Nonlinear Analysis for a Type-1 Diabetes Model with Focus on T-Cells and Pancreatic β-Cells Behavior


Abstract
:1. Introduction
2. Mathematical Model of T1DM
2.1. Localization of Compact Invariant Sets (LCIS)
2.1.1. Mathematical Preliminaries
2.1.2. Mathematical Development
2.2. LCIS for the Invariant Plane
2.3. LCIS for
3. Nonlinear Controller Design
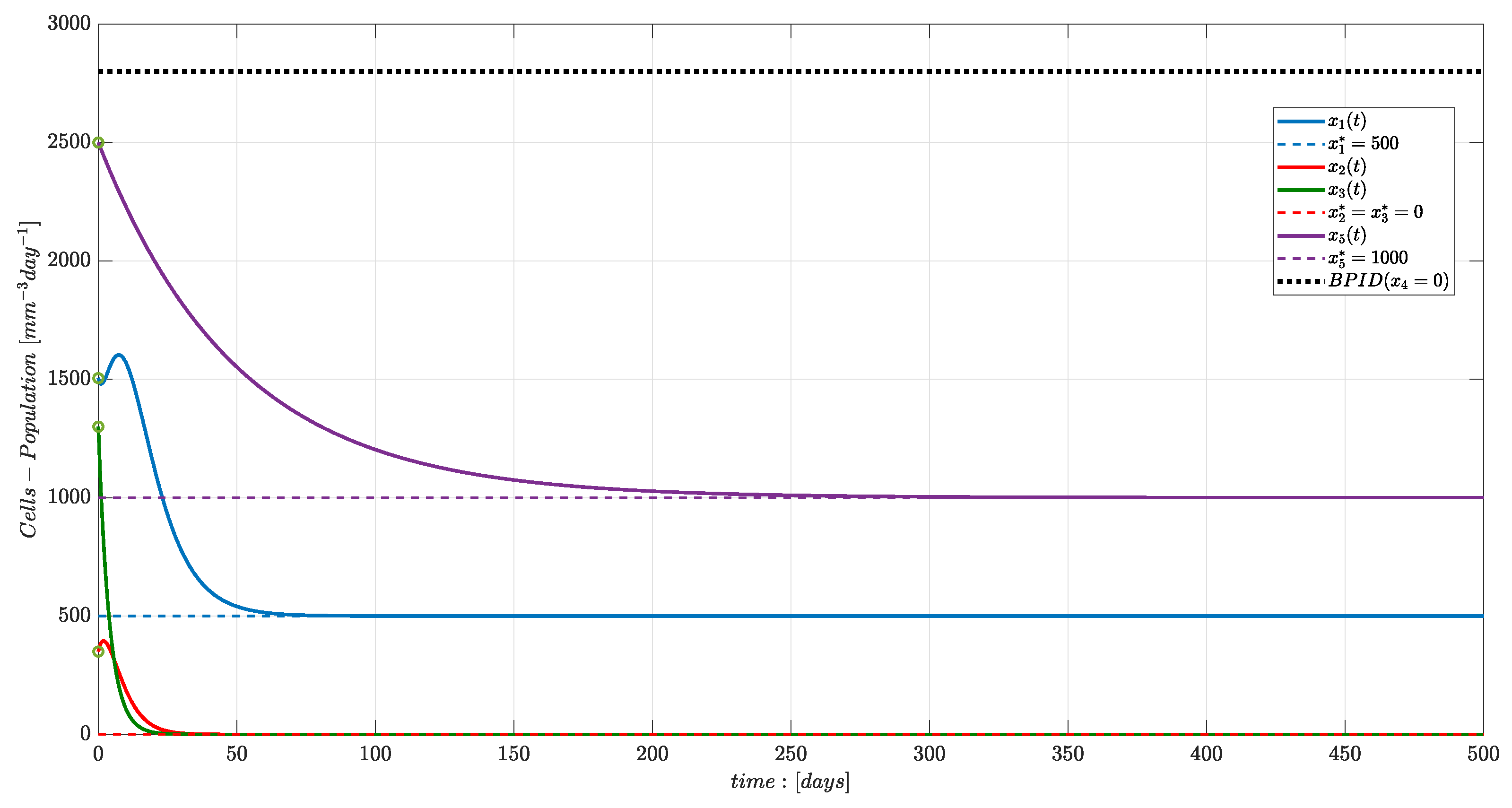
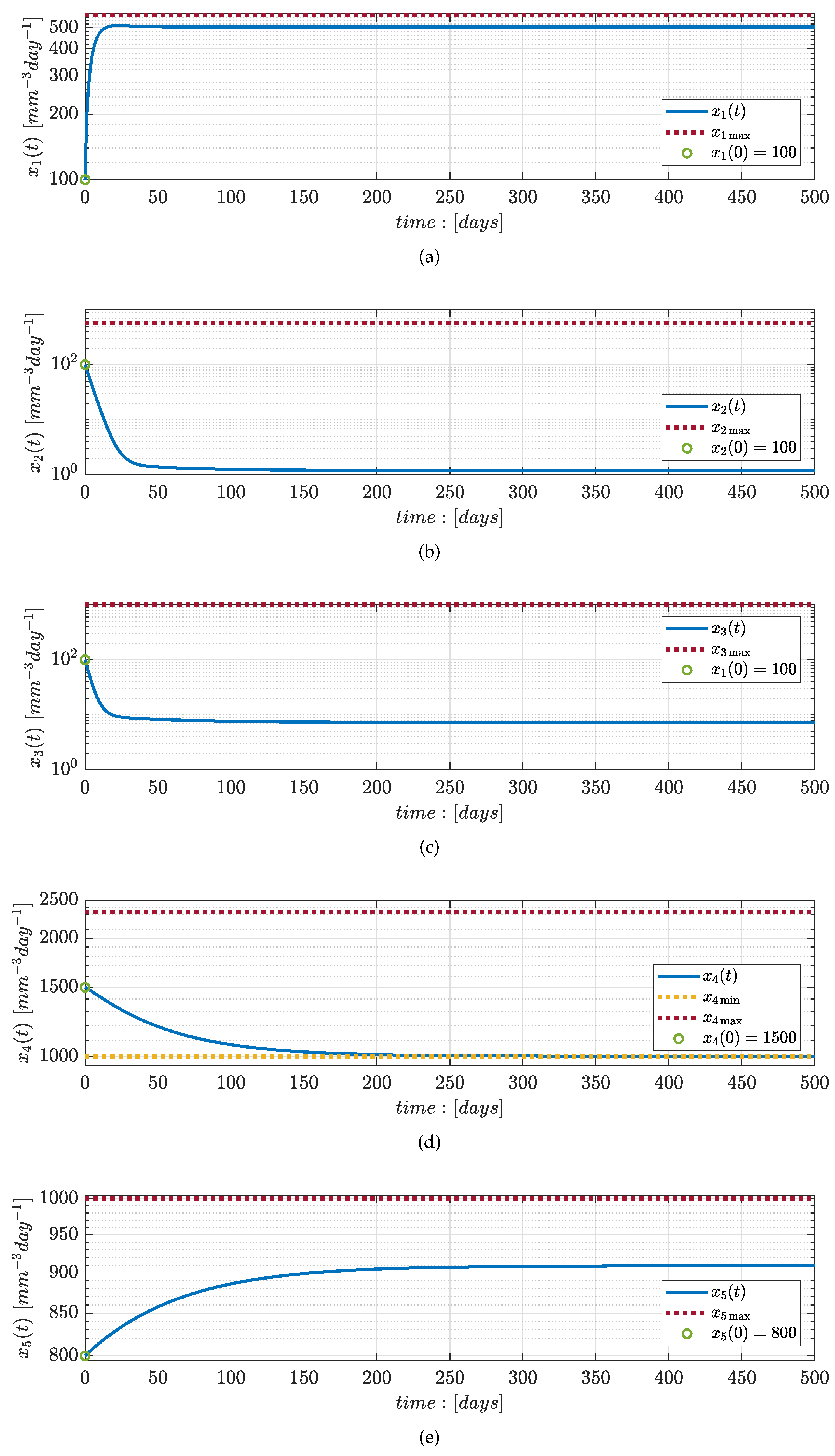
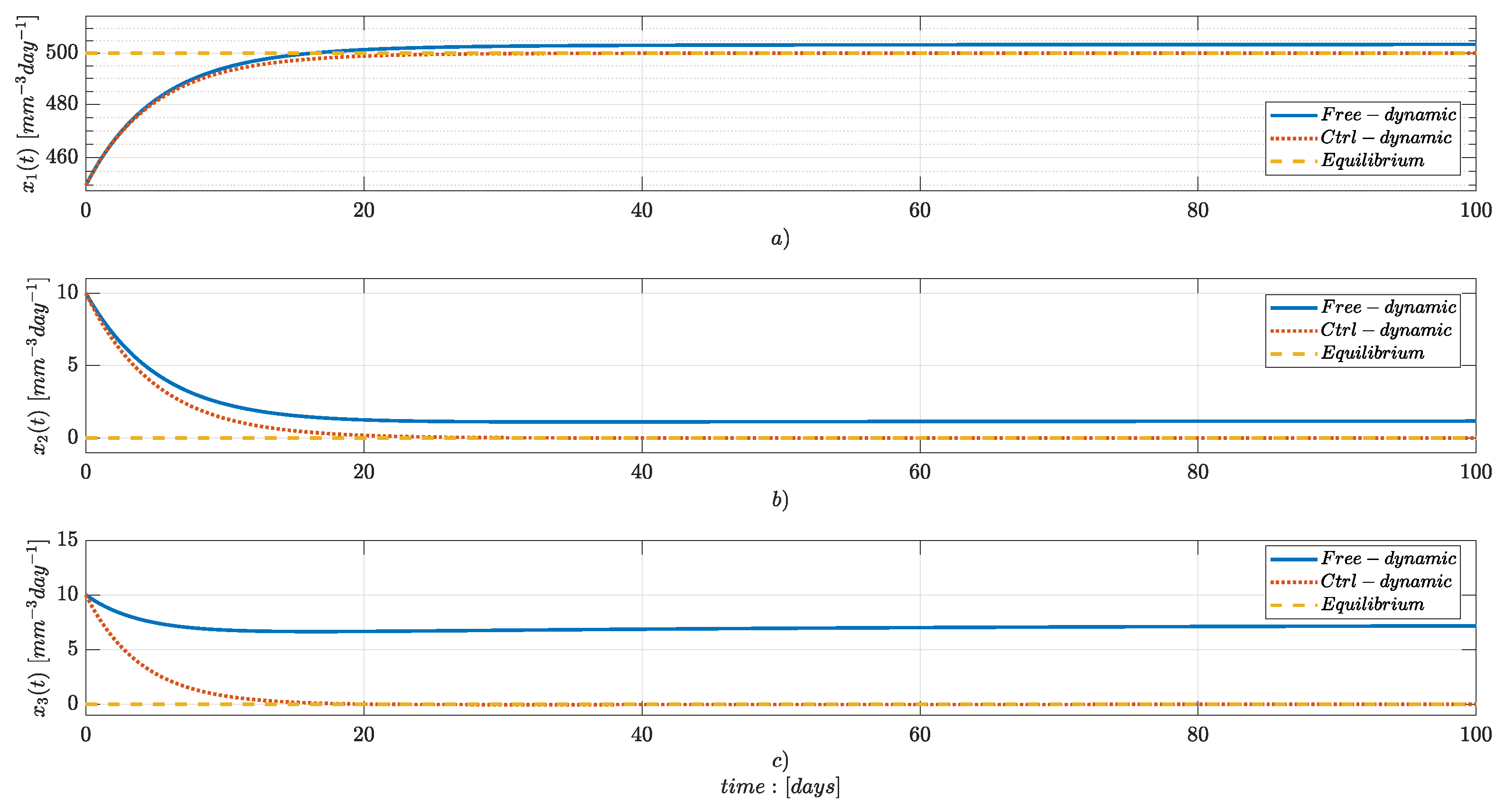
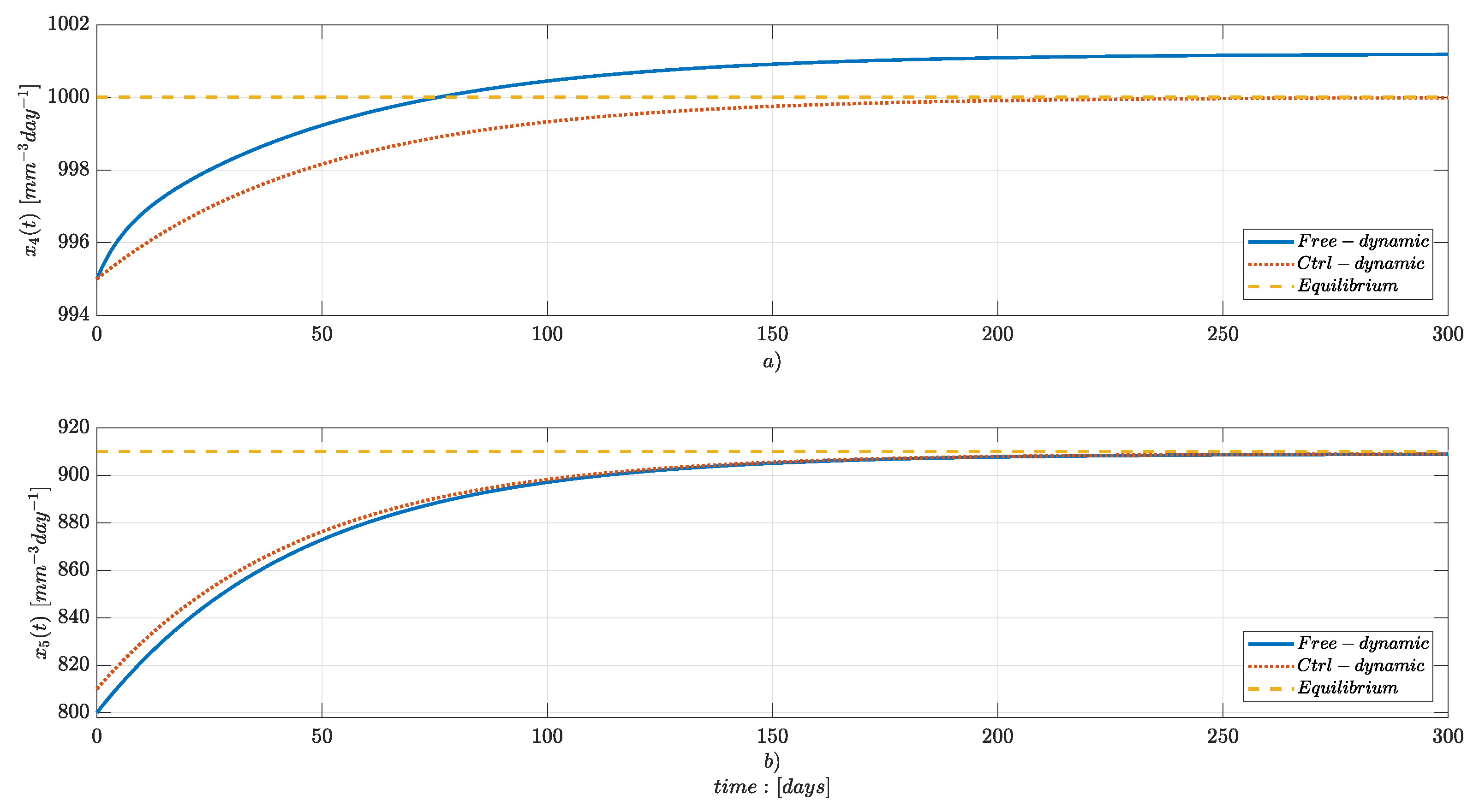
Numerical Simulations
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Forlenza, G.P.; Rewers, M. The epidemic of type 1 diabetes: What is it telling us? Curr. Opin. Endocrinol. Diabetes Obes. 2011, 18, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and Regional Diabetes Prevalence Estimates for 2019 and Projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saarela, K.; Tuomilehto, J.; Sund, R.; Keskimäki, I.; Hartikainen, S.; Pukkala, E. Cancer incidence among Finnish people with type 2 diabetes during 1989–2014. Eur. J. Epidemiol. 2019, 34, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Lu, Y.; Hajifathalian, K.; Bentham, J.; Di Cesare, M.; Danaei, G.; Bixby, H.; Cowan, M.; Ali, M.; Taddei, C.; et al. Worldwide trends in diabetes since 1980: A pooled analysis of 751 population-based studies with 4·4 million participants. Lancet 2016, 387, 1513–1530. [Google Scholar] [CrossRef] [Green Version]
- Harding, J.L.; Pavkov, M.E.; Magliano, D.J.; Shaw, J.E.; Gregg, E.W. Global trends in diabetes complications: A review of current evidence. Diabetologia 2019, 62, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Cho, N.H.; Shaw, J.E.; Karuranga, S.; Huang, Y.; da Rocha Fernandes, J.D.; Ohlrogge, A.W.; Malanda, B. IDF Diabetes Atlas: Global estimates of diabetes prevalence for 2017 and projections for 2045. Diabetes Res. Clin. Pract. 2018, 138, 271–281. [Google Scholar] [CrossRef]
- Akinsola, V.O.; Oluyo, T.O. Mathematical analysis with numerical solutions of the mathematical model for the complications and control of diabetes mellitus. J. Stat. Manag. Syst. 2019, 22, 845–869. [Google Scholar] [CrossRef]
- Bakhti, M.; Böttcher, A.; Lickert, H. Modelling the endocrine pancreas in health and disease. Nat. Rev. Endocrinol. 2019, 15, 155–171. [Google Scholar] [CrossRef] [Green Version]
- Ajmera, I.; Swat, M.; Laibe, C.; Le Novere, N.; Chelliah, V. The impact of mathematical modeling on the understanding of diabetes and related complications. CPT Pharmacomet. Syst. Pharmacol. 2013, 2, 1–14. [Google Scholar] [CrossRef]
- Fernández-Díaz, C.; Escobar-Curbelo, L.; López-Acosta, J.F.; Lobatón, C.D.; Moreno, A.; Sanz-Ortega, J.; Perdomo, G.; Cózar-Castellano, I. Insulin degrading enzyme is up-regulated in pancreatic β cells by insulin treatment. Histol. Histopathol. 2018. [Google Scholar] [CrossRef]
- Senior, P.; Pettus, J. Stem cell therapies for Type 1 diabetes: Current status and proposed road map to guide successful clinical trials. Diabet. Med. 2019, 36, 297–307. [Google Scholar] [CrossRef]
- Magdelaine, N.; Chaillous, L.; Guilhem, I.; Poirier, J.Y.; Krempf, M.; Moog, C.H.; Le Carpentier, E. A long-term model of the glucose–insulin dynamics of type 1 diabetes. IEEE Trans. Biomed. Eng. 2015, 62, 1546–1552. [Google Scholar] [CrossRef] [PubMed]
- Alali, H.; Boutayeb, W.; Boutayeb, A.; Merabet, N. A mathematical model on the effect of growth hormone on glucose homeostasis. ARIMA J. 2019, 30, 31–42. [Google Scholar]
- Valle, P.A.; Coria, L.N.; Gamboa, D.; Plata, C. Bounding the Dynamics of a Chaotic-Cancer Mathematical Model. Math. Probl. Eng. 2018, 2018, 9787015. [Google Scholar] [CrossRef]
- Starkov, K.E. On dynamic tumor eradication conditions under combined chemical/anti-angiogenic therapies. Phys. Lett. A 2018, 382, 387–393. [Google Scholar] [CrossRef]
- Li, P.; Yu, L.; Fang, Q.; Lee, S.Y. A simplification of Cobelli’s glucose–insulin model for type 1 diabetes mellitus and its FPGA implementation. Med. Biol. Eng. Comput. 2016, 54, 1563–1577. [Google Scholar] [CrossRef]
- Shabestari, P.S.; Rajagopal, K.; Safarbali, B.; Jafari, S.; Duraisamy, P. A Novel Approach to Numerical Modeling of Metabolic System: Investigation of Chaotic Behavior in Diabetes Mellitus. Complexity 2018, 2018, 6815190. [Google Scholar] [CrossRef] [Green Version]
- Kitano, H. A robustness-based approach to systems-oriented drug design. Nat. Rev. Drug Discov. 2007, 6, 202. [Google Scholar] [CrossRef]
- Parsa, N.T.; Vali, A.; Ghasemi, R. Back stepping sliding mode control of blood glucose for type I diabetes. World Acad. Sci. Eng. Technol. Int. J. Med. Health Biomed. Bioeng. Pharm. Eng. 2014, 8, 779–783. [Google Scholar]
- Babar, S.A.; Rana, I.A.; Arslan, M.; Zafar, M.W. Integral Backstepping Based Automated Control of Blood Glucose in Diabetes Mellitus Type 1 Patients. IEEE Access. 2019, 7, 173286–173293. [Google Scholar] [CrossRef]
- Vakili, S.; ToosianShandiz, H. Back-stepping sliding mode control design for glucose regulation in type 1 diabetic patients. Int. J. Nonlinear Anal. Appl. 2019, 10, 167–176. [Google Scholar]
- Patra, A.K.; Rout, P.K. Backstepping sliding mode Gaussian insulin injection control for blood glucose regulation in type I diabetes patient. J. Dyn. Syst. Meas. Control 2018, 140, 091006. [Google Scholar] [CrossRef]
- Nath, A.; Dey, R.; Aguilar-Avelar, C. Observer based nonlinear control design for glucose regulation in type 1 diabetic patients: An LMI approach. Biomed. Signal Process. Control 2019, 47, 7–15. [Google Scholar] [CrossRef]
- Lawrence, J.M.; Mayer-Davis, E.J. What do we know about the trends in incidence of childhood-onset type 1 diabetes? Diabetologia 2019, 62, 370–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzo-Almorós, A.; Hang, T.; Peiró, C.; Soriano-Guillén, L.; Egido, J.; Tuñón, J.; Lorenzo, Ó. Predictive and diagnostic biomarkers for gestational diabetes and its associated metabolic and cardiovascular diseases. Cardiovasc. Diabetol. 2019, 18, 140. [Google Scholar] [CrossRef] [PubMed]
- Magombedze, G.; Nduru, P.; Bhunu, C.P.; Mushayabasa, S. Mathematical modelling of immune regulation of type 1 diabetes. Biosystems 2010, 102, 88–98. [Google Scholar] [CrossRef]
- Efrat, S. Beta-cell replacement for insulin-dependent diabetes mellitus. Adv. Drug Deliv. Rev. 2008, 60, 114–123. [Google Scholar] [CrossRef]
- Krishchenko, A.P. Localization of invariant compact sets of dynamical systems. Differ. Equ. 2005, 41, 1669–1676. [Google Scholar] [CrossRef]
- Krishchenko, A.P.; Starkov, K.E. Localization of compact invariant sets of the Lorenz system. Phys. Lett. A 2006, 353, 383–388. [Google Scholar] [CrossRef]
- Jones, A.; Hattersley, A. The clinical utility of C-peptide measurement in the care of patients with diabetes. Diabet. Med. 2013, 30, 803–817. [Google Scholar] [CrossRef] [Green Version]
- Leighton, E.; Sainsbury, C.A.; Jones, G.C. A practical review of C-peptide testing in diabetes. Diabetes Ther. 2017, 8, 475–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Definition | Value | Units |
|---|---|---|---|
| a | Macrophage supply | 50 | mm day |
| b | Macrophage induced supply | day | |
| c | Macrophage death rate | mm day | |
| g | Rate of antigen uptake | day | |
| k | Macrophage deactivation | day | |
| Induced -cell damage | day | ||
| m | Decay rate of -cell proteins | day | |
| q | Damage of autolytic cells on -cells | mm day | |
| Supply of autolytic cells | 20 | mm day | |
| s | Proliferation of autolytic T-cells | day | |
| Death rate of T-cells | day | ||
| Supply of -cells | 20 | mm day | |
| Death rate of -cells | mm day |
| Localizing Functions | Conditions | Localizing Set |
|---|---|---|
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gamboa, D.; Vázquez, C.E.; Campos, P.J. Nonlinear Analysis for a Type-1 Diabetes Model with Focus on T-Cells and Pancreatic β-Cells Behavior. Math. Comput. Appl. 2020, 25, 23. https://0-doi-org.brum.beds.ac.uk/10.3390/mca25020023
Gamboa D, Vázquez CE, Campos PJ. Nonlinear Analysis for a Type-1 Diabetes Model with Focus on T-Cells and Pancreatic β-Cells Behavior. Mathematical and Computational Applications. 2020; 25(2):23. https://0-doi-org.brum.beds.ac.uk/10.3390/mca25020023
Chicago/Turabian StyleGamboa, Diana, Carlos E. Vázquez, and Paul J. Campos. 2020. "Nonlinear Analysis for a Type-1 Diabetes Model with Focus on T-Cells and Pancreatic β-Cells Behavior" Mathematical and Computational Applications 25, no. 2: 23. https://0-doi-org.brum.beds.ac.uk/10.3390/mca25020023