
The Use of Winery by-Products to Enhance the Functional Aspects of the Fresh Ovine “Primosale” Cheese

 ,
,  , , , , , ,
, , , , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Grape Pomace Powder Production
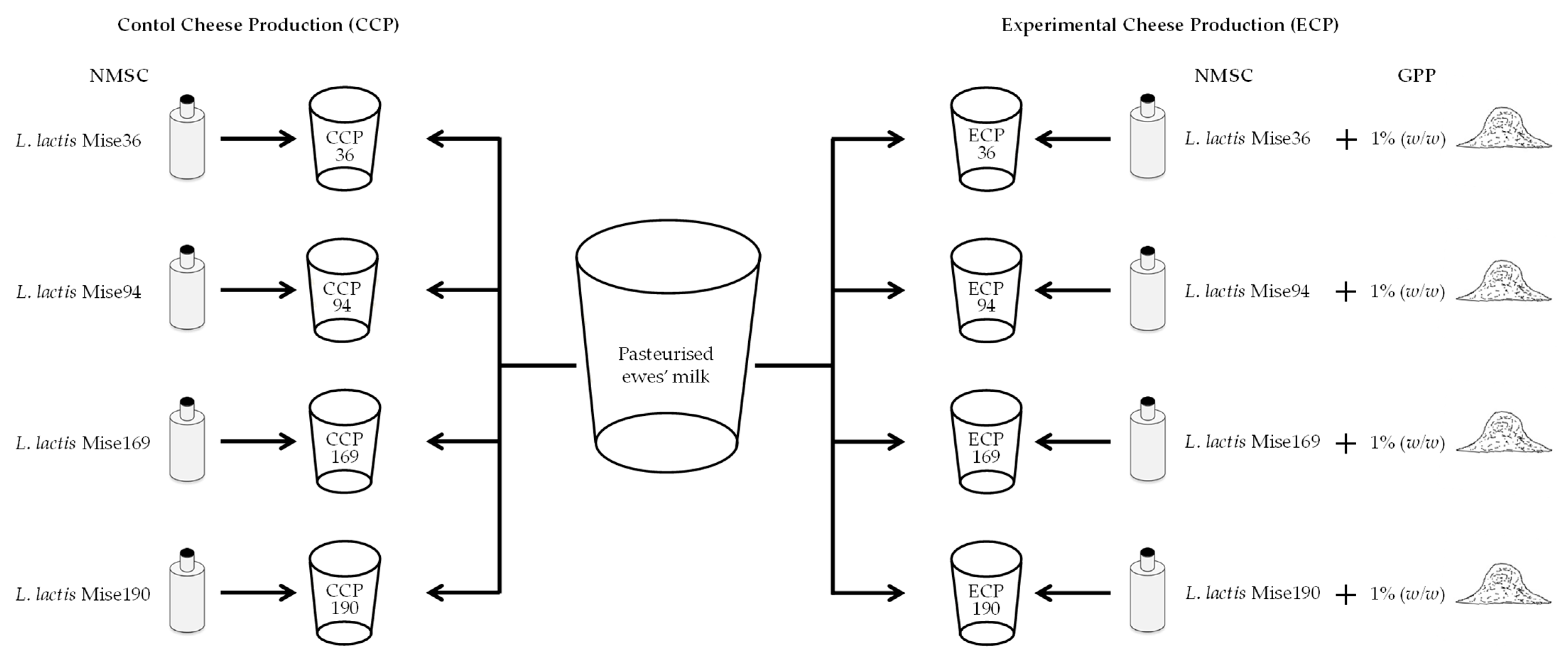
2.2. Strains and Development of Natural Milk Starter Cultures
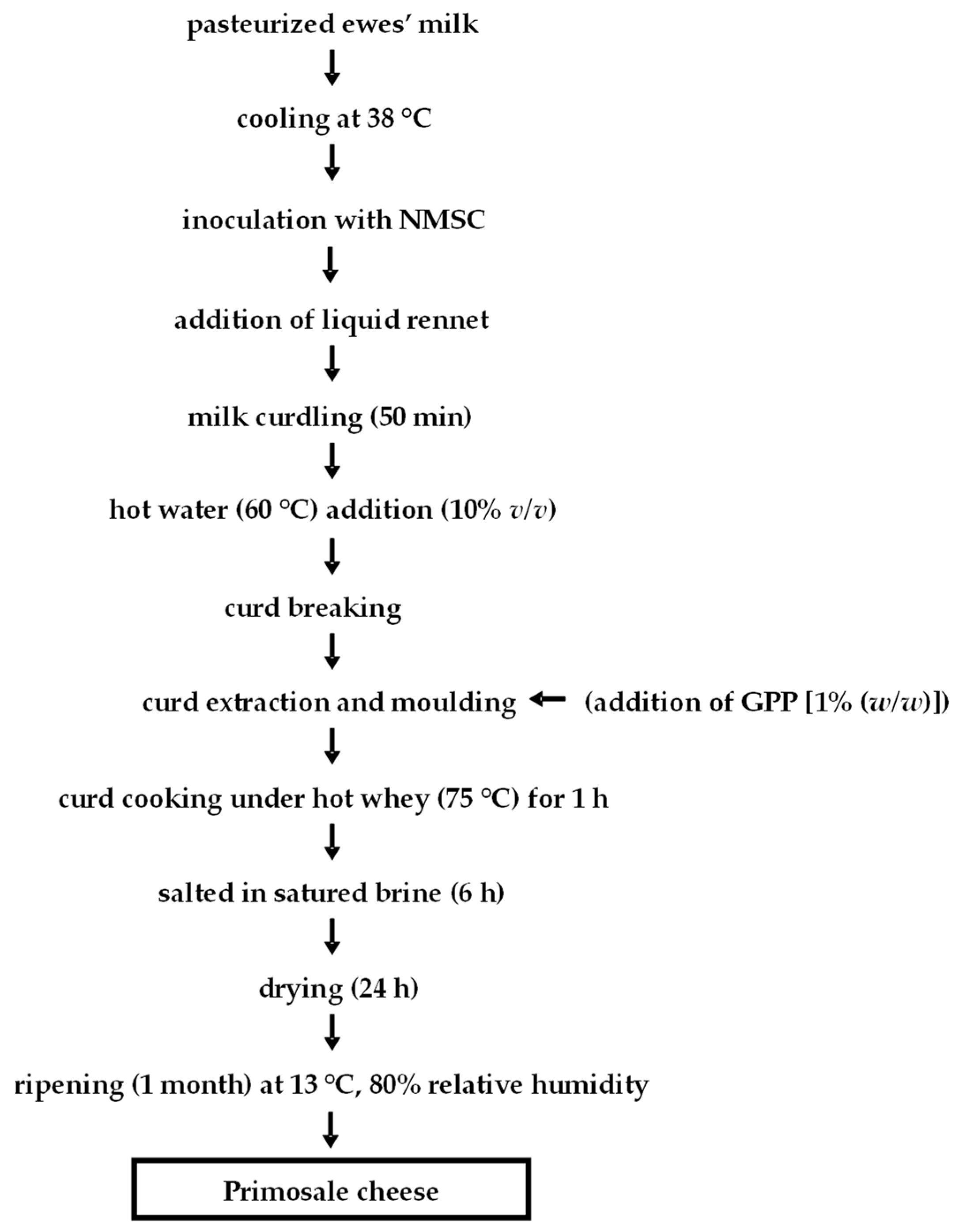
2.3. Experimental Cheese Production and Sample Collection
2.4. Microbiological Analyses
2.5. Phenotypic Grouping, Genotypic Differentiation and Identification of Thermoduric LAB
2.6. Persistence of the Added Strains
2.7. Physicochemical Analysis of Cheeses
2.8. Volatile Organic Compounds Emitted from Cheeses
2.9. Sensory Evaluation
2.10. Simulated Gastrointestinal Digestion
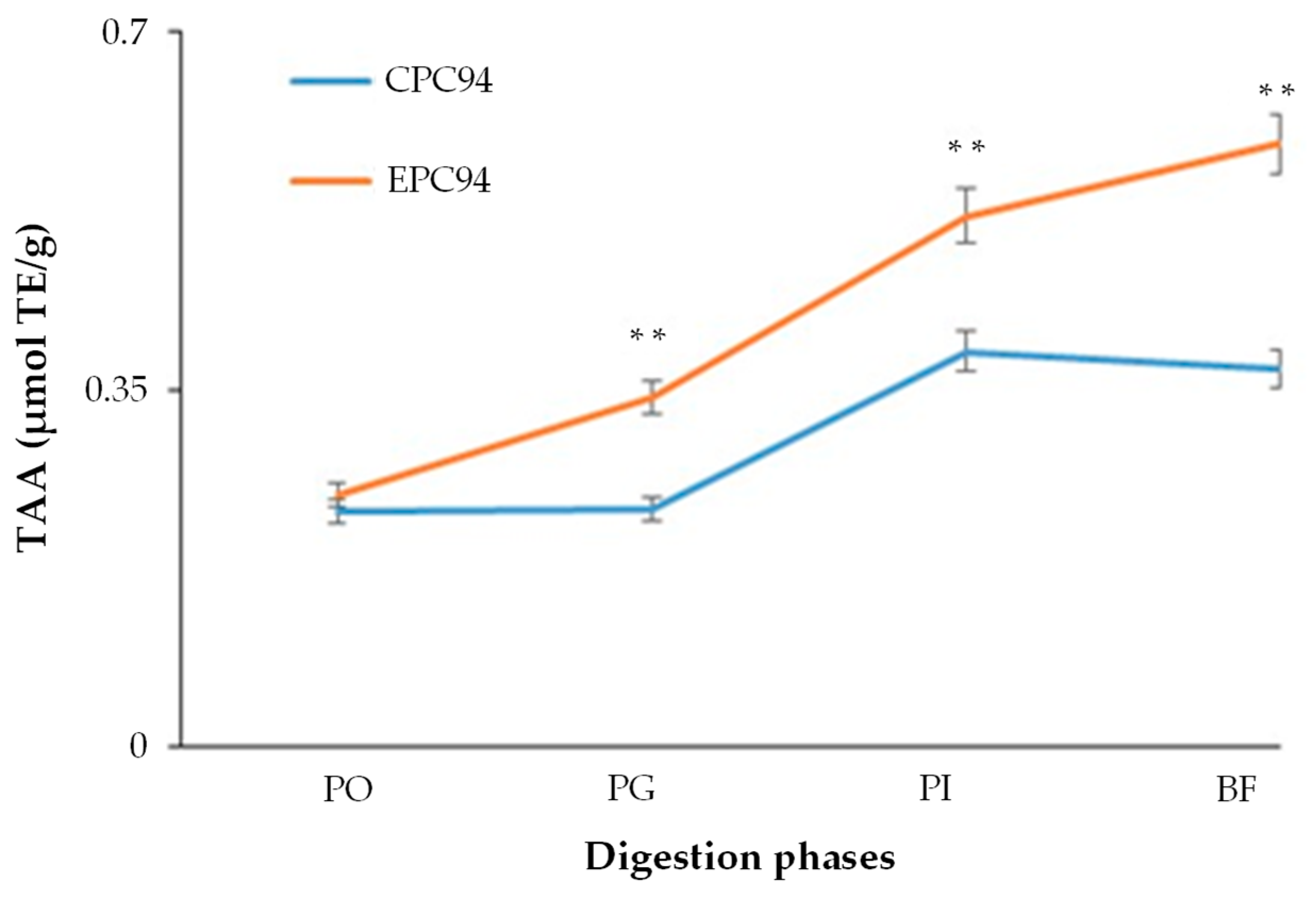
2.11. Total Antioxidant Activity
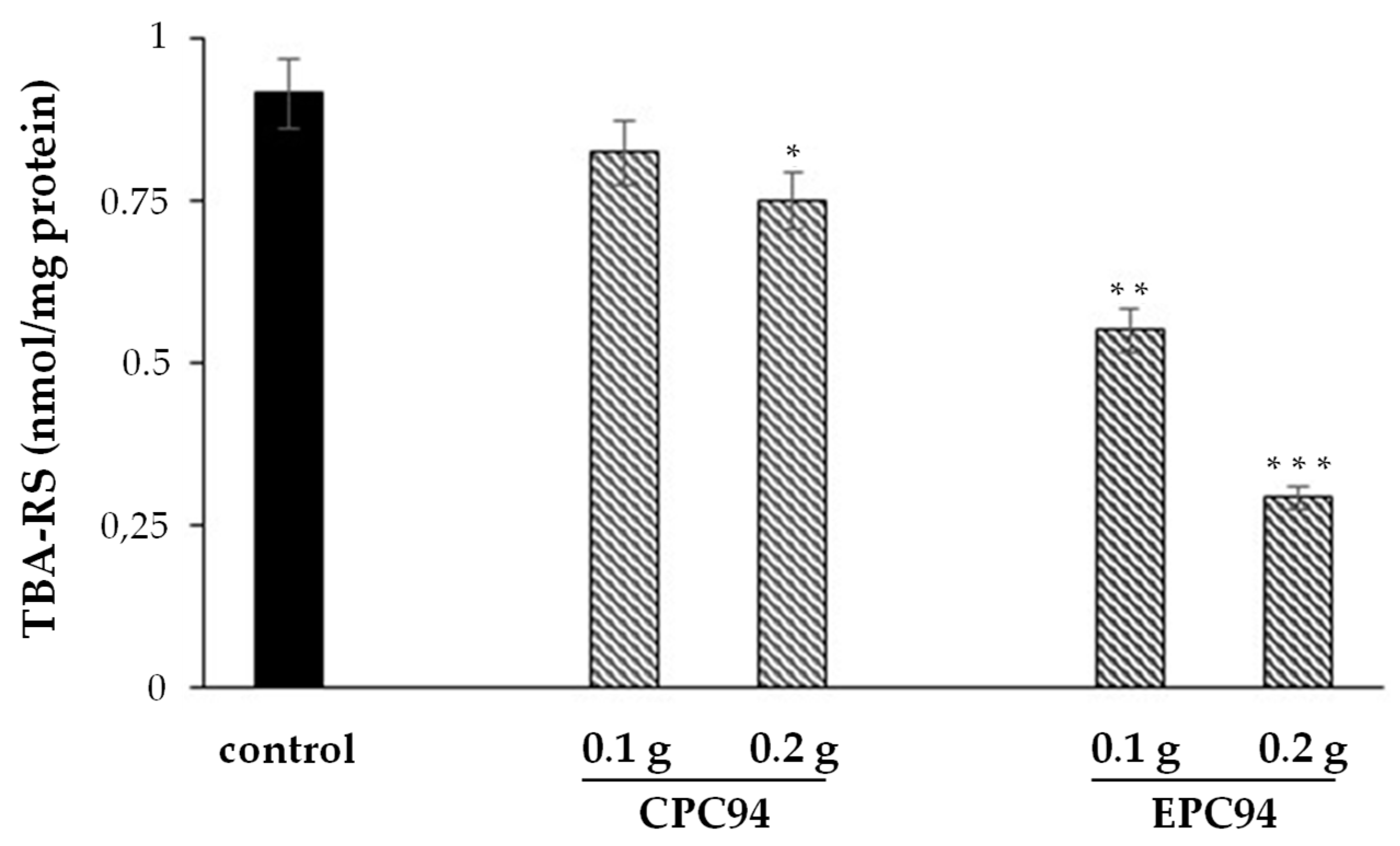
2.12. Membrane Lipid Peroxidation Assay
2.13. Statistical Analysis
3. Results and Discussion
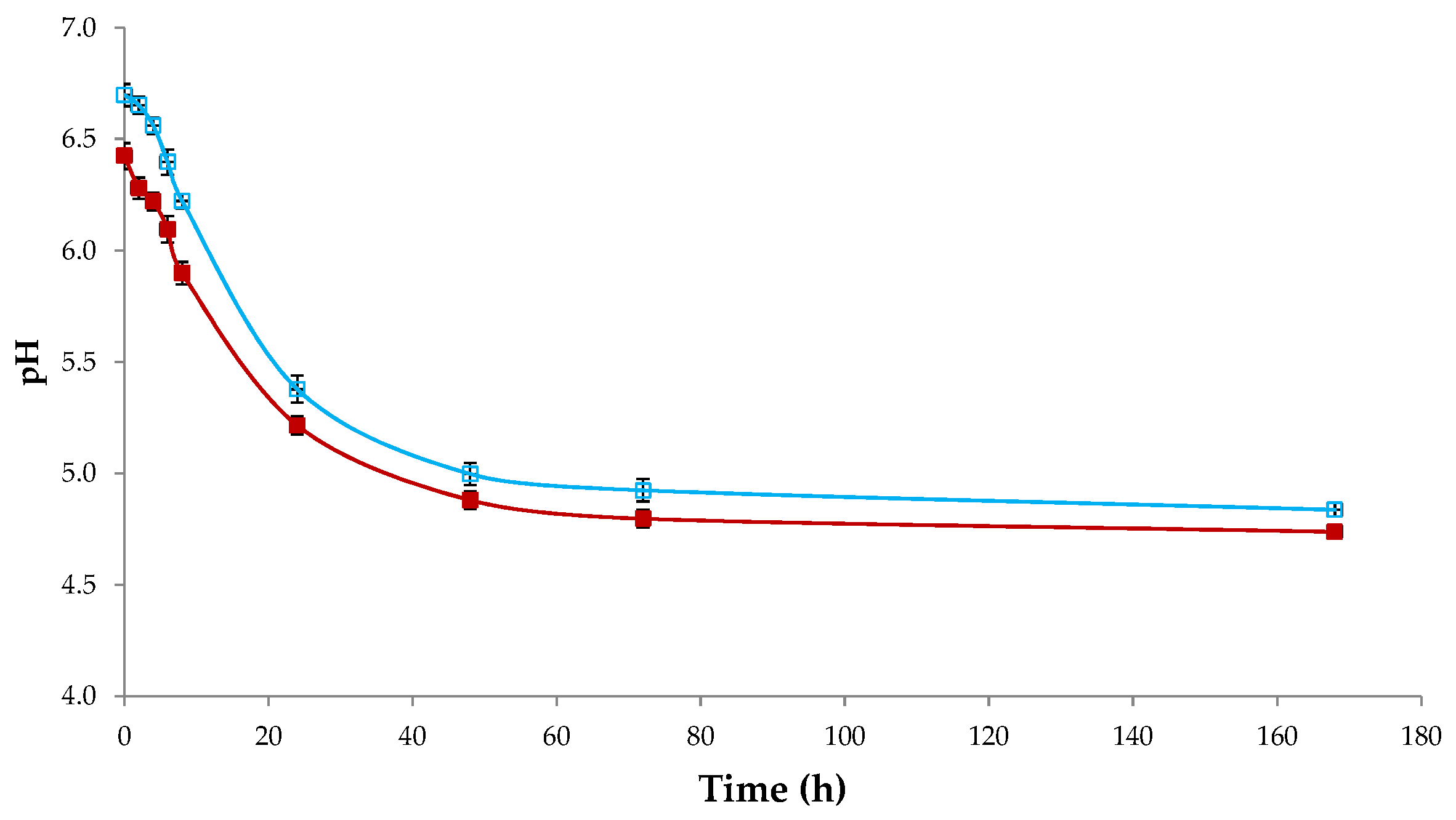
3.1. Acidification Kinetics
3.2. Microbiological Analyses
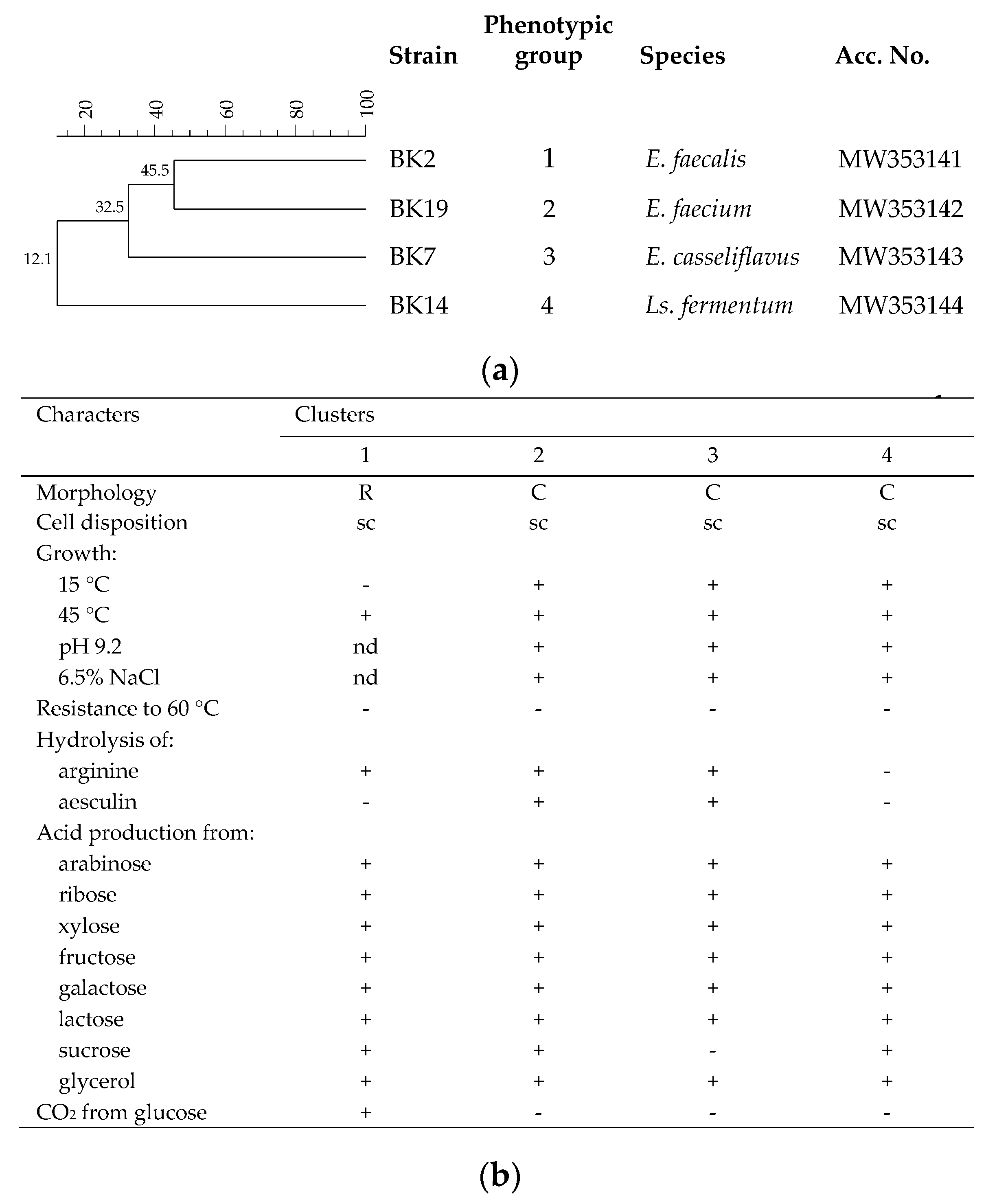
3.3. Identification of LAB Resistant to Pasteurization Process
3.4. Monitoring of the Added Strains
3.5. Physicochemical Analysis of Cheeses
3.6. Volatile Organic Compounds Composition of Cheeses
3.7. Sensory Test
3.8. Antioxidant Properties
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beres, C.; Costa, G.N.; Cabezudo, I.; da Silva-James, N.K.; Teles, A.S.; Cruz, A.P.; Mellinger-Silva, C.; Tonon, R.V.; Cabral, L.M.C.; Freitas, S.P. Towards integral utilization of grape pomace from winemaking process: A review. Waste Manag. 2017, 68, 581–594. [Google Scholar] [CrossRef] [PubMed]
- Ruggieri, L.; Cadena, E.; Martínez-Blanco, J.; Gasol, C.M.; Rieradevall, J.; Gabarrell, X.; Gea, T.; Sort, X.; Sánchez, A. Recovery of organic wastes in the Spanish wine industry. Technical, economic and environmental analyses of the composting process. J. Clean. Prod. 2009, 17, 830–838. [Google Scholar] [CrossRef] [Green Version]
- Fontana, A.R.; Antoniolli, A.; Bottini, R. Grape pomace as a sustainable source of bioactive compounds: Extraction, characterization, and biotechnological applications of phenolics. J. Agric. Food Chem. 2013, 61, 8987–9003. [Google Scholar] [CrossRef] [PubMed]
- Krishnaswamy, K.; Orsat, V.; Gariépy, Y.; Thangavel, K. Optimization of Microwave-Assisted Extraction of Phenolic Antioxidants from Grape Seeds (Vitis vinifera). Food Bioproc. Technol. 2013, 6, 441–455. [Google Scholar] [CrossRef]
- Kokkinomagoulos, E.; Kandylis, P. Sustainable Exploitation of By-Products of Vitivinicultural Origin in Winemaking. Proceedings 2020, 67, 5. [Google Scholar]
- Antonić, B.; Jančíková, S.; Dordević, D.; Tremlová, B. Grape Pomace Valorization: A Systematic Review and Meta-Analysis. Foods 2020, 9, 1627. [Google Scholar] [CrossRef]
- Food and Drug Administration (FDA). Agency Response Letter GRAS Notice No. GRN 000125. CFSAN/Office of Food Additive Safety. Available online: https://www.polyphenolics.com/wp-content/uploads/2015/08/Gras-2003.pdf (accessed on 11 January 2021).
- Mikovà, K. The regulation of antioxidants in foods. In Handbook of Food Preservation; Rahman, S., Ed.; Taylor & Francis Group: Milton Park, UK, 2007; pp. 83–267. [Google Scholar]
- García-Lomillo, J.; González-SanJosé, M.L. Applications of wine pomace in the food industry: Approaches and functions. Compr. Rev. Food Sci. Food Saf. 2017, 16, 3–22. [Google Scholar] [CrossRef]
- O’connell, J.E.; Fox, P.F. Significance and applications of phenolic compounds in the production and quality of milk and dairy products: A review. Int. Dairy J. 2001, 11, 103–120. [Google Scholar] [CrossRef]
- Shan, B.; Cai, Y.Z.; Brooks, J.D.; Corke, H. Potential application of spice and herb extracts as natural preservatives in cheese. J. Med. Food 2011, 14, 284–290. [Google Scholar] [CrossRef]
- Freire, F.C.; Adorno, M.A.T.; Sakamoto, I.K.; Antoniassi, R.; Chaves, A.C.S.D.; Dos Santos, K.M.O.; Sivieri, K. Impact of multi-functional fermented goat milk beverage on gut microbiota in a dynamic colon model. Food Res. Int. 2017, 99, 315–327. [Google Scholar] [CrossRef] [Green Version]
- Marchiani, R.; Bertolino, M.; Belviso, S.; Giordano, M.; Ghirardello, D.; Torri, L.; Piochi, M.; Zeppa, G. Yogurt enrichment with grape pomace: Effect of grape cultivar on physicochemical, microbiological and sensory properties. J. Food Qual. 2016, 39, 77–89. [Google Scholar] [CrossRef]
- Marchiani, R.; Bertolino, M.; Ghirardello, D.; McSweeney, P.L.; Zeppa, G. Physicochemical and nutritional qualities of grape pomace powder-fortified semi-hard cheeses. J. Food Sci. Technol. 2016, 53, 1585–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbaccia, P.; Francesca, N.; Gerlando, R.D.; Busetta, G.; Moschetti, G.; Gaglio, R.; Settanni, L. Biodiversity and dairy traits of indigenous milk lactic acid bacteria grown in presence of the main grape polyphenols. FEMS Microbiol. Lett. 2020, 367, fnaa066. [Google Scholar] [CrossRef] [PubMed]
- Cruciata, M.; Gaglio, R.; Scatassa, M.L.; Sala, G.; Cardamone, C.; Palmeri, M.; Moschetti, G.; La Mantia, T.; Settanni, L. Formation and characterization of early bacterial biofilms on different wood typologies applied in dairy production. Appl. Environ. Microbiol. 2018, 84, e02107-17. [Google Scholar] [CrossRef] [Green Version]
- ISO. Enumeration of Colony-Forming Units of Micro-Organisms—Colony-Count Technique at 30 Degrees C. Milk and Milk Products; International Standardization Organization (ISO): Geneva, Switzerland, 1992; Volume ISO 6610. [Google Scholar]
- Gaglio, R.; Francesca, N.; Di Gerlando, R.; Cruciata, M.; Guarcello, R.; Portolano, B.; Portolano, G.; Settanni, L. Identification, typing and investigation of the dairy characteristics of lactic acid bacteria isolated from “Vastedda della valle del Belìce” cheeses. Dairy Sci. Technol. 2014, 94, 157–180. [Google Scholar] [CrossRef] [Green Version]
- Gaglio, R.; Francesca, N.; Di Gerlando, R.; Mahony, J.; De Martino, S.; Stucchi, C.; Moschetti, G.; Settanni, L. Enteric bacteria of food ice and their survival in alcoholic beverages and soft drinks. Food Microbiol. 2017, 67, 17–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaglio, R.; Cruciata, M.; Di Gerlando, R.; Scatassa, M.L.; Mancuso, I.; Sardina, M.T.; Moschetti, G.; Portolano, B.; Settanni, L. Microbial activation of wooden vats used for traditional cheese production and evolution of the neo-formed biofilms. Appl. Environ. Microbiol. 2016, 82, 585–595. [Google Scholar] [CrossRef] [Green Version]
- CIE. Colorimetry; Commission Internationale de l’Eclairage (CIE): Vienna, Austria, 1986; Volume CIE 15.2. [Google Scholar]
- Bonanno, A.; Di Grigoli, A.; Vitale, F.; Di Miceli, G.; Todaro, M.; Alabiso, M.; Gargano, M.L.; Venturella, G.; Anike, F.N.; Isikhuemhenal, O.S. Effects of feeding diets supplemented with medicinal mushrooms myceliated grains on some production, health and oxidation traits of dairy ewes. Int. J. Med. Mushrooms 2019, 21, 89–103. [Google Scholar] [CrossRef] [PubMed]
- ISO. Sensory Analysis e General Guidance for the Design of Test. Rooms; International Standardization Organization (ISO): Geneva, Switzerland, 2007; Volume ISO 8589. [Google Scholar]
- Niro, S.; Fratianni, A.; Tremonte, P.; Sorrentino, E.; Tipaldi, L.; Panfili, G.; Coppola, R. Innovative Caciocavallo cheeses made from a mixture of cow milk with ewe or goat milk. J. Dairy Sci. 2014, 97, 1296–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, C.; Lucera, A.; Marinelli, V.; Del Nobile, M.A.; Conte, A. Influence of different by-products addition on sensory and physicochemical aspects of Primosale cheese. J. Food Sci. Technol. 2018, 55, 4174–4183. [Google Scholar] [CrossRef] [PubMed]
- Faccia, M.; Angiolillo, L.; Mastromatteo, M.; Conte, A.; Del Nobile, M.A. The effect of incorporating calcium lactate in the saline solution on improving the shelf life of fiordilatte cheese. Int. J. Dairy Technol. 2013, 66, 373–381. [Google Scholar]
- Attanzio, A.; Diana, P.; Barraja, P.; Carbone, A.; Spanò, V.; Parrino, B.; Cascioferro, S.M.; Allegra, M.; Cirrincione, G.; Tesoriere, L.; et al. Quality, functional and sensory evaluation of pasta fortified with extracts from Opuntia ficus-indica cladodes. J. Sci. Food Agric. 2019, 99, 4242–4247. [Google Scholar] [CrossRef]
- Miller, N.J.; Rice-Evans, C.A. Spectrophotometric determination of antioxidant activity. Redox Rep. 1996, 2, 161–171. [Google Scholar] [CrossRef]
- Pellegrini, N.; Ke, R.; Yang, M.; Rice-Evans, C. Screening of dietary carotenoids and carotenoid-rich fruit extracts for antioxidant activities applying 2,2-azinobis(3-ethylenebenzothiazoline- 6-sulfonic acid) radical cation decolorization assay. Meth. Enzymol. 1999, 299, 379–389. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Attanzio, A.; Tesoriere, L.; Allegra, M.; Livrea, M.A. Monofloral honeys by Sicilian black honeybee (Apis mellifera ssp. sicula) have high reducing power and antioxidant capacity. Heliyon 2016, 2, e00193. [Google Scholar] [PubMed] [Green Version]
- Mainente, F.; Menin, A.; Alberton, A.; Zoccatelli, G.; Rizzi, C. Evaluation of the sensory and physical properties of meat and fish derivatives containing grape pomace powders. Int. J. Food Sci. Technol. 2019, 54, 952–958. [Google Scholar] [CrossRef]
- Gaglio, R.; Todaro, M.; Scatassa, M.L.; Franciosi, E.; Corona, O.; Mancuso, I.; Di Gerlando, R.; Cardamone, C.; Settanni, L. Transformation of raw ewes’ milk applying “Grana” type pressed cheese technology: Development of extra-hard “Gran Ovino” cheese. Int. J. Food Microbiol. 2019, 307, 108277. [Google Scholar] [CrossRef] [PubMed]
- Settanni, L.; Gaglio, R.; Guarcello, R.; Francesca, N.; Carpino, S.; Sannino, C.; Todaro, M. Selected lactic acid bacteria as a hurdle to the microbial spoilage of cheese: Application on a traditional raw ewes’ milk cheese. Int. Dairy J. 2013, 32, 126–132. [Google Scholar] [CrossRef] [Green Version]
- Guarcello, R.; Carpino, S.; Gaglio, R.; Pino, A.; Rapisarda, T.; Caggia, C.; Marino, G.; Randazzo, C.L.; Settanni, L.; Todaro, M. A large factory-scale application of selected autochthonous lactic acid bacteria for PDO Pecorino Siciliano cheese production. Food Microbiol. 2016, 59, 66–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaglio, R.; Gentile, C.; Bonanno, A.; Vintaloro, L.; Perrone, A.; Mazza, F.; Barbaccia, P.; Settanni, L.; Di Grigoli, A. Effect of saffron addition on the microbiological, physicochemical, antioxidant and sensory characteristics of yoghurt. Int. J. Dairy Technol. 2019, 72, 208–217. [Google Scholar] [CrossRef]
- Rynne, N.M.; Beresford, T.P.; Kelly, A.L.; Guinee, T.P. Effect of milk pasteurisation temperature on age-related changes in lactose metabolism, pH and the growth of non-starter lactic acid bacteria in half-fat Cheddar cheese. Food Chem. 2007, 100, 375–382. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.; Harris, H.M.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 7, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- Settanni, L.; Moschetti, G. Non-starter lactic acid bacteria used to improve cheese quality and provide health benefits. Food Microbiol. 2010, 27, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Kopčáková, A.; Dubíková, K.; Šuľák, M.; Javorský, P.; Kmeť, V.; Lauková, A.; Pristaš, P. Restriction-modification systems and phage resistance of enterococci from ewe milk. LWT-Food Sci. Technol. 2018, 93, 131–134. [Google Scholar] [CrossRef]
- Gaglio, R.; Cruciata, M.; Scatassa, M.L.; Tolone, M.; Mancuso, I.; Cardamone, C.; Corona, O.; Todaro, M.; Settanni, L. Influence of the early bacterial biofilms developed on vats made with seven wood types on PDO Vastedda della valle del Belìce cheese characteristics. Int. J. Food Microbiol. 2019, 291, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Fusco, V.; Quero, G.M.; Poltronieri, P.; Morea, M.; Baruzzi, F. Autochthonous and probiotic lactic acid bacteria employed for production of “advanced traditional cheeses”. Foods 2019, 8, 412. [Google Scholar] [CrossRef] [Green Version]
- Todaro, M.; Francesca, N.; Reale, S.; Moschetti, G.; Vitale, F.; Settanni, L. Effect of different salting technologies on the chemical and microbiological characteristics of PDO Pecorino Siciliano cheese. Eur. Food Res. Technol. 2011, 233, 931–940. [Google Scholar] [CrossRef] [Green Version]
- Bonanno, A.; Di Grigoli, A.; Mazza, F.; De Pasquale, C.; Giosuè, C.; Vitale, F.; Alabiso, M. Effects of ewes grazing sulla or ryegrass pasture for different daily durations on forage intake, milk production and fatty acid composition of cheese. Animal 2016, 10, 2074–2082. [Google Scholar] [CrossRef] [Green Version]
- Frühbauerová, M.; Červenka, L.; Hájek, T.; Salek, R.N.; Velichová, H.; Buňka, F. Antioxidant properties of processed cheese spread after freeze-dried and oven-dried grape skin powder addition. Potravin. S. J. Food Sci. 2020, 14, 230–238. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, L.F.; Ribani, R.H.; Francisco, T.M.G.; Soares, A.A.; Pontarolo, R.; Haminiuk, C.W.I. Profile of bioactive compounds from grape pomace (Vitis vinifera and Vitis labrusca) by spectrophotometric, chromatographic and spectral analyses. J. Chromatogr. B 2015, 1007, 72–80. [Google Scholar] [CrossRef]
- Kırmacı, H.A.; Hayaloğlu, A.A.; Özer, H.B.; Atasoy, A.F.; Levent, O. Effects of Wild-Type Starter Culture (Artisanal Strains) on Volatile Profile of Urfa Cheese Made from Ewe Milk. Int. J. Food Prop. 2015, 18, 1915–1929. [Google Scholar] [CrossRef]
- Todaro, M.; Palmeri, M.; Cardamone, C.; Settanni, L.; Mancuso, I.; Mazza, F.; Scatassa, M.L.; Corona, O. Impact of packaging on the microbiological, physicochemical and sensory characteristics of a “pasta filata” cheese. Food Packag. Shelf Life 2018, 17, 85–90. [Google Scholar] [CrossRef]
- Virto, M.; Chavarri, F.; Bustamante, M.A.; Barron, L.J.R.; Aramburu, M.; Vicente, M.; Pérez-Elortondo, F.J.; Albisu, M.; de Renobales, M. Lamb rennet paste in ovine cheese manufacture. Lipolysis and flavour. Int. Dairy J. 2003, 13, 391–399. [Google Scholar] [CrossRef]
- Thierry, A.; Collins, Y.F.; Mukdsi, M.C.A.; McSweeney, P.L.H.; Wilkinson, M.G.; Spinnler, H.E. Lipolysis and metabolism of fatty acids in cheese. In Cheese: Chemistry, Physics and Microbiology; McSweeney, P.L.H., Fox, P.F., Cotter, P.D., Everett, D.W., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 423–444. [Google Scholar]
- Battelli, G.; Scano, P.; Albano, C.; Cagliani, L.R.; Brasca, M.; Consonni, R. Modifications of the volatile and nonvolatile metabolome of goat cheese due to adjunct of non-starter lactic acid bacteria. LWT-Food Sci. Technol. 2019, 116, 108576. [Google Scholar] [CrossRef]
- Fernandez-Garcia, E.; Gaya, P.; Medina, M.; Nunez, M. Evolution of the volatile components of raw ewe’s milk Castellano cheese: Seasonal variation. Int. Dairy J. 2004, 14, 39–46. [Google Scholar] [CrossRef]
- Guarrasi, V.; Sannino, C.; Moschetti, G.; Bonanno, A.; Di Grigoli, A.; Settanni, L. The individual contribution of starter and non-starter lactic acid bacteria to the volatile organic compound composition of Caciocavallo Palermitano cheese. Int. J. Food Microbiol. 2017, 259, 35–42. [Google Scholar] [CrossRef]
- Fox, P.F.; Guinee, T.P.; Cogan, T.M.; McSweeney, P.L.H. Biochemistry of cheese ripening. In Fundamentals of Cheese Science; Fox, P.F., Guinee, T.P., Cogan, T.M., McSweeney, P.L.H., Eds.; Springer: Berlin, Germany, 2020; pp. 391–442. [Google Scholar]
- Torri, L.; Piochi, M.; Marchiani, R.; Zeppa, G.; Dinnella, C.; Monteleone, E. A sensory-and consumer-based approach to optimize cheese enrichment with grape skin powders. J. Dairy Sci. 2016, 99, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Lucera, A.; Costa, C.; Marinelli, V.; Saccotelli, M.A.; Del Nobile, M.A.; Conte, A. Fruit and vegetable by-products to fortify spreadable cheese. Antioxidants 2018, 7, 61. [Google Scholar] [CrossRef] [Green Version]
- Hilario, M.C.; Puga, C.D.; Ocana, A.N.; Romo, F.P.G. Antioxidant activity, bioactive polyphenols in Mexican goats’ milk cheeses on summer grazing. J. Dairy Res. 2010, 77, 20–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helal, A.; Tagliazucchi, D.; Verzelloni, E.; Conte, A. Gastro-pancreatic release of phenolic compounds incorporated in a polyphenols-enriched cheese-curd. Food Sci. Technol. 2015, 60, 957–963. [Google Scholar] [CrossRef] [Green Version]
- Mattos, G.N.; Tonon, R.V.; Furtadob, A.A.; Cabralb, L.M.C. Grape by-product extracts against microbial proliferation and lipid oxidation: A review. J. Sci. Food Agric. 2017, 97, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Tagliazucchi, D.; Helal, A.; Verzelloni, E.; Conte, A. The type and concentration of milk increased the in vitro bioaccessibility of coffee chlorogenic acids. J. Agric. Food Chem. 2012, 60, 11056–11064. [Google Scholar] [CrossRef] [PubMed]
- Lucas, A.; Agabriel, C.; Martin, B.; Ferlay, A.; Verdier-Metz, I.; Coulon, J.B.; Rock, E. Relationships between the conditions of cow’s milk production and the contents in components of nutritional interest in raw milk farmhouse cheese. Lait 2006, 86, 177–202. [Google Scholar] [CrossRef]
- Gupta, A.; Mann, B.; Kumar, R.; Sangwan, R.B. Antioxidant activity of Cheddar cheeses at different stages of ripening. Int. J. Dairy Technol. 2009, 62, 339–347. [Google Scholar] [CrossRef]
- Ianni, A.; Martino, G. Dietary grape pomace supplementation in dairy cows: Effect on nutritional quality of milk and its derived dairy products. Foods 2020, 9, 168. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment (TR) | Starter Culture (NMSC) | SEM | Significance p < | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| MISE36 | MISE94 | MISE169 | MISE190 | TR | NMSC | TR*NMSC | |||||
| External color | lightness (L*) | Control | 62.24 | 62.17 | 61.15 | 60.96 | 64.70 | 2.34 | <0.0001 | 0.4294 | 0.3086 |
| Experimental | 31.40 | 26.93 | 33.39 | 33.21 | 32.07 | ||||||
| Total a | 44.55 | 47.27 | 47.08 | 48.38 | |||||||
| redness (a*) | Control | −5.82 | −5.82 | −5.99 | −5.79 | −5.68 | 0.26 | <0.0001 | 0.5537 | 0.6203 | |
| Experimental | 1.77 | 2.08 | 1.71 | 1.43 | 1.86 | ||||||
| Total | −1.87 | −2.14 | −2.18 | −1.91 | |||||||
| yellowness (b*) | Control | 13.64 | 13.57 | 14.04 | 12.54 | 14.40 | 0.47 | <0.0001 | 0.1419 | 0.3863 | |
| Experimental | 1.82 | 1.73 | 1.89 | 1.67 | 1.97 | ||||||
| Total | 7.65 | 7.97 | 7.11 | 8.19 | |||||||
| Internal color | lightness (L*) | Control | 70.39 | 71.31 | 70.47 | 69.16 | 70.61 | 1.46 | <0.0001 | 0.3541 | 0.7342 |
| Experimental | 43.22 | 42.70 | 43.72 | 41.35 | 45.13 | ||||||
| Total | 57.00 | 57.10 | 55.25 | 57.87 | |||||||
| redness (a*) | Control | −4.90 | −4.95 | −5.20 | −4.62 | −4.82 | 0.36 | <0.0001 | 0.2181 | 0.6629 | |
| Experimental | 3.45 | 3.12 | 3.37 | 4.13 | 3.16 | ||||||
| Total | −0.91 | −0.91 | −0.25 | −0.83 | |||||||
| yellowness (b*) | Control | 12.60 | 13.04 | 13.82 | 11.53 | 12.01 | 0.68 | <0.0001 | 0.2068 | 0.4600 | |
| Experimental | 3.67 | 4.09 | 3.61 | 3.27 | 3.69 | ||||||
| Total | 8.57 | 8.71 | 7.40 | 7.85 | |||||||
| Hardness, N/mm2 | Control | 1.03 | 1.01 | 0.98 | 1.10 | 1.02 | 0.048 | <0.0001 | 0.0906 | 0.4099 | |
| Experimental | 1.23 | 1.31 | 1.15 | 1.26 | 1.19 | ||||||
| Total | 1.16 | 1.06 | 1.18 | 1.11 | |||||||
| Chemical composition | Dry matter (DM), % | Control | 63.23 | 63.19 | 62.75 | 63.46 | 63.51 | 0.48 | 0.2497 | 0.8107 | 0.6814 |
| Experimental | 63.65 | 64.05 | 63.59 | 63.49 | 63.49 | ||||||
| Total | 63.62 | 63.17 | 63.47 | 63.50 | |||||||
| Ash, % DM | Control | 6.37 | 6.32 | 6.22 | 6.51 | 6.45 | 0.21 | 0.5420 | 0.8912 | 0.7650 | |
| Experimental | 6.47 | 6.62 | 6.43 | 6.43 | 6.40 | ||||||
| Total | 6.47 | 6.33 | 6.47 | 6.42 | |||||||
| Protein, % DM | Control | 45.45 | 45.43 | 43.55 | 46.62 | 46.61 | 1.44 | 0.0147 | 0.3339 | 0.7330 | |
| Experimental | 48.72 | 49.56 | 47.64 | 50.19 | 47.50 | ||||||
| Total | 47.49 | 45.59 | 48.40 | 46.85 | |||||||
| Fat, % DM | Control | 45.69 | 46.31 | 46.06 | 44.52 | 45.87 | 1.03 | 0.0002 | 0.2815 | 0.6349 | |
| Experimental | 40.63 | 39.71 | 41.64 | 39.35 | 41.83 | ||||||
| Total | 43.01 | 43.85 | 41.93 | 43.85 | |||||||
| TBARS, μg MDA/kg DM | Control | 31.49 | 33.30 | 31.47 | 31.25 | 29.94 | 3.8123 | 0.0173 | 0.5980 | 0.4708 | |
| Experimental | 38.41 | 39.46 | 37.45 | 32.98 | 43.75 | ||||||
| Total | 36.38 | 34.46 | 32.11 | 36.85 | |||||||
| Chemical Compounds a | Samples | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| GPP | CPC36 | EPC36 | CPC94 | EPC94 | CPC169 | EPC169 | CPC190 | EPC190 | |
| Acids | |||||||||
| Acetic acid | n.d. | 7.4 | 15.6 | 13.6 | 16.8 | 12.6 | 12.6 | 11.1 | 13.9 |
| Butanoic acid | n.d. | 4.1 | 10.6 | 12.5 | 7.3 | 7.3 | 7.3 | 6.0 | 9.9 |
| 4-Hydroxybutanoic acid | 4.1 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Hexanoic acid | 1.6 | 1.9 | 4.6 | 5.7 | 3.4 | 5.8 | 4.2 | 3.6 | 4.8 |
| Pentanoinc acid-2-hydroxy-4-methyl | n.d. | 1.7 | 3.4 | 1.4 | 1.9 | 1.4 | 1.2 | 1.2 | 1.2 |
| Nonanoic acid | 3.4 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| ketones | |||||||||
| 2-Pentanone | n.d. | 2.2 | 0.8 | 1.5 | 0.9 | 2.7 | 0.8 | 2.7 | 0.8 |
| 3-Hydroxy-2-butanone | n.d. | 23.8 | 41.8 | 31.1 | 49.6 | 7.6 | 22.3 | 14.0 | 34.6 |
| 2-Heptanone | n.d. | 0.2 | 0.4 | 0.8 | 0.4 | 0.5 | 0.6 | 1.3 | 0.3 |
| p-Phenylacetophenone | 4.2 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Alcohol | |||||||||
| Isoamyl alcohol | 4.9 | 7.8 | 2.7 | 6.7 | 8.2 | 27.2 | 37.7 | 10.4 | 15.0 |
| 2-Butanol | n.d. | 0.7 | 2.6 | 4.8 | 2.1 | 2.5 | 2.6 | 1.7 | 4.1 |
| 2-Phenylethanol | 11.3 | n.d. | 1.4 | n.d. | 1.0 | n.d. | 1.5 | n.d. | 0.9 |
| Hydrocarbons | |||||||||
| Hexane 2-methyl | n.d. | 3.0 | 0.7 | 1.0 | 0.6 | 2.2 | 0.7 | 2.6 | 0.8 |
| Heptane 2,4-dimethyl | 3.2 | 2.0 | 2.7 | 3.0 | 3.2 | 2.6 | 1.4 | 1.6 | 3.2 |
| Nonane | 2.2 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Nonane 2,5-methyl | 2.3 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Decane | 1.8 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Dodecane | 2.3 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Hexadecane | 1.7 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Aldeyde | |||||||||
| Hexanal | 3.2 | 22.0 | 2.8 | 5.6 | 0.2 | 18.1 | 1.0 | 15.8 | 2.4 |
| Heptanal | n.d. | 21.8 | 4.4 | 6.6 | 0.5 | 6.6 | 1.5 | 25.0 | 2.6 |
| Nonanal | 1.7 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Monoterpene | |||||||||
| D-Limonene | 6.3 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| α-Pinene | 2.1 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Carene | 1.5 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Esters | |||||||||
| Octanoinc acid, ethyl ester | 9.6 | n.d. | 1.3 | n.d. | 1.1 | n.d. | 1.2 | n.d. | 0.7 |
| Butanedioic acid, diethyl ester | 2.2 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Decanoic acid, ethyl ester | 9.7 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Diol | |||||||||
| 2,3-Butanediol | 20.6 | 1.4 | 4.1 | 5.6 | 2.9 | 2.9 | 3.4 | 2.8 | 4.9 |
| Attributes | Trial | SEM | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CPC36 | EPC36 | CPC94 | EPC94 | CPC169 | EPC169 | CPC190 | EPC190 | Judges | Cheeses | ||
| Intensity of odor | 5.32 cd | 5.94 bc | 6.22 b | 6.78 a | 5.06 d | 5.76 bcd | 5.26 cd | 5.91 bc | 0.07 | 0.053 | <0.0001 |
| Intensity of aroma | 5.41 cde | 5.95 b | 5.99 b | 6.55 a | 4.95 e | 5.46 cd | 5.16 de | 5.82 bc | 0.06 | 0.099 | <0.0001 |
| Sweet | 5.02 b | 4.47 c | 5.57 a | 5.05 b | 5.19 b | 4.71 c | 5.15 b | 4.62 c | 0.04 | 0.627 | <0.0001 |
| Salt | 3.53 a | 3.48 a | 3.43 a | 3.46 a | 3.47 a | 3.45 a | 3.44 a | 3.46 a | 0.03 | 0.999 | 0.999 |
| Acid | 2.42 b | 3.28 a | 2.44 b | 3.22 a | 2.37 b | 3.20 a | 2.48 b | 3.32 a | 0.05 | 0.733 | <0.0001 |
| Astringent | 0.00 b | 1.66 a | 0.00 b | 1.64 a | 0.00 b | 1.60 a | 0.00 b | 1.59 a | 0.08 | 0.999 | <0.0001 |
| Friability | 1.53 b | 2.42 a | 1.50 b | 2.46 a | 1.42 b | 2.31 a | 1.56 b | 2.33 a | 0.05 | 0.860 | <0.0001 |
| Fiber | 1.39 b | 2.56 a | 1.30 b | 2.48 a | 1.36 b | 2.56 a | 1.42 b | 2.58 a | 0.06 | 0.952 | <0.0001 |
| Adhesiveness | 2.41 b | 3.49 a | 2.45 b | 3.54 a | 2.38 b | 3.42 a | 2.46 b | 3.44 a | 0.05 | 0.998 | <0.0001 |
| Hardness | 4.18 a | 2.45 b | 4.05 a | 2.56 b | 4.02 a | 2.46 b | 4.06 a | 2.38 b | 0.08 | 0.985 | <0.0001 |
| Humidity | 2.53 b | 3.62 a | 2.40 b | 3.51 a | 2.28 b | 3.56 a | 2.60 b | 3.65 a | 0.06 | 0.971 | <0.0001 |
| Overall assessment | 4.36 c | 4.56 c | 5.74 b | 6.07 a | 4.33 c | 4.36 c | 4.46 c | 4.62 c | 0.06 | 0.999 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaglio, R.; Barbaccia, P.; Barbera, M.; Restivo, I.; Attanzio, A.; Maniaci, G.; Di Grigoli, A.; Francesca, N.; Tesoriere, L.; Bonanno, A.; et al. The Use of Winery by-Products to Enhance the Functional Aspects of the Fresh Ovine “Primosale” Cheese. Foods 2021, 10, 461. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10020461
Gaglio R, Barbaccia P, Barbera M, Restivo I, Attanzio A, Maniaci G, Di Grigoli A, Francesca N, Tesoriere L, Bonanno A, et al. The Use of Winery by-Products to Enhance the Functional Aspects of the Fresh Ovine “Primosale” Cheese. Foods. 2021; 10(2):461. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10020461
Chicago/Turabian StyleGaglio, Raimondo, Pietro Barbaccia, Marcella Barbera, Ignazio Restivo, Alessandro Attanzio, Giuseppe Maniaci, Antonino Di Grigoli, Nicola Francesca, Luisa Tesoriere, Adriana Bonanno, and et al. 2021. "The Use of Winery by-Products to Enhance the Functional Aspects of the Fresh Ovine “Primosale” Cheese" Foods 10, no. 2: 461. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10020461