
Pseudomonas spp.: Are Food Grade Organic Acids Efficient against These Spoilage Microorganisms in Fresh Cheeses?



Department of Health, Animal Science and Food Safety, University of Milan, Via dell’ Università 6, 26900 Lodi, Italy
*
Author to whom correspondence should be addressed.
Foods 2021, 10(4), 891; https://0-doi-org.brum.beds.ac.uk/10.3390/foods10040891
Submission received: 10 March 2021
/
Revised: 1 April 2021
/
Accepted: 15 April 2021
/
Published: 19 April 2021
(This article belongs to the Special Issue Dairy Spoilage Microorganisms, Mechanisms and Novel Preservation Strategies)
Abstract
:Psychrotolerant Pseudomonas spp. are among the most common spoilage agents in fresh, soft and semi-soft cheeses; therefore, hurdles inhibiting their growth are in strong demand by producers. This study aimed to establish Minimal Inhibiting Concentrations (MICs) of lactic and acetic acid towards P. fluorescens and to evaluate the efficacy of a cheese surface treatment with these two organic acids. MICs were determined in Brain Heart Infusion broth at 30 °C: the inhibition was achieved at a concentration of 49.96 mM and 44.40 mM of acetic and lactic acid, respectively. Two series of inhibition tests were performed on fresh “Primo sale” cheese, inoculated with P. brenneri MGM3, then dipped into different acid solutions (acetic acid: 49.96, 99.92 and 149.88 mM; lactic acid: 44.40, 88.80 and 133.20 mM) and stored at 6 °C. P. brenneri MGM3 were enumerated, including a control series. A significantly lower growth was revealed at the highest concentrations tested, both for acetic (p < 0.01) and lactic acid (p < 0.05) if compared to control samples. A conditioning of “Primo sale” surface with organic acid solutions could be a useful hurdle for Pseudomonas inhibition and shelf-life extension; it should be applied in combination with other mild interventions to fight spoilage and maintain the original product characteristics.
1. Introduction
A large proportion of food gets spoiled before reaching the end consumer. According to the Food and Agriculture Organization of the United Nations, one-third of the food produced for human consumption is spoiled or wasted [1]. Spoilage of food for human consumption is a world-wide issue that is of particular interest for stakeholders, food producers and consumers. Food spoilage could be defined as a loss of quality in terms of colour, odour, texture and in general a loss in sensorial characteristics [2], and could be attributed to a microbiological, chemical or physical source [3]. Food spoilage is mainly caused by microorganisms that can rapidly colonize and replicate on fresh food. Psychrotolerant Pseudomonas spp. are among the most common bacteria implicated in spoilage, especially of refrigerated food with a prolonged shelf life, where they are likely selected [4,5,6]. Pseudomonas spp. are aerobic, Gram-negative, non-spore-forming bacteria; some strains produce pigments, i.e., the yellow-green fluorescing pigment called pyoverdine (especially some P. fluorescens and P. aeruginosa strains) and the blue-green pigment called pyocyanin [7,8,9]. Pseudomonas spp. produce thermotolerant lipolytic and proteolytic enzymes that reduce the quality and the shelf-life of raw and processed milk [10]. These bacteria are inactivated by the thermal processes currently applied through the production of most dairy products, but they can also enter the food production chain as post-process contaminants, due to the contact of the final product with soil, water or raw material [11]. Pseudomonads are also recognized to be able to colonize environmental production as well as equipment and facilities for long periods, thanks to their ability to produce persistent biofilms [12,13].
Dairy products, and especially fresh cheeses made by enzymatic coagulation, very often get spoiled by microorganisms, owing to their high water activity and to the neutral pH, with P. fluorescens group recognized as a frequent cause of these alterations [14]. The reduction of these spoilage organisms in pasteurized dairy products is critical for extending their shelf life [15]. In this context, the use of weak organic acids may be convenient. The possibility for the undissociated form of these compounds to pass the bacterial cell membrane causes an alteration of the internal balance of bacterial cells: the decreased pH within the cytoplasm, due to the dissociation of the organic acid, requires an energy expense of the cell to pump out hydrogen ions, trying to re-establish the internal pH [16]. The addition of organic acids is allowed by the European Union legislation [17], and it may be suitable for many Ready to eat (RTE) foods [18], including fresh cheese, with the aim to reformulate the products leading to a substrate less permissive for bacterial replication.
The first aim of the present study was to establish Minimal Inhibiting Concentrations (MICs) of lactic and acetic acid towards P. brenneri MGM3, a member of P. fluorescens subgroup [19,20], isolated from a fresh cheese matrix and cultured in broth. Secondly, challenge tests with P. brenneri inoculation onto the surface of Primo sale cheese, an Italian fresh cheese made from pasteurized milk, were conducted: the ability of cheese dipping treatments with lactic or acetic acid in limiting the bacterial growth was evaluated.
2. Materials and Methods
2.1. MICs Determination
A strain of P. brenneri MGM3, previously isolated from a fresh dairy product (Mozzarella cheese) and identified by MALDI-TOF (Matrix-Assisted Laser Desorption-Ionisation-Time of Flight Mass Spectrometry) at the microbiology laboratory of the Department of Veterinary Medicine, University of Milan (Addis, personal communication) was used for this trial. The strains stock was kept frozen at −80 °C in Microbank Cryogenic vials (Pro-Lab Diagnostics U.K., Merseyside, UK). From the stock culture, a loopful was transferred to Brain Heart Infusion broth (BHI) (Oxoid, Basingstoke, UK) with pH 7.2, and incubated at 30 °C for 48 h. The culture was harvested in late exponential growth phase, defined as a relative change in Optical Density (OD) of 0.05–0.2 at 540 nm at 540 nm determined with a spectrophotometer (Jenway 6105, Staffordshire, UK). Cell concentrations of this pre-culture was determined by microscopy at 1000× magnification (Motic, B310, Wetzlar, Germany), considering that one cell per field of view corresponded to a concentration around 106 CFU/mL [21].
Ten millilitres tubes containing Brain Hear Infusion (BHI) broth added with aliquots of acetic acid (code 1005706, Sigma Aldrich, Steinheim, Germany) or lactic acid (code 252476, Sigma Aldrich) were prepared for the inoculation. The following final concentrations were obtained: 2.50 mM, 5.00 mM, 12.49 mM, 24.98 mM and 49.96 mM for acetic acid, and 2.78 mM, 5.55 mM, 11.10 mM, 22.20, mM and 44.40 mM for lactic acid; a control series (CTRL: BHI broth) was also prepared. After adjustment of bacterial concentration, aliquots of 0.1 mL of P. brenneri suspension were inoculated into the broths, achieving a final concentration of 100 CFU/mL. At the time of inoculation (t0), OD was measured, and the tubes were then incubated at 30 °C in duplicate. At fixed times (24, 48, 72, 96 h from inoculation), OD was newly measured. Blank (non-inoculated) BHI broth samples were prepared for each series and incubated in the same conditions; these samples were used for the comparison of the respective inoculated samples. At t0, the pH of the broths was also measured.
2.2. Challenge Tests
Specific challenge tests with Primo sale cheese, a fresh cheese made from cows’ milk produced and immediately marketed, were carried out to evaluate the effect of different concentrations of acetic and lactic acid on P. brenneri MGM3 inoculated on a real cheese matrix. The concentrations tested were set taking into account the MICs determined in Section 2.1.
Primo sale cheese samples (200 g) were taken on the first day after production. Before inoculation, the cheese was sliced, obtaining standard 8 g—weight pieces.
P. brenneri MGM3 stock was kept frozen at −80 °C in Microbank Cryogenic vials; from the culture, a loop was transferred to BHI and incubated at 30 °C for 48 h. In order to pre-adapt the cells to the environmental conditions of the challenge tests, the cultures were subsequently re-inoculated in BHI broth and then incubated at 6 °C. The culture was harvested as described in a previous section. Finally, the culture was diluted in sterile saline water (0.85% NaCl) and spread onto the surface of the cheese to obtain the starting concentration around 2 Log CFU/g. To minimize changes in product characteristics, the inoculum volume did not exceed 1% of the weight of the samples.
P. brenneri-inoculated samples were then divided into five series, four of which were dipped in the different acid solutions (acetic acid: Ac1 = 49.96 mM, Ac2 = 99.92 mM; lactic acid: Lac1 = 44.40 mM, Lac2 = 88.81 mM); the samples of the fifth series (control, CTRL) were dipped in sterile water. All the inoculated samples were then incubated at 6 °C; during storage, temperature was recorded by data loggers (Escort iLog, Escort Data Logging System Ltd., Aesch Bei Birmensdorf, Switzerland) and analysed in triplicate for Pseudomonas spp. enumeration and pH determination. The sampling times were set at t0 and after 3 h (aiming to verify the eventual microbial inactivation due to the contact with the acid solution), 48 and 96 h from inoculation. At each sampling time, the whole cheese aliquot (8 g) was 10-fold diluted in pre-chilled sterile saline and homogenized for 60 s in a peristaltic blender BagMixer® 400 W (Interscience, Saint Nom, France). Further appropriate 10-fold dilutions of the homogenates were made with pre-chilled (maintained at refrigeration temperatures) sterile saline. P. brenneri was enumerated by spread plating on Pseudomonas agar base added Cephalothin, Fucidin, Cetrimide (CFC) Supplement (Scharlab, Barcelona, Spain) and incubated at 30 °C for 48 h. An increase of +0.5 Log CFU/g was used to discriminate growth and no growth in the product. The surface pH of each cheese sample was measured with a pH meter equipped with a penetration electrode in triplicate (Amel, Milan, Italy), in order to evaluate the environmental condition of the specific niche where the microorganisms were inoculated (Amel, Milan, Italy).
Based on the results obtained from the first trial, a second challenge test was performed on Primo sale cheese samples; the same protocol described above was applied, with some modifications. Higher organic acid concentrations were tested, namely Ac3 (acetic acid, 149.87 Mm) and Lac3 (lactic acid, 133.2 mM); as for the previous challenge test, a control series (CTRL) was included. All the inoculated samples were then incubated at 6 °C and analysed in triplicate at t0 and after 3, 24 and 48 h from inoculation.
2.3. Statistical Analysis
Data obtained from Pseudomonas counts (expressed as Log CFU/g) were submitted to a two-way univariate Analysis of Variance (ANOVA) in SAS (version 9.1, 2016; SAS Institute Inc., Cary, NC, USA) to reveal potential differences in the treatments. Threshold values for statistical significance were set at p < 0.05 and p < 0.01.
3. Results and Discussion
In dairy industry, psychrotrophic microorganisms are predominant: among all, Pseudomonas spp. can represent till 75% of the total bacterial count of milk [5,22]. These unwanted microorganisms are responsible for spoilage of dairy products (in fact they dominate the microflora at the end of the shelf-life), acting a crucial role with negative consequences on the quality and shelf life of processed milk [5,10,23]. Pseudomonas are reduced in milk by heat treatments and hygiene plans, but post-pasteurization contamination is frequent and difficult to avoid in a dairy processing plant. Fresh cheeses, like Mozzarella or Primo sale cheese, are frequently involved in Pseudomonas spp. contamination [9,11,24,25]. Bioprotective cultures, EDTA-Na2 and lysozyme have been previously studied to evaluate their antagonistic activity against Pseudomonas spp. [6,26], but very limited literature is available on the effect of organic acids especially when applied to fresh cheese. Organic acids, according to Reg. EU 1333 (2008) [17] are allowed for addition to several food categories with the quantum satis principle; thus, the estimation of MIC is particularly important to set the minimum effective concentration to be applied.
In the present study, MICs for lactic and acetic acid against P. brenneri MGM3 growth were calculated in order to identify the growth boundary (the concentration separating growth/no-growth conditions for organic acid concentrations). The results are reported in Table 1. Our experiments highlighted that, at 30 °C, BHI with a concentration of acetic acid of 49.96 mM did not support P. brenneri MGM3 growth within 72 h; the same effect was achieved with a concentration of lactic acid of 44.40mM. In recent years, some studies reported the MIC of lactic and acetic acid for pseudomonads, showing variable results. The MIC value obtained in our study for acetic acid was higher than those reported by previous studies: Cruz-Romero et al. [27] investigated the antimicrobial activity of chitosan, organic acids and nano-sized solubilisates for potential use in smart antimicrobial-active packaging for food applications, revealing a MIC of 16.7 mM for acetic acid. These data were in agreement with the study by Moreira et al. [28] who found a MIC of 0.78 µL/mL (equal to 13.63 mM) towards P. aeruginosa; a lower value was observed by Nakai and Siebert [29], with a MIC of 0.346 g/L (5.8 mM). Bushell et al. [30] tested P. aeruginosa at 0, 5, 10, 20 mM of acetic acid at different pH, finding the MIC at the concentration of 20 mM at pH 5. Also, considering lactic acid, very different results were obtained by various authors: Moreira et al. [28] detected a higher MIC value (6.25 µL/mL, equal to 83.88 mM), whereas lower values were obtained by Nakai and Siebert (0.630 g/L, 7 mM) and by Burns et al. (1.41 mg/mL, 15.7 mM) [29,31]. Different outputs were found in literature, probably due to differences related to the microorganisms involved, to the use of different substrates involved in the determination of the MIC values (broths at different pH) and to the temperatures applied in the tests.
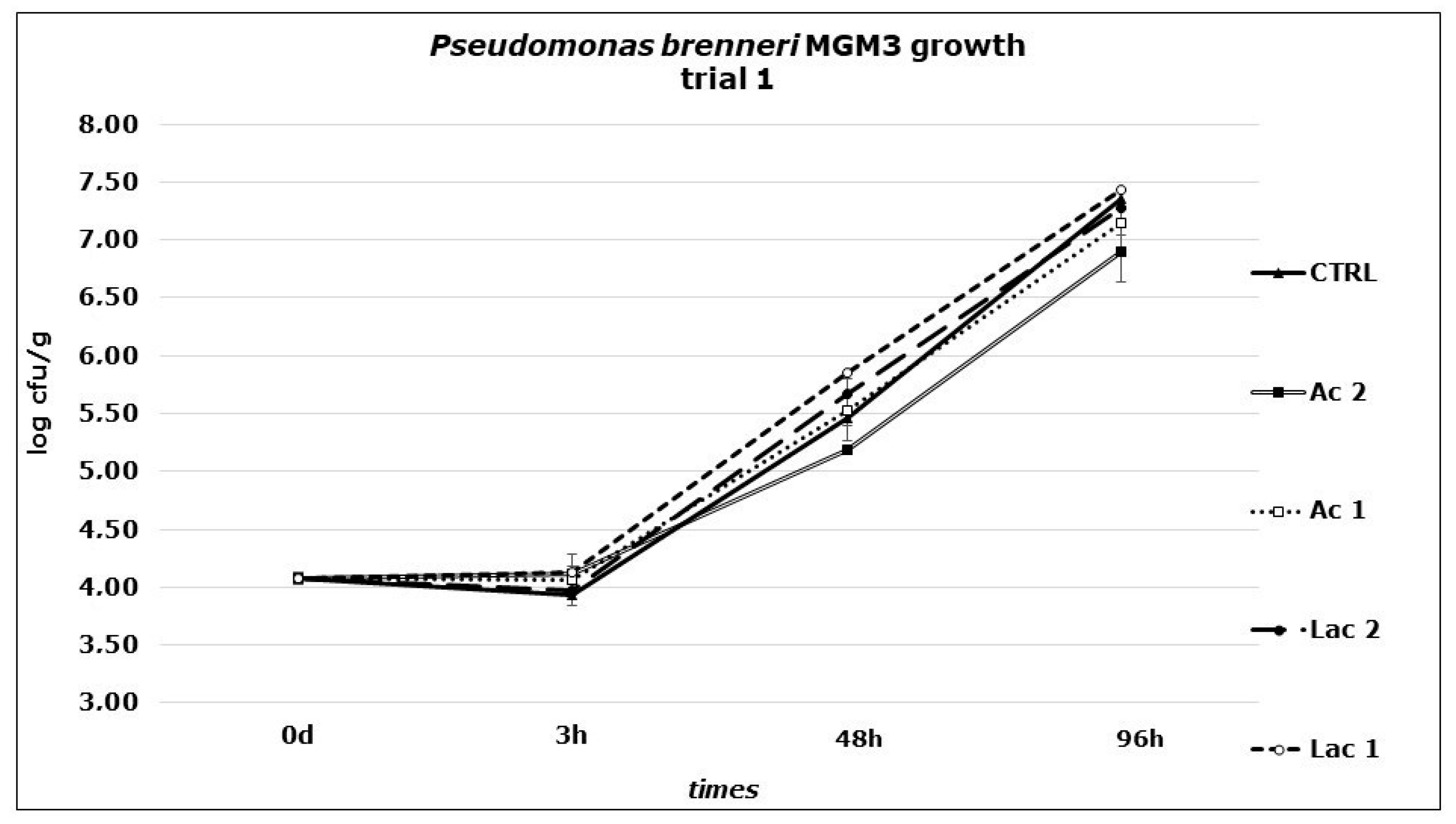
In the second stage of the study, experiments were carried out to identify the effect of the addition of acetic or lactic acid against P. brenneri MGM3 on Primo sale cheese: during the first trial (Figure 1), no significant differences were revealed in bacterial growth between control samples and treated ones, when applying surface dipping with acetic or lactic acid solutions with a concentration equal to the MIC determined in the first part of the study or doubling this concentration. No strain inactivation or growth reduction was observed, independently form the treatment (as shown by the values obtained at t3). A rapid growth at 6 °C was revealed in all the four series, with increases from t0 after 4 days of +3.28, +2.83, +3.08, +3.20 and +3.36 Log CFU/g in CTRL, Ac1, Ac2, Lac1 and Lac2, respectively. pH showed a slight decrease from t0 to the end of the trial (t96) from on average 6.31 to 6.02 (CTRL), 6.02 (Ac1), 6.08 (Ac2), 6.15 (Lac1) and 6.06 (Lac2), respectively. Such similarity among all the solutions tested, including those with a concentration corresponding to the double of the MIC obtained in broth, demonstrated the importance of the buffer activity of the food matrix. Moreover, the environmental conditions of a real cheese substrate can give some protection to the bacteria present on the surface, supplying numerous sites for attaching and growing; specific surface microenvironments can act as niches where the microorganisms can survive to harsh conditions (salt concentration, low temperature, low pH) and grow. Thus, these concentrations failed to prevent the growth of P. brenneri. in a real substrate condition.
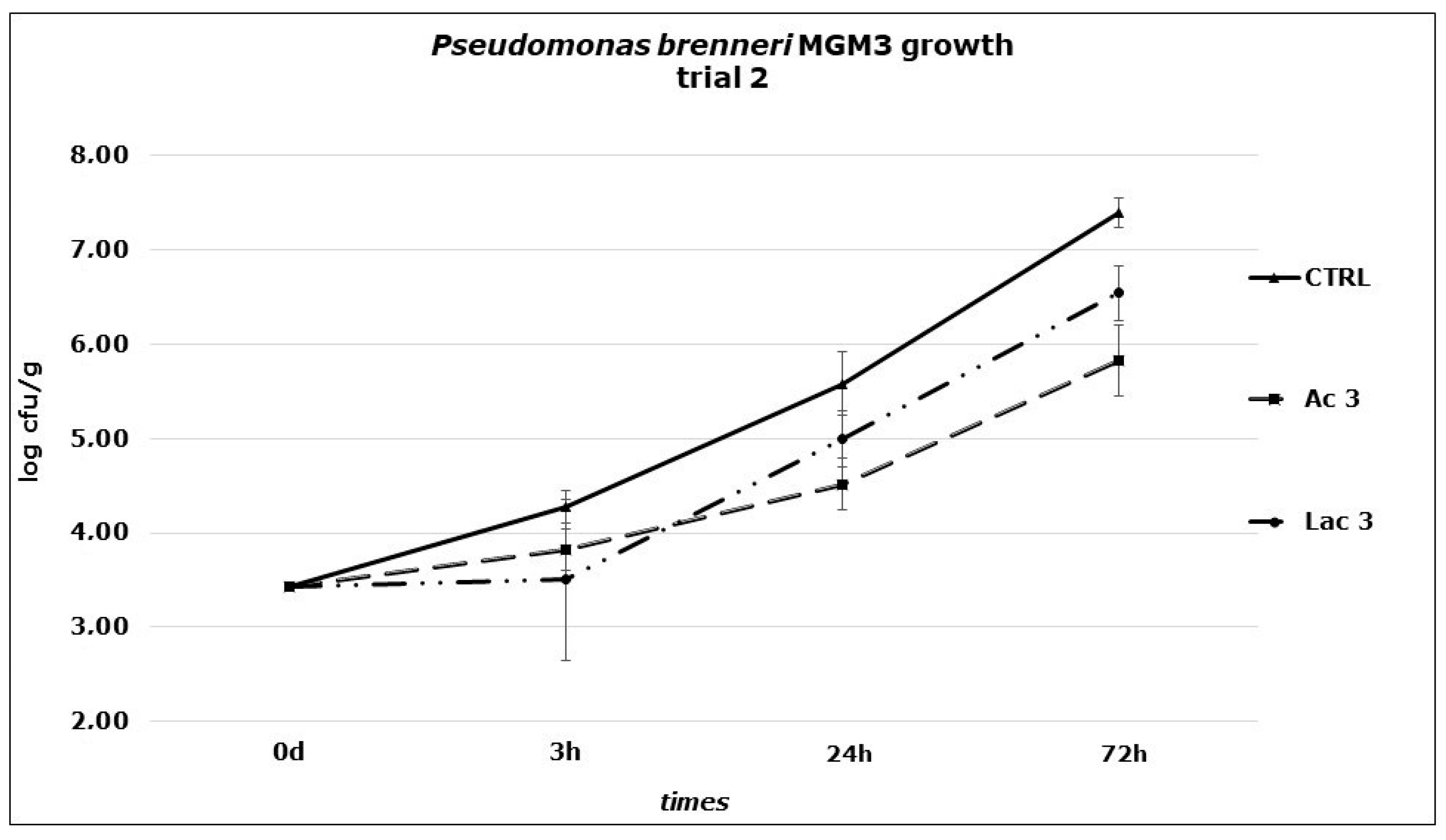
In the second trial, the effect of the two organic acids against P. brenneri MGM3 growth on Primo sale cheese at 6 °C at higher concentrations were investigated (three times the MICs obtained in broth). The results obtained (Figure 2) showed a significantly lower growth in Ac3 (149.92 mM) samples if compared to control ones (p < 0.01): starting from an initial concentration of 3 Log CFU/g, a faster increase was observed in CTRL samples, reaching a final load of 7.39 Log CFU/g while a delayed and slower growth in Ac3 series led to a final load not overcoming 6 Log CFU/g. At the same conditions, Lac3 (133.20 mM) samples showed a significantly lower growth if compared to CTRL samples (p < 0.05), reaching a final value of 6.54 Log CFU/g. Despite the lower value obtained with the addition of acetic acid if compared to lactic acid, the comparison of the two treated series did not show any significant difference. In this trial, cheese pH were shown to be affected by the addition of organic acid solutions: a slight and gradual decrease from t0 (6.30) to t72 (6.18, CTRL), (6.01, Ac3), (5.12 Lac3), was reached.
A conditioning of the substrate by dipping the surface of cheese in a solution of organic acids after production could be a useful hurdle to reduce the increase of P. brenneri. The growth of this spoilage agent was not completely avoided thus no inactivation was observed, but it was slowed down significantly, especially by acetic acid, obtaining a difference of 1.57 Log CFU/g after three days of storage in Ac3 series. This outcome could be exploited in the dairy industry to delay the spoilage by Pseudomonas: it could be advantageous in terms of shelf-life extension, taking in mind the threshold associated with lower product acceptability (6 Log CFU/g) and the value that corresponds to negative sensorial properties of the product (7 Log CFU/g) [32,33]. A reduction in Pseudomonas growth could allow to reach later these two limits.
When defining an application protocol, the producer should balance the needs to reach growth inhibition (and a consequently longer shelf life) with the minimization of the effect on the product taste, thus maintaining the original characteristics of this popular cheese. In this light, further research is needed, including the parallel evaluation of different combinations with concentration scales of lactic acid and acetic acid. This approach could allow to define the relative impact of each acid on both sensorial characters (evaluated by panellists) and distance from the microbial growth boundary.
4. Conclusions
The surface of fresh cheeses is characterized by permissive conditions for growth of spoilage microorganisms. Moreover, during production, contamination cannot be completely avoided; in this light, Pseudomonas spp. represents the main impediment to extend fresh cheeses’ shelf life. This study shows the potential inhibitory activity of acetic or lactic dipping as final surface treatment for Primo sale cheese; nevertheless, such treatment is intended to be applied as a part of an integrated hygiene plan, in order to couple a low initial microbial load with a scarce growth through the shelf life.
Author Contributions
The individual contribution of the authors of this paper is indicated below: E.T.: conception and design of the work; microbiological investigation; draft preparation. C.B.: microbiological investigation; chemical solutions; revision of the paper. S.S.: conception and design of the work; microbiological investigation; revision of the paper. All authors have read and agreed to the published version of the manuscript.
Funding
No funding were received.
Acknowledgments
We thank Maria Filippa Addis and Martina Penati of the Microbiology Laboratory of the Department of Veterinary Medicine University of Milan for providing the MALDI-TOF strain identification.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gustavsson, J.; Cederberg, C.; Sonesson, U.; Van Otterdijk, R.; Meybeck, A. Global Food Losses and Food Waste; Food and Agriculture Organization of the United Nations: Rome, Italy, 2011. [Google Scholar]
- Sperber, W.H.; Doyle, M.P. Compendium of the Microbiological Spoilage of Foods and Beverages; Springer: New York, NY, USA, 2009. [Google Scholar]
- Petruzzi, L.; Corbo, M.R.; Sinigaglia, M.; Bevilacqua, A. Chapter 1—Microbial spoilage of foods: Fundamentals. In The Micro-Biological Quality of Food—Foodborne Spoilers; Bevilacqua, A., Corbo, M.R., Sinigaglia, M., Eds.; Woodhead Publishing: Sawston, Cambridge, UK, 2017; pp. 1–21. [Google Scholar]
- Kraft, A.A. Psychrotrophic Bacteria in Foods: Disease and Spoilage; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]
- Wiedmann, M.; Weilmeier, D.; Dineen, S.S.; Ralyea, R.; Boor, K.J. Molecular and Phenotypic Characterization ofPseudomonas spp. Isolated from Milk. Appl. Environ. Microbiol. 2000, 66, 2085–2095. [Google Scholar] [CrossRef] [Green Version]
- Spanu, C.; Piras, F.; Mocci, A.; Nieddu, G.; De Santis, E.; Scarano, C. Use of Carnobacterium spp protective culture in MAP packed Ricotta fresca cheese to control Pseudomonas spp. Food Microbiol. 2018, 74, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Cantoni, C.; Stella, S.; Ripamonti, B.; Marchese, R. Pseudomonas fluorescens and anomalous colourations of mozzarella cheese. Ind. Aliment. 2001, 40, 33–35. [Google Scholar]
- Cantoni, C.; Stella, S.; Cozzi, M.; Iacumin, L.; Comi, G. Blue colouring in mozzarella cheese. Ind. Aliment. 2003, 42, 840–843. [Google Scholar]
- Carminati, D.; Bonvini, B.; Rossetti, L.; Zago, M.; Tidona, F.; Giraffa, G. Investigation on the presence of blue pigment-producing Pseudomonas strains along a production line of fresh mozzarella cheese. Food Control 2019, 100, 321–328. [Google Scholar] [CrossRef]
- Dogan, B.; Boor, K.J. Genetic Diversity and Spoilage Potentials among Pseudomonas spp. Isolated from Fluid Milk Products and Dairy Processing Plants. Appl. Environ. Microbiol. 2003, 69, 130–138. [Google Scholar] [CrossRef] [Green Version]
- Carrascosa, C.; Millán, R.; Jaber, J.R.; Lupiola, P.; del Rosario-Quintana, C.; Mauricio, C.; Sanjuán, E. Blue pigment in fresh cheese produced by Pseudomonas fluorescens. Food Control 2015, 54, 95–102. [Google Scholar] [CrossRef]
- Scatamburlo, T.; Yamazi, A.; Cavicchioli, V.; Pieri, F.; Nero, L.A.; Moreira, T. Spoilage potential of Pseudomonas species isolated from goat milk. J. Dairy Sci. 2015, 98, 759–764. [Google Scholar] [CrossRef]
- Rossi, C.; Serio, A.; Chaves-López, C.; Anniballi, F.; Auricchio, B.; Goffredo, E.; Cenci-Goga, B.T.; Lista, F.; Fillo, S.; Paparella, A. Biofilm formation, pigment production and motility in Pseudomonas spp. isolated from the dairy industry. Food Control 2018, 86, 241–248. [Google Scholar] [CrossRef]
- Arslan, S.; Eyi, A.; Özdemir, F. Spoilage potentials and antimicrobial resistance of Pseudomonas spp. isolated from cheeses. J. Dairy Sci. 2011, 94, 5851–5856. [Google Scholar] [CrossRef] [Green Version]
- Van Tassell, J.; Martin, N.; Murphy, S.; Wiedmann, M.; Boor, K.; Ivy, R. Evaluation of various selective media for the detection of Pseudomonas species in pasteurized milk. J. Dairy Sci. 2012, 95, 1568–1574. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, P.J. Coupling of Phosphorylation to Electron and Hydrogen Transfer by a Chemi-Osmotic type of Mechanism. Nat. Cell Biol. 1961, 191, 144–148. [Google Scholar] [CrossRef]
- European Union. Regulation (EC) No 1333/2008. In Proceedings of the European Parliament and of the Council of 16 December 2008 on Food Additives, Bruxelles, Belgium, 31 December 2008; Volume L 354, pp. 16–33. [Google Scholar]
- Tirloni, E.; Nauta, M.; Vasconi, M.; Di Pietro, V.; Bernardi, C.; Stella, S. Growth of Listeria monocytogenes in ready-to-eat “shrimp cocktail”: Risk assessment and possible preventive interventions. Int. J. Food Microbiol. 2020, 334, 108800. [Google Scholar] [CrossRef] [PubMed]
- Baïda, N.; Yazourh, A.; Singer, E.; Izard, D. Pseudomonas brenneri sp. nov., a new species isolated from natural mineral waters. Res. Microbiol. 2001, 152, 493–502. [Google Scholar] [CrossRef]
- Andreani, N.; Martino, M.; Fasolato, L.; Carraro, L.; Montemurro, F.; Mioni, R.; Bordin, P.; Cardazzo, B. Reprint of ‘Tracking the blue: A MLST approach to characterise the Pseudomonas fluorescens group’. Food Microbiol. 2015, 45, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.R.; Moss, M.O. Food Microbiology, 2nd ed.; Royal Society of Chemistry: London, UK, 2000. [Google Scholar]
- Gunasekera, T.; Dorsch, M.; Slade, M.; Veal, D. Specific detection of Pseudomonas spp. in milk by fluorescence in situ hybridi-zation using ribosomal RNA directed probes. J. Appl. Microbiol. 2003, 94, 936–945. [Google Scholar] [CrossRef] [Green Version]
- Baruzzi, F.; Lagonigro, R.; Quintieri, L.; Morea, M.; Caputo, L. Occurrence of non-lactic acid bacteria populations involved in protein hydrolysisof cold-stored high moisture Mozzarella cheese. Food Microbiol. 2012, 30, 37–44. [Google Scholar] [CrossRef]
- Nogarol, C.; Acutis, P.L.; Bianchi, D.M.; Maurella, C.; Peletto, S.; Gallina, S.; Adriano, D.; Zuccon, F.; Borrello, S.; Caramelli, M.; et al. Molecular Characterization of Pseudomonas fluorescens Isolates Involved in the Italian “Blue Mozzarella” Event. J. Food Prot. 2013, 76, 500–504. [Google Scholar] [CrossRef]
- Del Olmo, A.; Calzada, J.; Nuñez, M. The blue discoloration of fresh cheeses: A worldwide defect associated to specific contamination by Pseudomonas fluorescens. Food Control 2018, 86, 359–366. [Google Scholar] [CrossRef]
- Sinigaglia, M.; Bevilacqua, A.; Corbo, M.R.; Pati, S.; Del Nobile, M.A. Use of active compounds for prolonging the shelf life of mozzarella cheese. Int. Dairy J. 2008, 18, 624–630. [Google Scholar] [CrossRef]
- Cruz-Romero, M.; Murphy, T.; Morris, M.; Cummins, E.; Kerry, J. Antimicrobial activity of chitosan, organic acids and nano-sized solubilisates for potential use in smart antimicrobially-active packaging for potential food applications. Food Control 2013, 34, 393–397. [Google Scholar] [CrossRef]
- Moreira, T.S.M.; de Oliveira, A.; Barlati, T.; da Silva, V.; Rodrigues Dos Santos, A.; Gonçalves, O.H.; da Silva Gonzalez, O.; Droval, A.A.; Leimann, F.V. Hydrogels based on gelatin: Effect of lactic and acetic acids on microstructural modifications, water absorption mechanisms and antibacterial activity. LWT 2019, 103, 69–77. [Google Scholar] [CrossRef]
- Nakai, S.A.; Siebert, K.J. Organic acid inhibition models for Listeria innocua, Listeria ivanovii, Pseudomonas aeruginosa and Oenococcus oeni. Food Microbiol. 2004, 21, 67–72. [Google Scholar] [CrossRef]
- Bushell, F.M.L.; Tonner, P.D.; Jabbari, S.; Schmid, A.K.; Lund, P.A. Synergistic Impacts of Organic Acids and pH on Growth of Pseudomonas aeruginosa: A Comparison of Parametric and Bayesian Non-parametric Methods to Model Growth. Front. Microbiol. 2019, 9, 3196. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.; McCoy, C.P.; Irwin, N.J. Synergistic activity of weak organic acids against uropathogens. J. Hosp. Infect. 2021. [Google Scholar] [CrossRef]
- Leriche, F.; Bordessoules, A.; Fayolle, K.; Karoui, R.; Laval, K.; Leblanc, L.; Dufour, E. Alteration of raw-milk cheese by Pseudomonas spp.: Monitoring the sources of contamination using fluorescence spectroscopy and metabolic profiling. J. Microbiol. Methods 2004, 59, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Lucera, A.; Mastromatteo, M.; Conte, A.; Zambrini, A.V.; Faccia, M.; Del Nobile, M.A. Effect of active coating on microbiological and sensory properties of fresh mozzarella cheese. Food Packag. Shelf 2014, 1, 25–29. [Google Scholar] [CrossRef]
Figure 1.
P. brenneri MGM3 counts obtained in Primo sale cheese during the first challenge test performed at 6 °C. Dipping: Ac1: acetic acid, 49.96 mM; Ac2: acetic acid, 99.92 mM; Lac1: lactic acid, 44.40 mM; Lac2: lactic acid, 88.80 mM; CTRL: distilled water.
Figure 1.
P. brenneri MGM3 counts obtained in Primo sale cheese during the first challenge test performed at 6 °C. Dipping: Ac1: acetic acid, 49.96 mM; Ac2: acetic acid, 99.92 mM; Lac1: lactic acid, 44.40 mM; Lac2: lactic acid, 88.80 mM; CTRL: distilled water.

Figure 2.
P. brenneri MGM3 counts obtained in Primo sale cheese during the second challenge test performed at 6 °C. Dipping: Ac3: acetic acid, 149.89 mM; Lac3: lactic acid, 133.20 mM; CTRL: distilled water.
Figure 2.
P. brenneri MGM3 counts obtained in Primo sale cheese during the second challenge test performed at 6 °C. Dipping: Ac3: acetic acid, 149.89 mM; Lac3: lactic acid, 133.20 mM; CTRL: distilled water.


{kind=link}
{kind=link}
Table 1.
Optical Density of P. brenneri MGM3 suspensions in Brain Heart Infusion (BHI) broth at 30 °C in presence of different concentrations of lactic and acetic acid (the value indicates the difference towards the respective blank sample). MIC detected are written in bold.
Table 1.
Optical Density of P. brenneri MGM3 suspensions in Brain Heart Infusion (BHI) broth at 30 °C in presence of different concentrations of lactic and acetic acid (the value indicates the difference towards the respective blank sample). MIC detected are written in bold.
| Acetic Acid | t0 | t24 | t48 | t72 |
|---|---|---|---|---|
| Average ± Std.Dev | Average ± Std.Dev | Average ± Std.Dev | Average ± Std.Dev | |
| 49.96 mM | −0.02 ± 0.03 | 0.01 ± 0.04 | 0.01 ± 0.04 | 0.01 ± 0.04 |
| 24.98 mM | −0.05 ± 0.01 | −0.03 ± 0.42 | 0.28 ± 0.42 | 0.57 ± 0.16 |
| 12.49 mM | −0.04 ± 0.01 | 0.12 ± 0.01 | 1.54 ± 0.01 | 1.76 ± 0.13 |
| 5.00 mM | −0.12 ± 0.01 | 0.20 ± 0.18 | 1.55 ± 0.18 | 1.55 ± 0.14 |
| 2.50 mM | −0.09 ± 0.01 | 0.28 ± 0.02 | 1.47 ± 0.02 | 1.61 ± 0.03 |
| CTRL | 0.01 ± 0.09 | 0.35 ± 0.01 | 1.65 ± 0.01 | 1.72 ± 0.06 |
| Lactic Acid | t0 | t24 | t48 | t72 |
| Average ± std.dev | Average ± std.dev | Average ± std.dev | Average ± std.dev | |
| 44.40 mM | −0.06 ± 0.03 | −0.04 ± 0.02 | −0.05 ± 0.02 | 0.01 ± 0.01 |
| 22.20 mM) | −0.08 ± 0.01 | −0.03 ± 0.04 | 1.42 ± 0.01 | 1.79 ± 0.06 |
| 11.10 mM | −0.08 ± 0.05 | 0.24 ± 0.03 | 1.60 ± 0.04 | 1.65 ± 0.04 |
| 5.55 mM | −0.22 ± 0.26 | 0.35 ± 0.01 | 1.58 ± 0.15 | 1.56 ± 0.12 |
| 2.78 mM | 0.24 ± 0.39 | 0.35 ± 0.07 | 1.57 ± 0.04 | 1.80 ± 0.09 |
| CTRL | 0.01 ± 0.09 | 0.35 ± 0.05 | 1.65 ± 0.01 | 1.72 ± 0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tirloni, E.; Bernardi, C.; Stella, S. Pseudomonas spp.: Are Food Grade Organic Acids Efficient against These Spoilage Microorganisms in Fresh Cheeses? Foods 2021, 10, 891. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10040891
AMA Style
Tirloni E, Bernardi C, Stella S. Pseudomonas spp.: Are Food Grade Organic Acids Efficient against These Spoilage Microorganisms in Fresh Cheeses? Foods. 2021; 10(4):891. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10040891
Chicago/Turabian StyleTirloni, Erica, Cristian Bernardi, and Simone Stella. 2021. "Pseudomonas spp.: Are Food Grade Organic Acids Efficient against These Spoilage Microorganisms in Fresh Cheeses?" Foods 10, no. 4: 891. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10040891
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.