
Enrichment Free qPCR for Rapid Identification and Quantification of Campylobacter jejuni, C. coli, C. lari, and C. upsaliensis in Chicken Meat Samples by a New Couple of Primers




Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
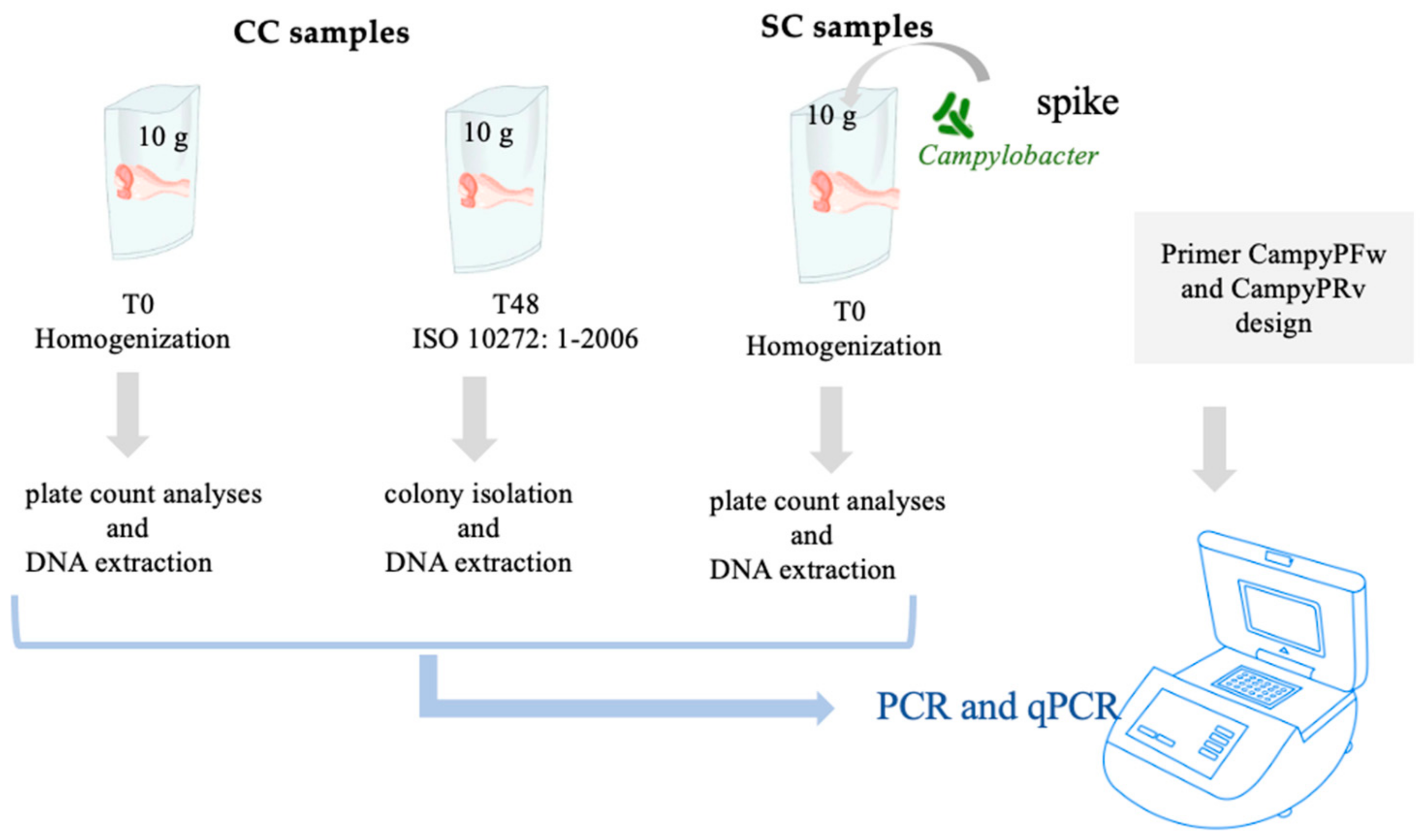
2.2. Chicken Samples and Plate Count Enumeration
2.3. DNA Extraction
2.4. Primer Design
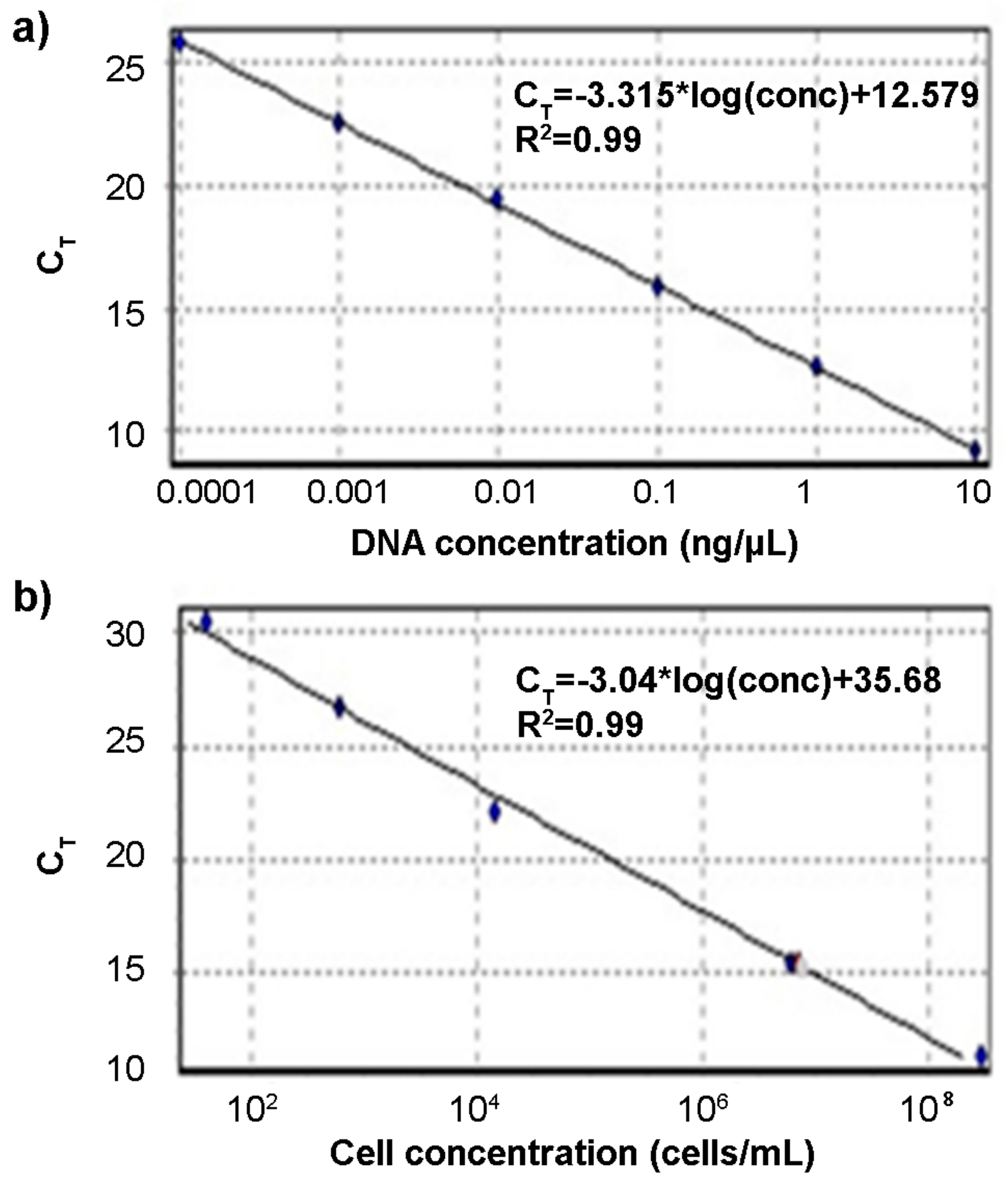
2.5. PCR and qPCR Protocols
3. Results and Discussion
3.1. Microbiological Analysis of Samples
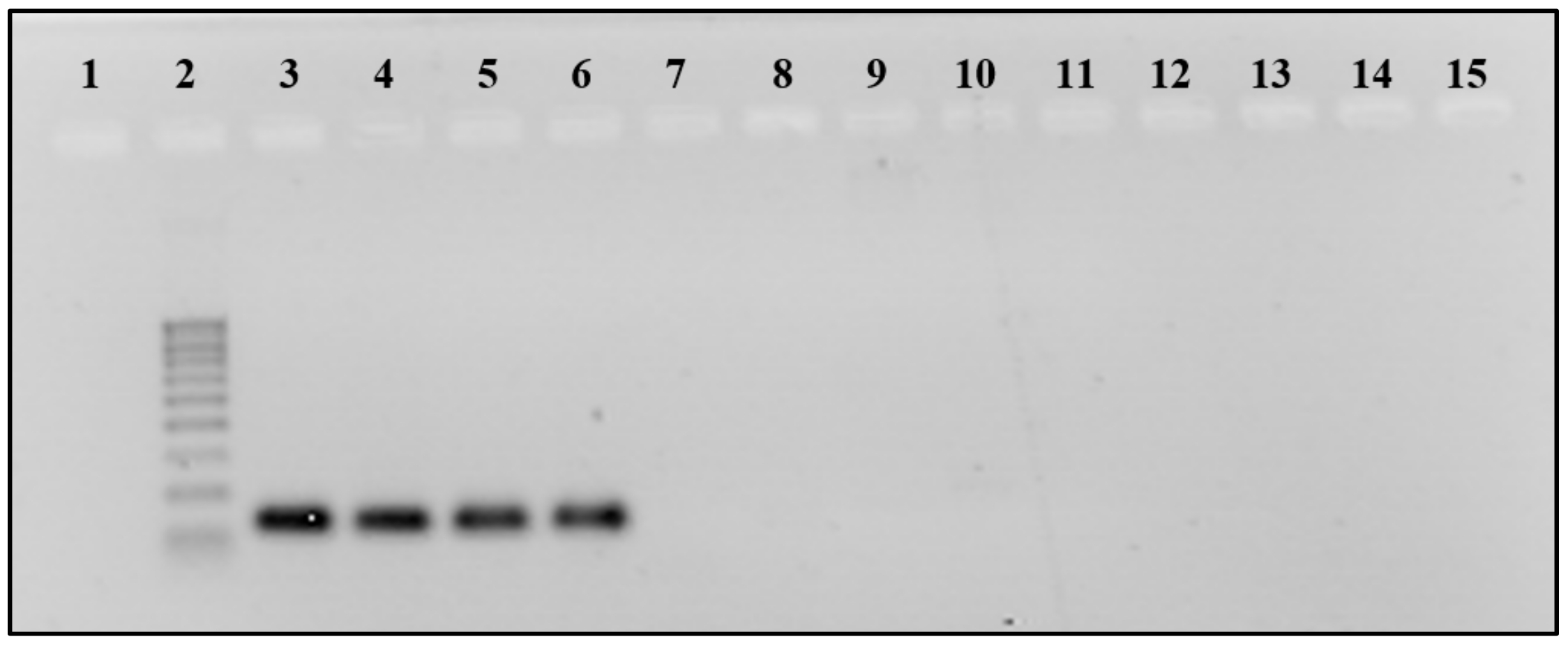
3.2. PCR and qPCR Analysis
3.3. Sequencing
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vizzini, P.; Braidot, M.; Vidic, J.; Manzano, M. Electrochemical and optical biosensors for the detection of campylobacter and listeria: An update look. Micromachines 2019, 10, 500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scallan Walter, E.J.; Crim, S.M.; Bruce, B.B.; Griffin, P.M. Incidence of Campylobacter-associated Guillain-Barre Syndrome estimated from health insurance data. Foodborne Pathog. Dis. 2020, 17, 23–28. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Yao, X.; Gunther, N.W.; Xie, Y.; Tu, S.-I.; Shi, X. Simultaneous detection and differentiation of Campylobacter jejuni, C. coli, and C. lari in chickens using a multiplex real-time PCR assay. Food Anal. Methods 2010, 3, 321–329. [Google Scholar] [CrossRef]
- Nastasijevic, I.; Proscia, F.; Boskovic, M.; Glisic, M.; Blagojevic, B.; Sorgentone, S.; Kirbis, A.; Ferri, M. The European Union control strategy for Campylobacter spp. in the broiler meat chain. J. Food Saf. 2020, 40, e12819. [Google Scholar] [CrossRef]
- El-Shibiny, A.; Scott, A.; Timms, A.; Metawea, Y.; Connerton, P.; Connerton, I. Application of a group II Campylobacter bacteriophage to reduce strains of Campylobacter jejuni and Campylobacter coli colonizing broiler chickens. J. Food Prot. 2009, 72, 733–740. [Google Scholar] [CrossRef]
- Stingl, K.; Knüver, M.-T.; Vogt, P.; Buhler, C.; Krüger, N.-J.; Alt, K.; Tenhagen, B.-A.; Hartung, M.; Schroeter, A.; Ellerbroek, L. Quo vadis?—Monitoring Campylobacter in Germany. Eur. J. Microbiol. Immunol. 2012, 2, 88–96. [Google Scholar] [CrossRef] [Green Version]
- Haddad, N.; Burns, C.M.; Bolla, J.M.; Prévost, H.; Fédérighi, M.; Drider, D.; Cappelier, J.M. Long-term survival of Campylobacter jejuni at low temperatures is dependent on polynucleotide phosphorylase activity. Appl. Environ. Microbiol. 2009, 75, 7310–7318. [Google Scholar] [CrossRef] [Green Version]
- Vidic, J.; Manzano, M.; Chang, C.-M.; Jaffrezic-Renault, N. Advanced biosensors for detection of pathogens related to livestock and poultry. Vet. Res. 2017, 48, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Vizzini, P.; Manzano, M.; Farre, C.; Meylheuc, T.; Chaix, C.; Ramarao, N.; Vidic, J. Highly sensitive detection of Campylobacter spp. in chicken meat using a silica nanoparticle enhanced dot blot DNA biosensor. Biosens. Bioelectron. 2020, 171, 112689. [Google Scholar] [CrossRef]
- Vidic, J.; Vizzini, P.; Manzano, M.; Kavanaugh, D.; Ramarao, N.; Zivkovic, M.; Radonic, V.; Knezevic, N.; Giouroudi, I.; Gadjanski, I. Point-of-need DNA testing for detection of foodborne pathogenic bacteria. Sensors 2019, 19, 1100. [Google Scholar] [CrossRef] [Green Version]
- Ricke, S.C.; Feye, K.M.; Chaney, W.E.; Shi, Z.; Pavlidis, H.; Yang, Y. Developments in rapid detection methods for the detection of foodborne campylobacter in the United States. Front. Microbiol. 2019, 9, 3280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botteldoorn, N.; van Coillie, E.; Piessens, V.; Rasschaert, G.; Debruyne, L.; Heyndrickx, M.; Herman, L.; Messens, W. Quantification of Campylobacter spp. in chicken carcass rinse by real-time PCR. J. Appl. Microbiol. 2008, 105, 1909–1918. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.; Botteldoorn, N.; Coucke, W.; Denayer, S.; Dierick, K.; Uyttendaele, M. Effect of exposure to stress conditions on propidium monoazide (PMA)-qPCR based Campylobacter enumeration in broiler carcass rinses. Food Microbiol. 2015, 48, 182–190. [Google Scholar] [CrossRef]
- Josefsen, M.H.; Löfström, C.; Hansen, T.B.; Christensen, L.S.; Olsen, J.E.; Hoorfar, J. Rapid quantification of viable Campylobacter bacteria on chicken carcasses, using real-time PCR and propidium monoazide treatment, as a tool for quantitative risk assessment. Appl. Environ. Microbiol. 2010, 76, 5097–5104. [Google Scholar] [CrossRef] [Green Version]
- Englen, M.D.; Kelley, L.C. A rapid DNA isolation procedure for the identification of Campylobacter jejuni by the polymerase chain reaction. Lett. Appl. Microbiol. 2000, 31, 421–426. [Google Scholar] [CrossRef]
- Reis, L.P.; Menezes, L.D.M.; Lima, G.K.; Santos, E.L.D.S.; Dorneles, E.M.S.; Assis, D.C.S.D.; Lage, A.P.; Cançado, S.D.V.; Figueiredo, T.C.D. Detection of Campylobacter spp. in chilled and frozen broiler carcasses comparing immunoassay, PCR and real time PCR methods. Ciência Rural 2018, 48, e20161034. [Google Scholar] [CrossRef] [Green Version]
- De Boer, P.; Rahaoui, H.; Leer, R.; Montijn, R.; van der Vossen, J. Real-time PCR detection of Campylobacter spp.: A comparison to classic culturing and enrichment. Food Microbiol. 2015, 51, 96–100. [Google Scholar] [CrossRef]
- Lund, M.; Nordentoft, S.; Pedersen, K.; Madsen, M. Detection of Campylobacter spp. in chicken fecal samples by real-time PCR. J. Clin. Microbiol. 2004, 42, 5125–5132. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, M.; Singh, R.; Dharmasena, M.; Gong, C.; Krastanov, A.; Jiang, X. Rapid identification of Campylobacter jejuni from poultry carcasses and slaughtering environment samples by real-time PCR. Poult. Sci. 2014, 93, 1587–1597. [Google Scholar] [CrossRef] [PubMed]
- Papić, B.; Pate, M.; Henigman, U.; Zajc, U.; Gruntar, I.; Biasizzo, M.; Ocepek, M.; Kušar, D. New approaches on quantification of Campylobacter jejuni in poultry samples: The use of digital PCR and real-time PCR against the ISO standard plate count method. Front. Microbiol. 2017, 8, 331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnider, A.; Overesch, G.; Korczak, B.; Kuhnert, P. Comparison of real-time PCR assays for detection, quantification, and differentiation of Campylobacter jejuni and Campylobacter coli in broiler neck skin samples. J. Food Prot. 2010, 73, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Borges, K.A.; Cisco, I.C.; Furian, T.Q.; Tedesco, D.C.; Rodrigues, L.B.; Do Nascimento, V.P.; dos Santos, L.R. Detection and quantification of Campylobacter spp. in Brazilian poultry processing plants. J. Infect. Dev. Ctries. 2020, 14, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Cocolin, L.; Aggio, D.; Manzano, M.; Cantoni, C.; Comi, G. An application of PCR-DGGE analysis to profile the yeast populations in raw milk. Int. Dairy J. 2002, 12, 407–411. [Google Scholar] [CrossRef]
- Manzano, M.; Cocolin, L.; Cantoni, C.; Comi, G. Bacillus cereus, Bacillus thuringiensis and Bacillus mycoides differentiation using a PCR-RE technique. Int. J. Food Microbiol. 2003, 81, 249–254. [Google Scholar] [CrossRef]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef] [PubMed]
- Vizzini, P.; Beltrame, E.; Zanet, V.; Vidic, J.; Manzano, M. Development and Evaluation of qPCR Detection Method and Zn-MgO/Alginate Active Packaging for Controlling Listeria monocytogenes Contamination in Cold-Smoked Salmon. Foods 2020, 9, 1353. [Google Scholar] [CrossRef]
- Klijn, N.; Weerkamp, A.H.; de Vos, W.M. Identification of mesophilic lactic acid bacteria by using polymerase chain reaction-amplified variable regions of 16S rRNA and specific DNA probes. Appl. Environ. Microbiol. 1991, 57, 3390–3393. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Fontanot, M.; Iacumin, L.; Cecchini, F.; Comi, G.; Manzano, M. Rapid detection and differentiation of important Campylobacter spp. in poultry samples by dot blot and PCR. Food Microbiol. 2014, 43, 28–34. [Google Scholar] [CrossRef]
- Alves, J.; Hirooka, E.Y.; de Oliveira, T.C.R.M. Development of a multiplex real-time PCR assay with an internal amplification control for the detection of Campylobacter spp. and Salmonella spp. in chicken meat. LWT Food Sci. Technol. 2016, 72, 175–181. [Google Scholar] [CrossRef]
- Wolffs, P.; Norling, B.R.; Hoorfar, J.; Griffiths, M.; Rådström, P. Quantification of Campylobacter spp. in chicken rinse samples by using flotation prior to real-time PCR. Appl. Environ. Microbiol. 2005, 71, 5759–5764. [Google Scholar] [CrossRef] [Green Version]
- Wolffs, P.F.; Glencross, K.; Norling, B.; Griffiths, M.W. Simultaneous quantification of pathogenic Campylobacter and Salmonella in chicken rinse fluid by a flotation and real-time multiplex PCR procedure. Int. J. Food Microbiol. 2007, 117, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Edge, T. Development of a novel triplex PCR assay for the detection and differentiation of thermophilic species of Campylobacter using 16S-23S rDNA internal transcribed spacer (ITS) region. J. Appl. Microbiol. 2007, 103, 2561–2569. [Google Scholar] [CrossRef] [PubMed]
- Pacholewicz, E.; Swart, A.; Lipman, L.J.; Wagenaar, J.A.; Havelaar, A.H.; Duim, B. Propidium monoazide does not fully inhibit the detection of dead Campylobacter on broiler chicken carcasses by qPCR. J. Microbiol. Methods 2013, 95, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Rédei, G.P. Genetics Manual: Current Theory, Concepts, Terms; World Scientific: Singapore, 1998. [Google Scholar]
- Osimani, A.; Aquilanti, L.; Pasquini, M.; Clementi, F. Prevalence and risk factors for thermotolerant species of Campylobacter in poultry meat at retail in Europe. Poult. Sci. 2017, 96, 3382–3391. [Google Scholar] [CrossRef] [PubMed]
- Rasschaert, G.; Houf, K.; van Hende, J.; De Zutter, L. Campylobacter contamination during poultry slaughter in Belgium. J. Food Prot. 2006, 69, 27–33. [Google Scholar] [CrossRef]
- Lynch, Ó.A.; Cagney, C.; McDowell, D.A.; Duffy, G. Occurrence of fastidious Campylobacter spp. in fresh meat and poultry using an adapted cultural protocol. Int. J. Food Microbiol. 2011, 150, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Sinulingga, T.S.; Aziz, S.A.; Bitrus, A.A.; Zunita, Z.; Abu, J. Occurrence of Campylobacter species from broiler chickens and chicken meat in Malaysia. Trop. Anim. Health Prod. 2020, 52, 151–157. [Google Scholar] [CrossRef]
- Di Giannatale, E.; Calistri, P.; Di Donato, G.; Decastelli, L.; Goffredo, E.; Adriano, D.; Mancini, M.E.; Galleggiante, A.; Neri, D.; Antoci, S. Thermotolerant Campylobacter spp. in chicken and bovine meat in Italy: Prevalence, level of contamination and molecular characterization of isolates. PLoS ONE 2019, 14, e0225957. [Google Scholar] [CrossRef] [Green Version]
- Szosland-Fałtyn, A.; Bartodziejska, B.; Krolasik, J.; Paziak-Domańska, B.; Korsak, D.; Chmiela, M. The prevalence of Campylobacter spp. in Polish poultry meat. Pol. J. Microbiol. 2018, 67, 117–120. [Google Scholar] [CrossRef] [Green Version]
- Korsak, D.; Maćkiw, E.; Rożynek, E.; Żyłowska, M. Prevalence of Campylobacter spp. in retail chicken, turkey, pork, and beef meat in Poland between 2009 and 2013. J. Food Prot. 2015, 78, 1024–1028. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Samples | Total Viable Count | Enterobacteriaceae | Coliforms | E. coli | Yeasts | Molds |
|---|---|---|---|---|---|---|
| 1 CC | 9.9 × 107 | 3.8 × 103 | 2.0 × 103 | 1.7 × 103 | 2.3 × 104 | <50 * |
| 2 CC | 5.1 × 105 | 1.1 × 103 | 9.8 × 101 | 2.0 × 102 | 1.9 × 103 | <50 * |
| 3 CC | 1.9 × 105 | 9.4 × 102 | 1.3 × 101 | 7.4 × 102 | 7.8 × 102 | <50 * |
| 4 CC | 4.8 × 106 | 1.2 × 104 | 3.0 × 103 | 5.3 × 103 | 9.4 × 104 | <50 * |
| 5 CC | 9.5 × 106 | 2.1 × 104 | 7.0 × 103 | 1.3 × 103 | 6.3 × 105 | <50 * |
| 6 CS | 7.3 × 105 | 1.7 × 103 | 2.3 × 103 | 7.6 × 102 | 6.6 × 104 | <50 * |
| 7 CC | 9.6 × 105 | 9.2 × 103 | 6.4 × 103 | 4.7 × 102 | 3.3 × 104 | <50 * |
| 8 CC | 9.8 × 104 | 5.2 × 103 | 5.2 × 103 | 3.2 × 102 | 1.5 × 103 | <50 * |
| 9 CC | 1.8 × 105 | 2.7 × 103 | 1.4 × 103 | 3.5 × 102 | 2.2 × 103 | <50 * |
| 10 CC | 1.3 × 104 | 1.5 × 103 | 7.7 × 102 | 8.3 × 101 | 1.7 × 102 | <50 * |
| 11 CC | 2.1 × 106 | 2.1 × 102 | 4.2 × 101 | 2.0 × 101 | 6.1 × 103 | <50 * |
| 12 CC | 7.0 × 105 | 2.6 × 103 | <50 * | 8.0 × 102 | 2.0 × 104 | <50 * |
| 13 CC | 1.2 × 107 | 9.6 × 103 | 9.5 × 101 | 2.7 × 102 | 3.5 × 104 | <50 * |
| 14 CC | 1.7 × 107 | 2.4 × 103 | 3.0 × 102 | 0.6 × 103 | 5.3 × 104 | <50 * |
| 15 CC | 3.7 × 107 | 6.9 × 103 | 6.5 × 101 | 2.2 × 103 | 4.5 × 104 | <50 * |
| 16 CC | 1.1 × 107 | 1.6 × 104 | 5.6 × 102 | 2.6 × 102 | 2.4 × 105 | <50 * |
| 17 CC | <50 * | 3.3 × 102 | <50 * | 3.1 × 102 | <50 * | <50 * |
| 18 CC | 3.3 × 109 | 2.1 × 104 | 3.2 × 104 | 6.0 × 102 | 4.9 × 105 | <50 * |
| 19 CC | 3.8 × 109 | 5.4 × 103 | 1.3 × 104 | <50 * | 3.9 × 105 | <50 * |
| 20 CC | 5.1 × 109 | 2.4 × 104 | 2.4 × 104 | 1.5 × 102 | 5.4 × 105 | <50 * |
| Sample | mCCDA * | SKR ° | CAB § | Confirmation Medium CAB | Oxidase | Motility | |
|---|---|---|---|---|---|---|---|
| 41.5 °C, Aerobic | 25 °C, Microaerophilic | ||||||
| 1 CC | + | − | + | + | − | + | − |
| 2 CC | + | − | + | − | − | + | + |
| 3 CC | + | − | + | − | − | + | + |
| 4 CC | + | − | + | + | + | + | − |
| 5 CC | + | − | + | + | + | + | − |
| 6 CC | + | − | + | + | + | + | − |
| 7 CC | + | − | + | + | + | + | − |
| 8 CC | + | − | + | − | − | + | + |
| 9 CC | + | + | + | + | + | + | − |
| 10 CC | + | − | + | − | − | + | + |
| 11 CC | − | − | |||||
| 12 CC | − | − | |||||
| 13 CC | − | − | |||||
| 14 CC | − | − | |||||
| 15 CC | − | − | |||||
| 16 CC | + | + | + | − | − | + | + |
| 17 CC | + | + | + | − | − | + | + |
| 18 CC | + | + | + | + | + | + | − |
| 19 CC | + | + | + | + | + | + | − |
| 20 CC | + | + | + | + | + | + | − |
| CC Samples | t0 | t48 | |||
|---|---|---|---|---|---|
| Mean Ct ± DS | Cell/mL | DNA fg/uL | Mean Ct ± DS | DNA fg/uL | |
| 1 CC | 27.55 ± 0.25 | - | - | 26.99 ± 0.16 | - |
| 2 CC | 24.87 ± 0.10 | 3.60 × 103 | 1.96 × 102 | 13.19 ± 0.19 | 6.54 × 105 |
| 3 CC | 23.62 ± 0.38 | 9.27 × 103 | 4.67 × 102 | 12.97 ± 0.03 | 7.62 × 105 |
| 4 CC | 27.45 ± 0.22 | - | - | 27.40 ± 0.23 | - |
| 5 CC | 28.1 ± 0.25 | - | - | 28.15 ± 0.17 | |
| 6 CC | 28.3 ± 0.19 | - | - | 29.91 ± 0.09 | |
| 7 CC | 28.58 ± 0.42 | - | - | 28.46 ± 0.23 | |
| 8 CC | 23.95 ± 0.05 | 7.22 × 103 | 3.71 × 102 | 18.89 ± 0.10 | 1.25 × 104 |
| 9 CC | 27.3 ± 0.19 | - | - | 22.81 ± 0.18 | 8.20 × 102 |
| 10 CC | 25.91 ± 0.14 | 1.78 × 103 | 9.59 × 101 | 19.25 ± 0.25 | 9.72 × 103 |
| 11 CC | 27.94 ± 0.18 | 27.72 ± 0. 28 | |||
| 12 CC | 27.58 ± 0.23 | - | - | 26.48 ± 0.09 | |
| 13 CC | 27.06 ± 0.10 | - | - | 26.68 ± 0.32 | |
| 14 CC | 27.07 ± 0.18 | - | - | 26.82 ± 0.12 | |
| 15 CC | 27.34 ± 0.25 | - | - | 26.74 ± 0.10 | |
| 16 CC | 28.05 ± 0.15 | - | - | 25.98 ± 0.08 | 9.07 × 101 |
| 17 CC | 25.94 ± 0.32 | 1.60 × 103 | 9.32 × 101 | 22.62 ± 0.32 | 9.29 × 102 |
| 18 CC | 30.83 ± 0.39 | - | - | 27.33 ± 0.34 | |
| 19 CC | 28.7 ± 0.31 | - | - | 26.96 ± 0.33 | |
| 20 CC | 28.41 ± 0.35 | - | - | 27.10 ± 0.32 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vizzini, P.; Vidic, J.; Manzano, M. Enrichment Free qPCR for Rapid Identification and Quantification of Campylobacter jejuni, C. coli, C. lari, and C. upsaliensis in Chicken Meat Samples by a New Couple of Primers. Foods 2021, 10, 2341. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10102341
Vizzini P, Vidic J, Manzano M. Enrichment Free qPCR for Rapid Identification and Quantification of Campylobacter jejuni, C. coli, C. lari, and C. upsaliensis in Chicken Meat Samples by a New Couple of Primers. Foods. 2021; 10(10):2341. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10102341
Chicago/Turabian StyleVizzini, Priya, Jasmina Vidic, and Marisa Manzano. 2021. "Enrichment Free qPCR for Rapid Identification and Quantification of Campylobacter jejuni, C. coli, C. lari, and C. upsaliensis in Chicken Meat Samples by a New Couple of Primers" Foods 10, no. 10: 2341. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10102341