy stored by the material during a loading cycle, the loss modulus is proportional to the energy dissipated by the material. The ratio of the loss modulus to the storage modulus is related to the viscosity of the material and is known as the damping factor (tan δ). It defines the way in which a material absorbs and disperses energy. At room temperature (20 °C), before the glass transition temperature of all three samples, where the chain segments are still not fully mobile but also not frozen anymore, the lowest damping is indicated for the WGB film, which has more potential to store the applied external load rather than dissipating it. In contrast, the HS film shows the highest damping, meaning it would rather dissipate the load in the form of scattered heat than store it. This is the consequence of the very pronounced loss modulus of HS film when compared to HS-C + WGB and WGB films at room temperature (Table 2). The higher damping capacity of HS film, despite its high storage modulus, can probably be attributed to its more porous structure (Figure 5a), which prevails over its higher crystallinity where the overall response of this complex 3D network structure is concerned. The greater porosity may also be in correlation with the higher glass transition measurement of the HS films since the porosity is inversely related to thermal conductivity, which increases linearly with bulk density. As stated by Ross et al. [45], materials with higher porosity will take a longer time to reach the desired temperature, resulting in higher detected glass transition temperatures. Additionally, during DMA testing, frictional forces arise in the materials due to the constant supplied oscillatory strain. As suggested by Ross et al. [45], more friction arises in the sample with lower porosity due to the closer proximity of the solid structure on a macroscopic level. While the increased friction occurs in the less porous structure, it may serve as an additional energy source to the thermal energy being supplied to the material during DMA testing. Thus, there is more energy associated with less-porous samples than more-porous samples at the same temperature. This may also be an explanation for the less-porous WGB films presenting lower glass transition regions.
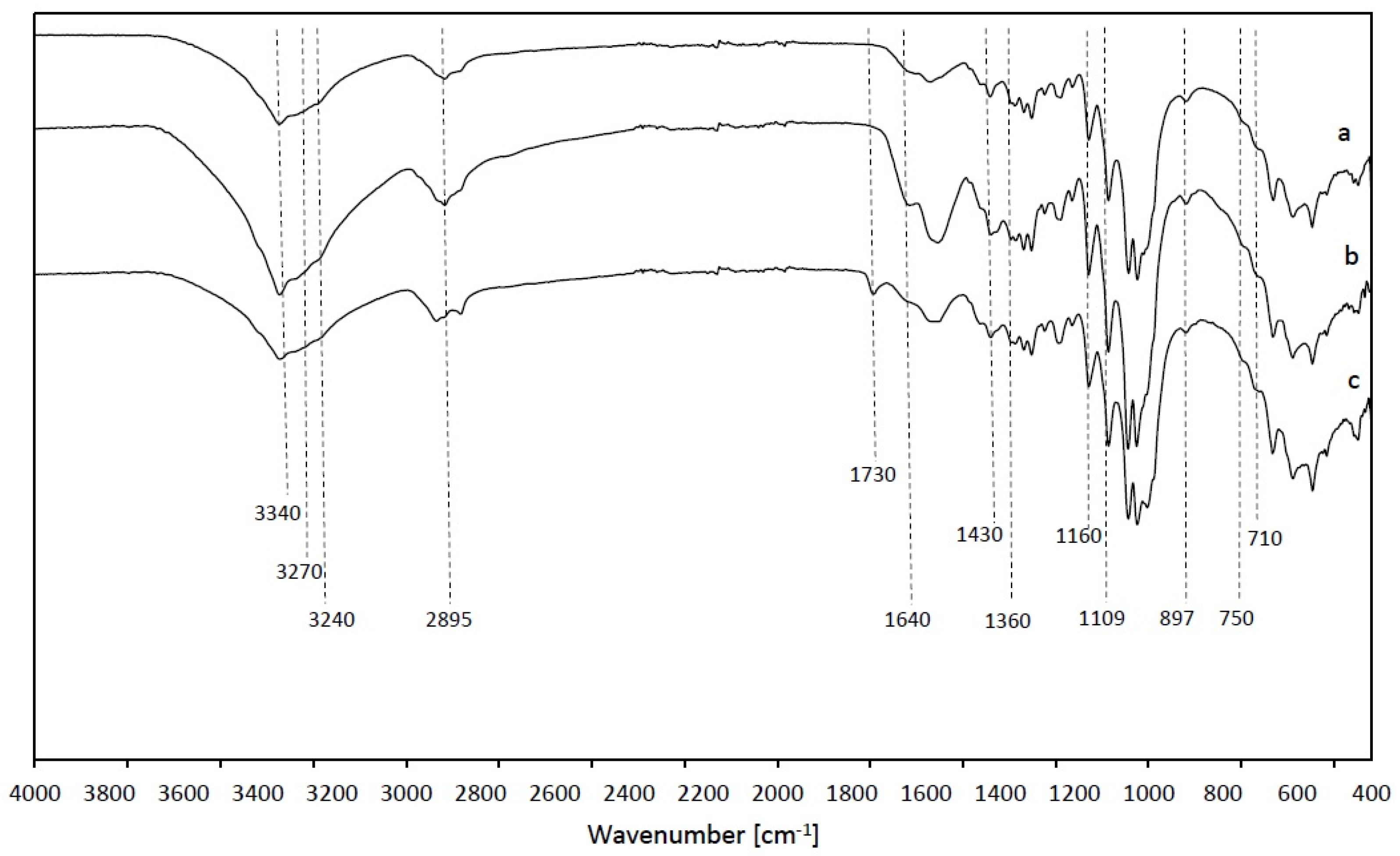
The FTIR spectra of the produced BC films are shown in Figure 7. The observed bands are typical fingerprints of cellulose I, with a strong band at 3340 cm–1 as the stretching vibration on intra and inter O–H groups in cellulose I, around 2895 cm–1 C–H stretching, 1640 cm–1 H–OH (absorbed water), 1430 cm–1 CH2 symmetrical bending, 1109 cm–1 C–O bond stretching, 1360 cm–1 C–H bending, 1160 cm–1 C–OC groups of glycosidic bonds, and 897 cm–1 β-linked glucose polymers [20,29,46]. In the spectrum of sample WGB, an additional peak at 1730 cm–1 is noticed, which is related to the stretching vibrations of conjugated C=O of carbonyl groups, which are attributed to the hemicellulose [47] present in grape bagasse, as already indicated by the results of DMA analysis. All samples of the produced bacterial cellulose contained both allomorphs. The absorbances at 3240 and 750 cm–1 are assigned to the triclinic Iα allomorph, and the absorbances at 3270 and 710 cm–1 are assigned to the monoclinic Iβ form [20,28].
4. Conclusions
The results of the study conducted on the use of white or red grape bagasse as a replacement carbon source in HS medium or as a sole nutrient for the production of bacterial cellulose showed that white grape bagasse is a better substitute for expensive commercial glucose than red grape bagasse. The use of white bagasse as a substitute carbon source resulted in a five times higher yield of bacterial cellulose, with almost five times higher water-holding capacity and greater flexibility than the BC produced in standard HS medium. There were no significant chemical differences between the BC films produced, except for the sample prepared with white grape bagasse extract only, which also contained carbonyl groups due to the presence of hemicellulose. The latter sample had a lower storage modulus and the lowest transition temperature. Although red grape bagasse showed potential as a substitute carbon source or as a sole nutrient source and produced BC films, the films were rather brittle and could not be analyzed by DMA. However, the BC produced in red bagasse media could be of value in an industry where fragmented, highly porous BC particles are needed, such as the food industry. The white bagasse BC, on the other hand, promises greater applicability in a wide range of industries, especially in the textile and biomedical industries as wound dressing, hygienic face masks, and carriers for active ingredients.
Author Contributions
Conceptualization, methodology, M.G., L.O. and J.L.; investigation, L.O.; writing, L.O., M.L., M.G. and J.L.; supervision, M.G.; analysis, L.O., J.L. and M.L.; review M.G., F.Č., J.L. and M.L. All authors have read and agreed to the published version of the manuscript.
Funding
The research was performed as a part of the PhD program of L.O. and was financially supported by the Slovenian Research Agency (programs P2-0213 Textiles and Ecology and P4-0072 Agrobiodiversity) and the Infrastructural Centre (RIC UL-NTF).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bertolino, V.; Cavallaro, G.; Milioto, S.; Lazzara, G. Polysaccharides/Halloysite nanotubes for smart bionanocomposite materials. Carbohydr. Polym. 2020, 245, 116502. [Google Scholar] [CrossRef]
- Lisuzzo, L.; Caruso, M.R.; Cavallaro, G.; Milioto, S.; Lazzara, G. Hydroxypropyl Cellulose Films Filled with Halloysite Nanotubes/Wax Hybrid Microspheres. Ind. Eng. Chem. Res. 2021, 60, 1656–1665. [Google Scholar] [CrossRef]
- Arca, H.C.; Mosquera-Giraldo, L.I.; Bi, V.; Xu, D.; Taylor, L.S.; Edgar, K.J. Pharmaceutical Applications of Cellulose Ethers and Cellulose Ether Esters. Biomacromolecules 2018, 19, 2351–2376. [Google Scholar] [CrossRef]
- Lin, S.-P.; Calvar, I.L.; Catchmark, J.M.; Liu, J.-R.; Demirci, A.; Cheng, K.-C. Biosynthesis, production and applications of bacterial cellulose. Cellulose 2013, 20, 2191–2219. [Google Scholar] [CrossRef]
- Rodrigues, A.C.; Fontão, A.I.; Coelho, A.; Leal, M.; da Silva, F.A.S.; Wan, Y.; Dourado, F.; Gama, M. Response surface statistical optimization of bacterial nanocellulose fermentation in static culture using a low-cost medium. New Biotechnol. 2019, 49, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Singh, O.; Panesar, P.S.; Chopra, H.K. Response surface optimization for cellulose production from agro industrial waste by using new bacterial isolate Gluconacetobacter xylinus C18. Food Sci. Biotechnol. 2017, 26, 1019–1028. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, L.; Hua, J.; Jia, S.; Zhang, J.; Liu, H. Production of nano bacterial cellulose from waste water of candied jujube-processing industry using Acetobacter xylinum. Carbohydr. Polym. 2015, 120, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Ha, J.H.; Shehzad, O.; Khan, S.; Lee, S.Y.; Park, J.W.; Khan, T.; Park, J.K. Production of bacterial cellulose by a static cultivation using the waste from beer culture broth. Korean J. Chem. Eng. 2008, 25, 812–815. [Google Scholar] [CrossRef]
- Azeredo, H.M.C.; Barud, H.; Farinas, C.S.; Vasconcellos, V.M.; Claro, A.M. Bacterial Cellulose as a Raw Material for Food and Food Packaging Applications. Front. Sustain. Food Syst. 2019, 3. [Google Scholar] [CrossRef] [Green Version]
- Hestrin, S.; Schramm, M. Synthesis of cellulose by Acetobacter xylinum. 2. Preparation of freeze-dried cells capable of polymerizing glucose to cellulose. Biochem. J. 1954, 58, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Hussain, Z.; Sajjad, W.; Khan, T.; Wahid, F. Production of bacterial cellulose from industrial wastes: A review. Cellulose 2019, 26, 2895–2911. [Google Scholar] [CrossRef]
- Phruksaphithak, N.; Kaewnun, C.; O-Thong, S. Bacterial cellulose production and applications. Sci. Eng. Health Stud. 2019, 13, 1–7. [Google Scholar] [CrossRef]
- Hungund, B.; Prabhu, S.; Shetty, C.; Acharya, S.; Prabhu, V.; Sg, G. Production of Bacterial Cellulose from Gluconacetobacter persimmonis GH-2 using Dual and Cheaper Carbon Sources. J. Microb. Biochem. Technol. 2013, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Andritsou, V.; De Melo, E.M.; Tsouko, E.; Ladakis, D.; Maragkoudaki, S.; Koutinas, A.A.; Matharu, A.S. Synthesis and Characterization of Bacterial Cellulose from Citrus-Based Sustainable Resources. ACS Omega 2018, 3, 10365–10373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurosumi, A.; Sasaki, C.; Yamashita, Y.; Nakamura, Y. Utilization of various fruit juices as carbon source for production of bacterial cellulose by Acetobacter xylinum NBRC 13693. Carbohydr. Polym. 2009, 76, 333–335. [Google Scholar] [CrossRef]
- Algar, I.; Fernandes, S.C.M.; Mondragon, G.; Castro, C.; García-Astrain, C.; Gabilondo, N.; Retegi, A.; Eceiza, A. Pineapple agroindustrial residues for the production of high value bacterial cellulose with different morphologies. J. Appl. Polym. Sci. 2014, 132, 132. [Google Scholar] [CrossRef]
- Carreira, P.; Mendes, J.A.; Trovatti, E.; Serafim, L.S.; Freire, C.S.; Silvestre, A.; Neto, C. Utilization of residues from agro-forest industries in the production of high value bacterial cellulose. Bioresour. Technol. 2011, 102, 7354–7360. [Google Scholar] [CrossRef] [PubMed]
- Molina-Ramírez, C.; Cañas-Gutiérrez, A.; Castro, C.; Zuluaga, R.; Gañán, P. Effect of production process scale-up on the characteristics and properties of bacterial nanocellulose obtained from overripe Banana culture medium. Carbohydr. Polym. 2020, 240, 116341. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Lopez-Sanchez, P.; Li, R.; Li, Z. Production of bacterial cellulose by Gluconacetobacter hansenii CGMCC 3917 using only waste beer yeast as nutrient source. Bioresour. Technol. 2014, 151, 113–119. [Google Scholar] [CrossRef]
- Castro, C.; Zuluaga, R.; Putaux, J.-L.; Caro, G.; Mondragon, I.; Gañán, P. Structural characterization of bacterial cellulose produced by Gluconacetobacter swingsii sp. from Colombian agroindustrial wastes. Carbohydr. Polym. 2011, 84, 96–102. [Google Scholar] [CrossRef]
- Rani, M.U.; Appaiah, K.A.A. Production of bacterial cellulose by Gluconacetobacter hansenii UAC09 using coffee cherry husk. J. Food Sci. Technol. 2011, 50, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Abdelraof, M.; Hasanin, M.S.; El-Saied, H. Ecofriendly green conversion of potato peel wastes to high productivity bacterial cellulose. Carbohydr. Polym. 2019, 211, 75–83. [Google Scholar] [CrossRef]
- Jozala, A.F.; Pértile, R.A.N.; Dos Santos, C.A.; Santos-Ebinuma, V.D.C.; Seckler, M.M.; Gama, F.M.; Pessoa, A. Bacterial cellulose production by Gluconacetobacter xylinus by employing alternative culture media. Appl. Microbiol. Biotechnol. 2015, 99, 1181–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, F.P.; Silva, N.H.; Trovatti, E.; Serafim, L.S.; Duarte, M.F.; Silvestre, A.; Neto, C.; Freire, C.S. Production of bacterial cellulose by Gluconacetobacter sacchari using dry olive mill residue. Biomass Bioenergy 2013, 55, 205–211. [Google Scholar] [CrossRef]
- FAO. Available online: http://www.fao.org/faostat/en/#data/FBS (accessed on 12 December 2020).
- Galanakis, C. Chapter 3 Vermicomposting of Winemaking By-Products. In Handbook Of Grape Processing By-Products; Elsevier Academic Press: London, UK, 2017; pp. 55–78. [Google Scholar]
- Zacharof, M.-P. Grape Winery Waste as Feedstock for Bioconversions: Applying the Biorefinery Concept. Waste Biomass Valoriz. 2016, 8, 1011–1025. [Google Scholar] [CrossRef] [Green Version]
- Vazquez, A.; Foresti, M.L.; Cerrutti, P.; Galvagno, M.A. Bacterial Cellulose from Simple and Low Cost Production Media by Gluconacetobacter xylinus. J. Polym. Environ. 2013, 21, 545–554. [Google Scholar] [CrossRef]
- Cerrutti, P.; Roldán, P.; García, R.M.; Galvagno, M.A.; Vázquez, A.; Foresti, M.L. Production of bacterial nanocellulose from wine industry residues: Importance of fermentation time on pellicle characteristics. J. Appl. Polym. Sci. 2016, 133, 9. [Google Scholar] [CrossRef]
- Singh, R.; Mathur, A.; Goswami, N.; Mathur, G. Effect of carbon sources on physicochemical properties of bacterial cellulose produced from Gluconacetobacter xylinus MTCC 7795. E-Polymers 2016, 16, 331–336. [Google Scholar] [CrossRef]
- Asquieri, E.R.; de Moura e Silva, A.G.; de Cássia Sousa Mendes, D.; Batista, R.D. Comparison of titulometric and spectrophotometric approaches towards the determination of total soluble and insoluble carbohydrates in foodstuff. Carpathian J. Food Sci. Technol. 2019, 69–79. [Google Scholar] [CrossRef]
- Antonić, B.; Jančíková, S.; Dordević, D.; Tremlová, B. Grape Pomace Valorization: A Systematic Review and Meta-Analysis. Foods 2020, 9, 1627. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-M.; Li, T.-C.; Jhan, Y.-L.; Weng, J.-H.; Chou, C.-H. The Impact of Microbial Biotransformation of Catechin in Enhancing the Allelopathic Effects of Rhododendron formosanum. PLoS ONE 2013, 8, e85162. [Google Scholar] [CrossRef] [Green Version]
- Gorgieva, S. Bacterial Cellulose as a Versatile Platform for Research and Development of Biomedical Materials. Processes 2020, 8, 624. [Google Scholar] [CrossRef]
- Ul-Islam, M.; Khan, T.; Park, J.K. Water holding and release properties of bacterial cellulose obtained by in situ and ex situ modification. Carbohydr. Polym. 2012, 88, 596–603. [Google Scholar] [CrossRef]
- Rebelo, A.R.; Archer, A.J.; Chen, X.; Liu, C.; Yang, G.; Liu, Y. Dehydration of bacterial cellulose and the water content effects on its viscoelastic and electrochemical properties. Sci. Technol. Adv. Mater. 2018, 19, 203–211. [Google Scholar] [CrossRef]
- Ross, P.; Mayer, R.; Benziman, M. Cellulose biosynthesis and function in bacteria. Microbiol. Rev. 1991, 55, 35–58. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.J.; Oporto, G.S.; Mills, R.; Samir, M.A.S.A. Adhesion and Surface Issues in Cellulose and Nanocellulose. J. Adhes. Sci. Technol. 2008, 22, 545–567. [Google Scholar] [CrossRef] [Green Version]
- Nanocellulose; Wiley: Hoboken, NJ, USA, 2019; pp. 81–113.
- Dayal, M.S.; Catchmark, J.M. Mechanical and structural property analysis of bacterial cellulose composites. Carbohydr. Polym. 2016, 144, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-Y.; Buldum, G.; Mantalaris, A.; Bismarck, A. More Than Meets the Eye in Bacterial Cellulose: Biosynthesis, Bioprocessing, and Applications in Advanced Fiber Composites. Macromol. Biosci. 2014, 14, 10–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ul-Islam, M.; Khattak, W.A.; Kang, M.; Kim, S.M.; Khan, T.; Park, J.K. Effect of post-synthetic processing conditions on structural variations and applications of bacterial cellulose. Cellulose 2013, 20, 253–263. [Google Scholar] [CrossRef]
- Goelzer, F.; Faria-Tischer, P.; Vitorino, J.; Sierakowski, M.-R.; Tischer, C. Production and characterization of nanospheres of bacterial cellulose from Acetobacter xylinum from processed rice bark. Mater. Sci. Eng. C 2009, 29, 546–551. [Google Scholar] [CrossRef]
- Cheng, K.-C.; Catchmark, J.M.; Demirci, A. Effects of CMC Addition on Bacterial Cellulose Production in a Biofilm Reactor and Its Paper Sheets Analysis. Biomacromolecules 2011, 12, 730–736. [Google Scholar] [CrossRef]
- Ross, K.A.; Campanella, O.; Okos, M.R. The effect of porosity on glass transition measurement. Int. J. Food Prop. 2002, 5, 611–628. [Google Scholar] [CrossRef]
- Wang, S.-S.; Han, Y.-H.; Ye, Y.-X.; Shi, X.-X.; Xiang, P.; Chen, D.-L.; Li, M. Physicochemical characterization of high-quality bacterial cellulose produced by Komagataeibacter sp. strain W1 and identification of the associated genes in bacterial cellulose production. RSC Adv. 2017, 7, 45145–45155. [Google Scholar] [CrossRef] [Green Version]
- Nejad, S.M.M.; Madhoushi, M.; Vakili, M.; Rasouli, D. Evaluation of degradation in chemical compounds of wood in historical buildings using FT-IR and FT-Raman vibrational spectroscopy. Maderas. Cienc. Y Tecnol. 2019, 21, 381–392. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Excluding a (a) carbon or (b) nitrogen source from HS media either produces BC film that is too brittle to analyze or does not produce BC at all.
Figure 1.
Excluding a (a) carbon or (b) nitrogen source from HS media either produces BC film that is too brittle to analyze or does not produce BC at all.

Figure 2.
BC produced from (a) HS medium, (b) HS-C + WGB medium, (c) WGB medium, (d) HS-C + RGB medium, and (e) RGB medium.
Figure 2.
BC produced from (a) HS medium, (b) HS-C + WGB medium, (c) WGB medium, (d) HS-C + RGB medium, and (e) RGB medium.


Figure 3.
Bacterial cellulose production (g/L) in different culture media.

Figure 4.
Water-holding capacity (g/g) of BC produced in different culture media.

Figure 5.
Scanning electron microscope images of BC films produced from (a) HS medium, (b) HS-C + WGB medium, (c) WGB medium, (d) HS-C + RGB medium, and (e) RGB medium.
Figure 5.
Scanning electron microscope images of BC films produced from (a) HS medium, (b) HS-C + WGB medium, (c) WGB medium, (d) HS-C + RGB medium, and (e) RGB medium.


Figure 6.
The viscoelastic curves and parameters of samples: (a) HS, (b) HS-C + WGB, and (c) WGB at 10 Hz of oscillation.
Figure 6.
The viscoelastic curves and parameters of samples: (a) HS, (b) HS-C + WGB, and (c) WGB at 10 Hz of oscillation.


Figure 7.
FTIR spectra of bacterial cellulose samples. (a) HS; (b) HS-C + WGB; (c) WGB.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Bacterial cellulose (BC) preparation treatments and sample coding according to the procedure.
Table 1.
Bacterial cellulose (BC) preparation treatments and sample coding according to the procedure.
| Medium | Medium Composition | Sample Coding of Produced BC |
|---|---|---|
| Standard HS medium | 8 g/L glucose 2 g/L yeast extract 2 g/L peptone 1.8 g/L Na2HPO4 distilled water to 1 L | HS |
| HS medium without a carbon source | 2 g/L yeast extract 2 g/L peptone 1.8 g/L Na2HPO4 distilled water to 1 L | HS-C |
| HS medium without a nitrogen source | 1.5 g/L glucose 0.34 g/L Na2HPO4 distilled water to 1 L | HS-N |
| HS medium with white grape extract as a replacement of carbon source | HS-C: white bagasse extract = 1:1 (v/v) | HS-C + WGB |
| HS-C with red grape extract as a carbon source | HS-C: red bagasse extract = 1:1 (v/v) | HS-C + RGB |
| White grape extract as a sole nutrient source | white grape bagasse extract only | WGB |
| Red grape extract as a sole nutrient source | red grape bagasse extract only | RGB |
Table 2.
Storage modulus (E’), loss modulus (E’’), and tangent delta (tan δ) at 20 °C and glass transition temperature Tg, loss modulus (E’’) peak temperature, and tangent delta (tan δ) peak temperature of samples HS, HS-C + WGB, and WGB.
Table 2.
Storage modulus (E’), loss modulus (E’’), and tangent delta (tan δ) at 20 °C and glass transition temperature Tg, loss modulus (E’’) peak temperature, and tangent delta (tan δ) peak temperature of samples HS, HS-C + WGB, and WGB.
| Sample | E’ at 20 °C [GPa] | E’’ at 20 °C [GPa] | tan δ at 20 °C | Tg [°C] * | E’’ Peak [°C] | tan δ Peak [°C] |
|---|---|---|---|---|---|---|
| HS | 4.09 | 0.12 | 0.04 | 34.10 | 37.90 | 46.88 |
| HS-C + WGB | 3.18 | 0.01 | 0.03 | 28.30 | 67.93 | 67.02 |
| WGB | 1.14 | 0.03 | 0.02 | 24.30 | 44.36 | 48.00 |
* Tg is determined by the inflection point of the storage modulus.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ogrizek, L.; Lamovšek, J.; Čuš, F.; Leskovšek, M.; Gorjanc, M. Properties of Bacterial Cellulose Produced Using White and Red Grape Bagasse as a Nutrient Source. Processes 2021, 9, 1088. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9071088
AMA Style
Ogrizek L, Lamovšek J, Čuš F, Leskovšek M, Gorjanc M. Properties of Bacterial Cellulose Produced Using White and Red Grape Bagasse as a Nutrient Source. Processes. 2021; 9(7):1088. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9071088
Chicago/Turabian StyleOgrizek, Linda, Janja Lamovšek, Franc Čuš, Mirjam Leskovšek, and Marija Gorjanc. 2021. "Properties of Bacterial Cellulose Produced Using White and Red Grape Bagasse as a Nutrient Source" Processes 9, no. 7: 1088. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9071088
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.