
Integrated Phenotypic–Genotypic Analysis of Latilactobacillus sakei from Different Niches

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Strains, Genome Sequencing, and Data Assembly
2.2. Genome Features Prediction and Annotation
2.3. Pan-Genome and Core-Genome Analysis
2.4. Average Nucleotide Identity (ANI) Values and Phylogenetic Analyses
2.5. Genotype/Phenotype Association Applied to Carbohydrate Metabolism
2.6. Genotype/Phenotype Association Applied to Antibiotic Resistance
2.7. CRISPR Identification and Characterization of Isolated Strains
3. Results
3.1. General Genome Characteristics of Latilactobacillus sakei
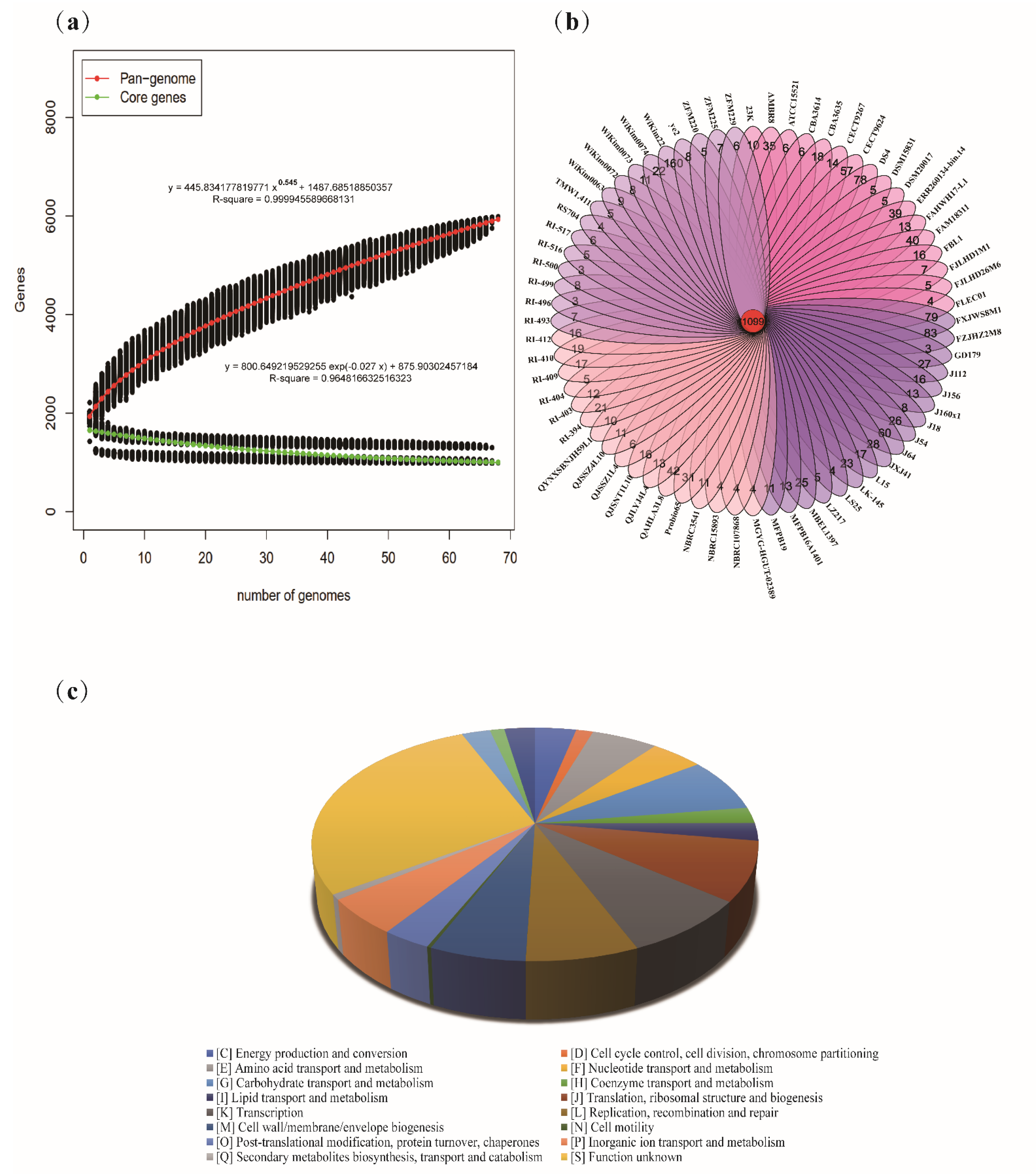
3.2. Pan-Genome and Core Genes of Latilactobacillus sakei
3.3. ANI and Phylogenetic Analyses of Latilactobacillus sakei
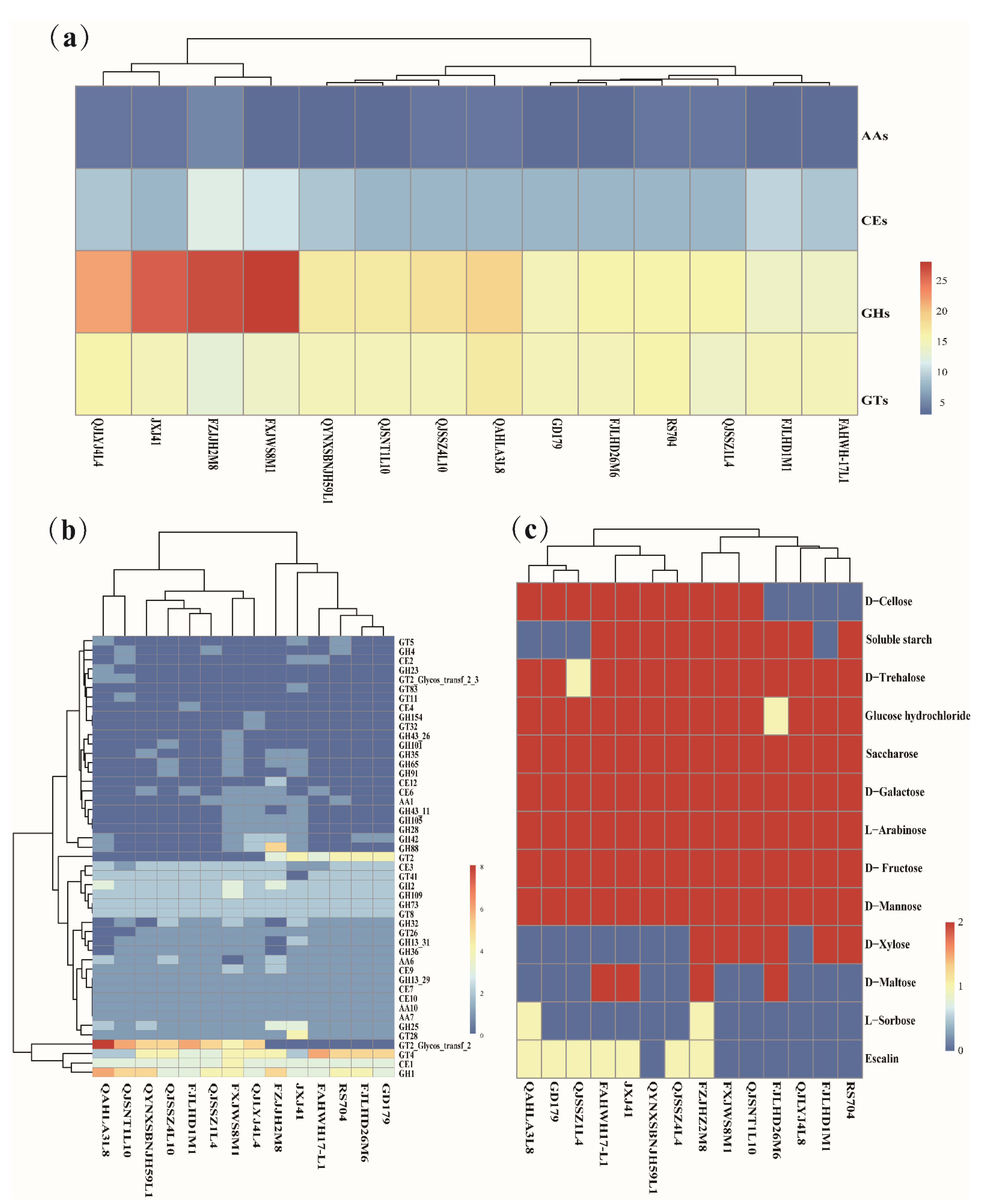
3.4. Genotype/Phenotype Association Analysis for Carbohydrates Utilization in Latilactobacillus sakei
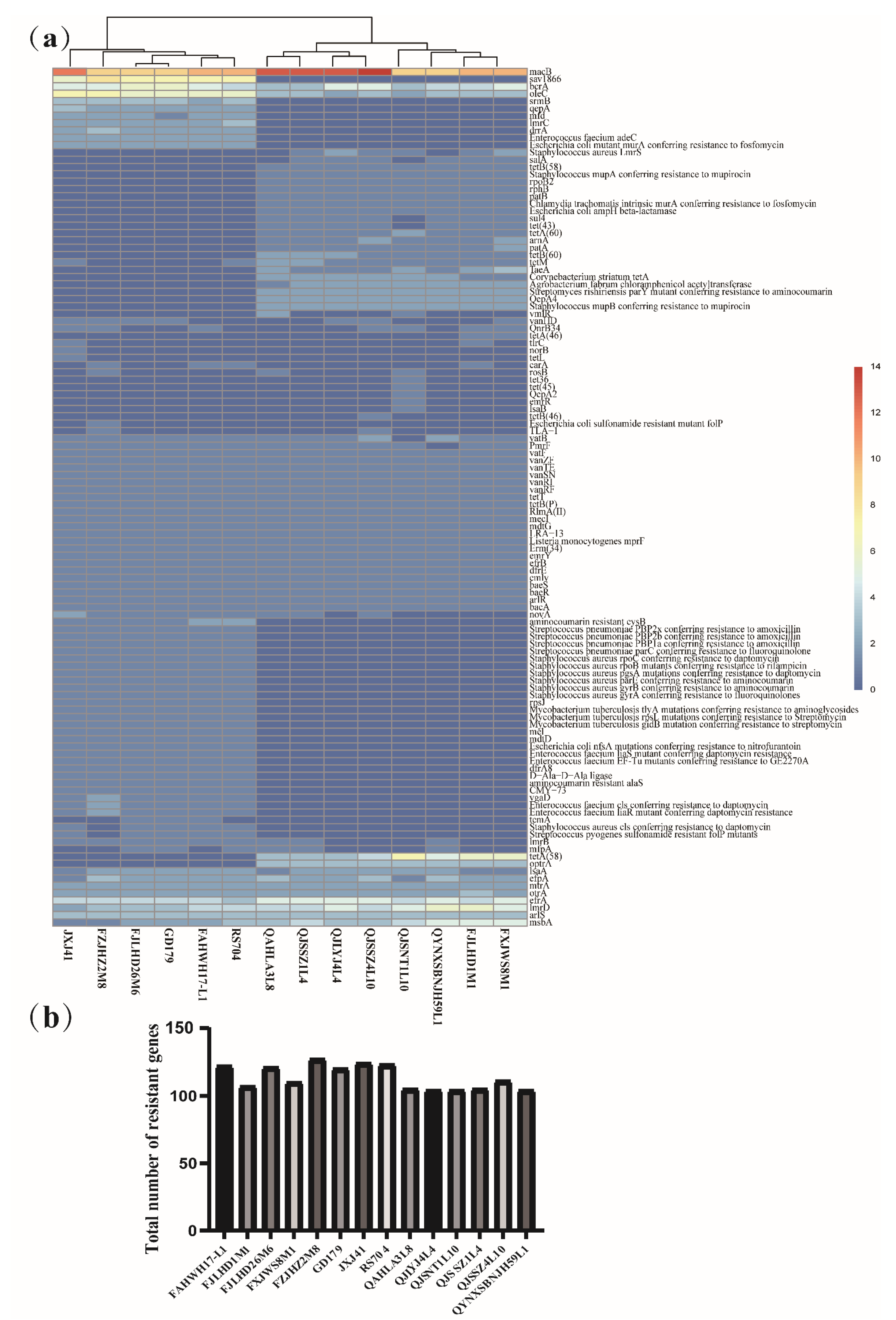
3.5. Genotype/Phenotype Association Analysis for Antibiotic Resistance in Latilactobacillus sakei
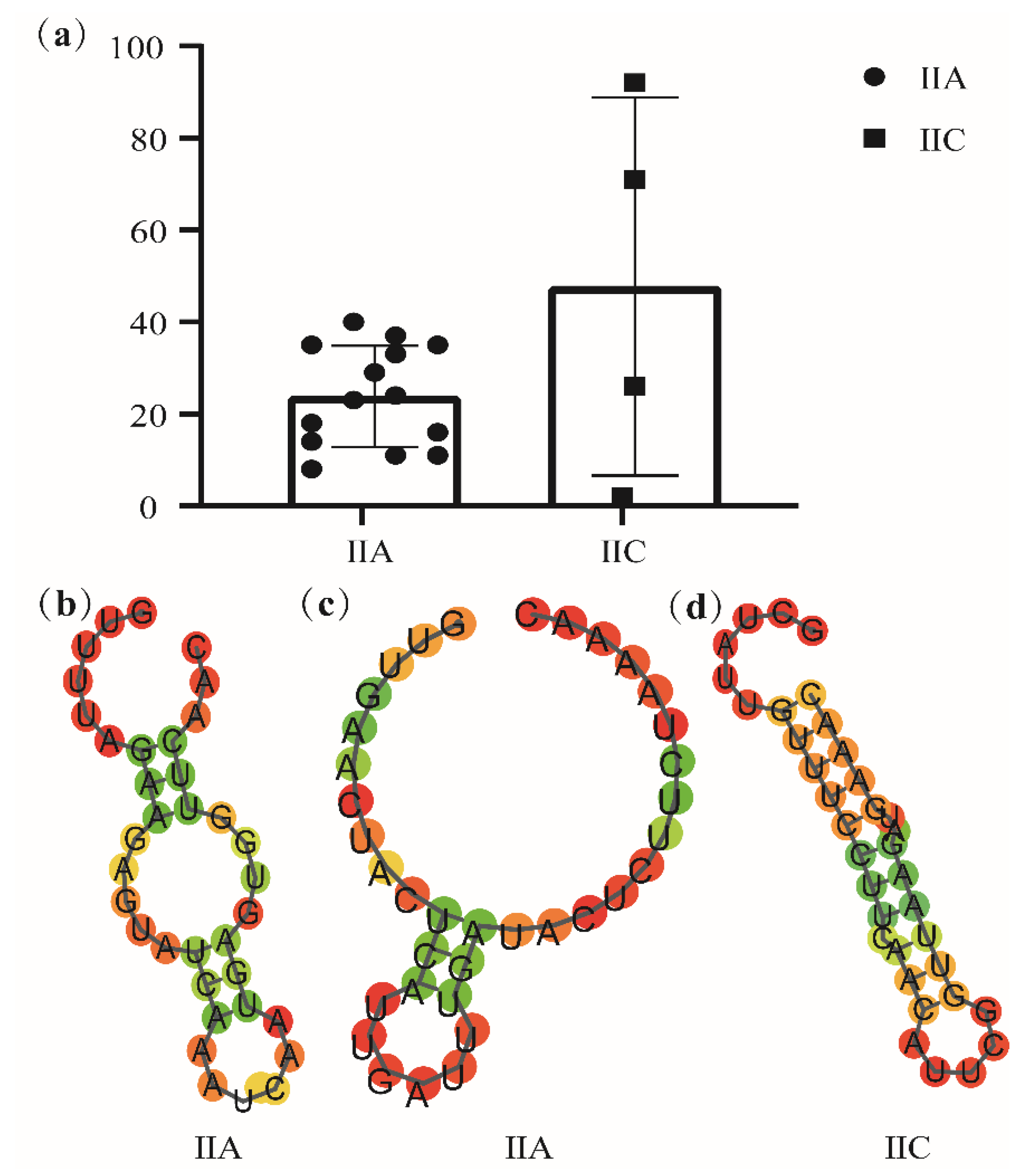
3.6. Prediction of CRISPR–Cas Systems in Latilactobacillus sakei
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Katagiri, H.; Kitahara, K.; Fukami, K.; Sugase, M. The Characteristics of the Lactic Acid Bacteria Isolated from Moto, Yeast Mashes for Saké Manufacture. Bull. Agric. Chem. Soc. Jpn. 1934, 10, 153–157. [Google Scholar] [CrossRef]
- Moroni, A.V.; Arendt, E.K.; Bello, F.D. Biodiversity of lactic acid bacteria and yeasts in spontaneously-fermented buckwheat and teff sourdoughs. Food Microbiol. 2011, 28, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Reale, A.; Di Renzo, T.; Boscaino, F.; Nazzaro, F.; Fratianni, F.; Aponte, M. Lactic Acid Bacteria Biota and Aroma Profile of Italian Traditional Sourdoughs from the Irpinian Area in Italy. Front. Microbiol. 2019, 10, 1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashaolu, T.J.; Reale, A. A Holistic Review on Euro-Asian Lactic Acid Bacteria Fermented Cereals and Vegetables. Microorganisms 2020, 8, 1176. [Google Scholar] [CrossRef]
- Chaillou, S.; Chaulot-Talmon, A.; Caekebeke, H.; Cardinal, M.; Christieans, S.; Denis, C.; Desmonts, M.H.; Dousset, X.; Feurer, C.; Hamon, E.; et al. Origin and ecological selection of core and food-specific bacterial communities associated with meat and seafood spoilage. ISME J. 2015, 9, 1105–1118. [Google Scholar] [CrossRef] [Green Version]
- Bello, F.D.; Walter, J.; Hammes, W.; Hertel, C. Increased Complexity of the Species Composition of Lactic Acid Bacteria in Human Feces Revealed by Alternative Incubation Condition. Microb. Ecol. 2003, 45, 455–463. [Google Scholar] [CrossRef]
- Carvalho, K.G.; Bambirra, F.H.S.; Nicoli, J.R.; Oliveira, J.S.; Santos, A.M.C.; Bemquerer, M.; Miranda, A.; Franco, B.D.G.M. Characterization of multiple antilisterial peptides produced by sakacin P-producing Lactobacillus sakei subsp. sakei 2a. Arch. Microbiol. 2018, 200, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Mathiesen, G.; Huehne, K.; Kroeckel, L.; Axelsson, L.; Eijsink, V.G.H. Characterization of a New Bacteriocin Operon in Sakacin P-Producing Lactobacillus sakei, Showing Strong Translational Coupling between the Bacteriocin and Immunity Genes. Appl. Environ. Microbiol. 2005, 71, 3565–3574. [Google Scholar] [CrossRef] [Green Version]
- Winkelstroter, L.; Gomes, B.C.; Thomaz, M.R.; Souza, V.M.; De Martinis, E.C. Lactobacillus sakei 1 and its bacteriocin influence adhesion of Listeria monocytogenes on stainless steel surface. Food Control. 2011, 22, 1404–1407. [Google Scholar] [CrossRef]
- Gao, Y.; Jia, S.; Gao, Q.; Tan, Z. A novel bacteriocin with a broad inhibitory spectrum produced by Lactobacillus sake C2, isolated from traditional Chinese fermented cabbage. Food Control. 2010, 21, 76–81. [Google Scholar] [CrossRef]
- Lopez-Arvizu, A.; Rocha-Mendoza, D.; Ponce-Alquicira, E.; García-Cano, I. Characterization of antibacterial activity of a N-acetylmuramoyl-l-alanine amidase produced by Latilactobacillus sakei isolated from salami. World J. Microbiol. Biotechnol. 2021, 37, 1–13. [Google Scholar] [CrossRef]
- Gao, Y.; Li, D.; Liu, X. Effects of Lactobacillus sakei C2 and sakacin C2 individually or in combination on the growth of Listeria monocytogenes, chemical and odor changes of vacuum-packed sliced cooked ham. Food Control. 2015, 47, 27–31. [Google Scholar] [CrossRef]
- Najjari, A.; Boumaiza, M.; Jaballah, S.; Boudabous, A.; Ouzari, H. Application of isolated Lactobacillus sakei and Staphylococcus xylosus strains as a probiotic starter culture during the industrial manufacture of Tunisian dry-fermented sausages. Food Sci. Nutr. 2020, 8, 4172–4184. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.H.; Moon, J.S.; Chang, H.C. Rapid manufacture and quality evaluation of long-term fermented kimchi (mukeunji) using Lactobacillus sakei SC1. Food Sci. Biotechnol. 2015, 24, 1797–1804. [Google Scholar] [CrossRef]
- Hong, Y.-F.; Kim, H.; Kim, H.R.; Gim, M.G.; Chung, D.K. Different immune regulatory potential of Lactobacillus plantarum and Lactobacillus sakei isolated from Kimchi. J. Microbiol. Biotechnol. 2014, 24, 1629–1635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Won, S.-M.; Chen, S.; Lee, S.Y.; Lee, K.E.; Park, K.W.; Yoon, J.-H. Lactobacillus sakei ADM14 Induces Anti-Obesity Effects and Changes in Gut Microbiome in High-Fat Diet-Induced Obese Mice. Nutrients 2020, 12, 3703. [Google Scholar] [CrossRef]
- Rather, I.A.; Bajpai, V.K.; Ching, L.L.; Majumder, R.; Nam, G.-J.; Indugu, N.; Singh, P.; Kumar, S.; Hajrah, N.H.; Sabir, J.S.; et al. Effect of a bioactive product SEL001 from Lactobacillus sakei probio65 on gut microbiota and its anti-colitis effects in a TNBS-induced colitis mouse model. Saudi J. Biol. Sci. 2020, 27, 261–270. [Google Scholar] [CrossRef]
- Rather, I.A.; Bajpai, V.K.; Huh, Y.S.; Han, Y.-K.; Bhat, E.A.; Lim, J.; Paek, W.K.; Park, Y.-H. Probiotic Lactobacillus sakei proBio-65 Extract Ameliorates the Severity of Imiquimod Induced Psoriasis-Like Skin Inflammation in a Mouse Model. Front. Microbiol. 2018, 9, 1021. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.-Y.; Shin, J.-S.; Lee, S.-G.; Rhee, Y.K.; Cho, C.-W.; Hong, H.-D.; Lee, K.-T. Lactobacillus sakei K040706 evokes immunostimulatory effects on macrophages through TLR 2-mediated activation. Int. Immunopharmacol. 2015, 28, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Chaillou, S.; Champomier-Vergès, M.-C.; Cornet, M.; Coq, A.-M.C.-L.; Dudez, A.-M.; Martin, V.; Beaufils, S.; Darbon-Rongère, E.; Bossy, R.; Loux, V.; et al. The complete genome sequence of the meat-borne lactic acid bacterium Lactobacillus sakei 23K. Nat. Biotechnol. 2005, 23, 1527–1533. [Google Scholar] [CrossRef] [Green Version]
- Chaillou, S.; Lucquin, I.; Najjari, A.; Zagorec, M.; Champomier-Vergès, M.-C. Population Genetics of Lactobacillus sakei Reveals Three Lineages with Distinct Evolutionary Histories. PLoS ONE 2013, 8, e73253. [Google Scholar] [CrossRef]
- Nyquist, O.L.; McLeod, A.; Brede, D.A.; Snipen, L.; Aakra, Å.; Nes, I.F. Comparative genomics of Lactobacillus sakei with emphasis on strains from meat. Mol. Genet. Genom. 2011, 285, 297–311. [Google Scholar] [CrossRef]
- Donnell, M.O.; Harris, H.; Jeffery, I.; Claesson, M.; Younge, B.; Toole, P.O.; Ross, R. The core faecal bacterial microbiome of Irish Thoroughbred racehorses. Lett. Appl. Microbiol. 2013, 57, 492–501. [Google Scholar] [CrossRef]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y.; et al. SOAPdenovo2: An empirically improved memory-efficient short-read de novo assembler. GigaScience 2012, 1, 18. [Google Scholar] [CrossRef]
- Loux, V.; Coeuret, G.; Zagorec, M.; Vergès, M.-C.C.; Chaillou, S. Complete and Draft Genome Sequences of Nine Lactobacillus sakei Strains Selected from the Three Known Phylogenetic Lineages and Their Main Clonal Complexes. Genome Announc. 2018, 6, e00082-18. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.H.; Chun, B.H.; Baek, J.H.; Roh, S.W.; Lee, S.H.; Jeon, C.O. Genomic and metabolic features of Lactobacillus sakei as revealed by its pan-genome and the metatranscriptome of kimchi fermentation. Food Microbiol. 2020, 86, 103341. [Google Scholar] [CrossRef] [PubMed]
- Inglin, R.C.; Meile, L.; Stevens, M.J.A. Draft Genome Sequences of 43 Lactobacillus Strains from the Species L. curvatus, L. fermentum, L. paracasei, L. plantarum, L. rhamnosus, and L. sakei, Isolated from Food Product. Genome Announc. 2017, 5, e00632-17. [Google Scholar] [CrossRef] [Green Version]
- Jans, C.; Lagler, S.; Lacroix, C.; Meile, L.; Stevens, M.J.A. Complete Genome Sequences of Lactobacillus curvatus KG6, L. curvatus MRS6, and Lactobacillus sakei FAM18311, Isolated from Fermented Meat Products. Genome Announc. 2017, 5, e00915-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, H.I.; Lee, J.; Jang, J.Y.; Park, H.W.; Choi, H.J.; Kim, T.W.; Kang, M.R.; Lee, J.H. Draft Genome Sequence of Lactobacillus sakei Strain wikim 22, Isolated from Kimchi in Chungcheong Province, South Korea. Genome Announc. 2014, 2, e01184-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilıkkan, Ö.K. CRISPR/Cas systems and anti-repeat sequences of Lactobacillus curvatus, Lactobacillus graminis, Lactobacillus fuchuensis, and Lactobacillus sakei genomes. Korean J. Microbiol. 2021, 57, 12–22. [Google Scholar] [CrossRef]
- Kwun, S.-Y.; Yoon, J.-A.; Park, E.-H.; Kim, M.-D. Complete genome sequence data of Lactobacillus sakei MBEL1397 isolated from kimchi. Data Brief 2020, 31, 105740. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; Kim, E.; Kim, C.-G.; Choo, D.-W.; Kim, H.-Y. Draft Genome Sequence of Lactobacillus sakei Strain FBL1, a Probiotic Bacterium Isolated from Mukeunji, a Long-Fermented Kimchi, in South Korea. Genome Announc. 2016, 4, e00365-16. [Google Scholar] [CrossRef] [Green Version]
- Schuster, J.A.; Vogel, R.F.; Ehrmann, M.A. Characterization and distribution of CRISPR–Cas systems in Lactobacillus sakei. Arch. Microbiol. 2019, 201, 337–347. [Google Scholar] [CrossRef] [PubMed]
- McLeod, A.; Brede, D.A.; Rud, I.; Axelsson, L. Genome Sequence of Lactobacillus sakei subsp. sakei LS25, a Commercial Starter Culture Strain for Fermented Sausage. Genome Announc. 2013, 1, 1. [Google Scholar] [CrossRef] [Green Version]
- Kato, S.; Oikawa, T. Genome Sequence of Lactobacillus sakei LK-145 Isolated from a Japanese Sake Cellar as a High Producer of d-Amino Acids. Genome Announc. 2017, 5, e00656-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuno, K.; Mizuno, M.; Yamauchi, M.; Takemura, A.J.; Romero, V.M.; Morikawa, K. Adjacent-possible ecological niche: Growth of Lactobacillus species co-cultured with Escherichia coli in a synthetic minimal medium. Sci. Rep. 2017, 7, 12880. [Google Scholar] [CrossRef] [Green Version]
- Fujii, T.; Yamauchi, T.; Ishiyama, M.; Gogami, Y.; Oikawa, T.; Hata, Y. Crystallographic studies of aspartate racemase from Lactobacillus sakei NBRC 15893. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2015, 71, 1012–1016. [Google Scholar] [CrossRef] [Green Version]
- De Boeck, I.; Broek, M.F.V.D.; Allonsius, C.N.; Spacova, I.; Wittouck, S.; Martens, K.; Wuyts, S.; Cauwenberghs, E.; Jokicevic, K.; Vandenheuvel, D.; et al. Lactobacilli Have a Niche in the Human Nose. Cell Rep. 2020, 31, 107674. [Google Scholar] [CrossRef] [PubMed]
- Delcher, A.L.; Bratke, K.A.; Powers, E.C.; Salzberg, S. Identifying bacterial genes and endosymbiont DNA with Glimmer. Bioinformatics 2007, 23, 673–679. [Google Scholar] [CrossRef]
- Bairoch, A.; Apweiler, R. The SWISS-PROT protein sequence data bank and its supplement TrEMBL in 1999. Nucleic Acids Res. 1999, 27, 49–54. [Google Scholar] [CrossRef]
- Pruitt, K.D.; Tatusova, T.; Maglott, D.R. NCBI reference sequences (RefSeq): A curated non-redundant sequence database of genomes, transcripts and proteins. Nucleic Acids Res. 2007, 35, D61–D65. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Wu, J.; Yang, J.; Sun, S.; Xiao, J.; Yu, J. PGAP: Pan-genomes analysis pipeline. Bioinformatics 2012, 28, 416–418. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Stoeckert, C.J.; Roos, D.S. OrthoMCL: Identification of Ortholog Groups for Eukaryotic Genomes. Genome Res. 2003, 13, 2178–2189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valeriano, V.D.V.; Oh, J.K.; Bagon, B.B.; Kim, H.; Kang, D.-K. Comparative genomic analysis of Lactobacillus mucosae LM1 identifies potential niche-specific genes and pathways for gastrointestinal adaptation. Genomics 2019, 111, 24–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [Green Version]
- Kelleher, P.; Bottacini, F.; Mahony, J.; Kilcawley, K.N.; Van Sinderen, U. Comparative and functional genomics of the Lactococcus lactis taxon; insights into evolution and niche adaptation. BMC Genom. 2017, 18, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mailund, T.; Brodal, G.S.; Fagerberg, R.; Pedersen, C.N.S.; Phillips, D. Recrafting the neighbor-joining method. BMC Bioinform. 2006, 7, 29. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.-H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef]
- Lombard, V.; Ramulu, H.G.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [Green Version]
- Deng, W.; Wang, Y.; Liu, Z.; Cheng, H.; Xue, Y. HemI: A Toolkit for Illustrating Heatmaps. PLoS ONE 2014, 9, e111988. [Google Scholar] [CrossRef]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. CARD 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2017, 45, D566–D573. [Google Scholar] [CrossRef]
- Geneva, I.; Brussels, I.D.F. Milk and Milk Products. Determination of the Minimal Inhibitory Concentration (MIC) of Antibiotics Applicable to Bifidobacteria and Non-Enterococcal Lactic Acid Bacteria (LAB). Available online: https://www.iso.org/obp/ui/#iso:std:iso:10932:ed-1:v1:en (accessed on 24 May 2021).
- Grissa, I.; Vergnaud, G.; Pourcel, C. The CRISPRdb database and tools to display CRISPRs and to generate dictionaries of spacers and repeats. BMC Bioinform. 2007, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, R.; Bernhart, S.H.F.; Zu Siederdissen, C.H.; Tafer, H.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. ViennaRNA Package 2.0. Algorithms Mol. Biol. 2011, 6, 26. [Google Scholar] [CrossRef]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.; Rooney, A.; Yi, H.; Xu, X.-W.; De Meyer, S.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef]
- Gao, Y.; Li, D.; Liu, X. Bacteriocin-producing Lactobacillus sakei C2 as starter culture in fermented sausages. Food Control. 2014, 35, 1–6. [Google Scholar] [CrossRef]
- Malheiros, P.; Cuccovia, I.M.; Franco, B.D. Inhibition of Listeria monocytogenes in vitro and in goat milk by liposomal nanovesicles containing bacteriocins produced by Lactobacillus sakei subsp. sakei 2a. Food Control. 2016, 63, 158–164. [Google Scholar] [CrossRef]
- Ji, Y.; Park, S.; Chung, Y.; Kim, B.; Park, H.; Huang, E.; Jeong, D.; Jung, H.-Y.; Kim, B.; Hyun, C.-K.; et al. Amelioration of obesity-related biomarkers by Lactobacillus sakei CJLS03 in a high-fat diet-induced obese murine model. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-Y.; Park, B.-K.; Park, H.-J.; Park, Y.-H.; Kim, B.-O.; Pyo, S. Atopic dermatitis-mitigating effects of new Lactobacillus strain, Lactobacillus sakei probio 65 isolated from Kimchi. J. Appl. Microbiol. 2013, 115, 517–526. [Google Scholar] [CrossRef]
- Seo, S.; Shin, J.-S.; Lee, W.-S.; Rhee, Y.K.; Cho, C.-W.; Hong, H.-D.; Lee, K.-T. Anti-colitis effect of Lactobacillus sakei K040706 via suppression of inflammatory responses in the dextran sulfate sodium-induced colitis mice model. J. Funct. Foods 2017, 29, 256–268. [Google Scholar] [CrossRef]
- Eisenbach, L.; Geissler, A.J.; Ehrmann, M.A.; Vogel, R.F. Comparative genomics of Lactobacillus sakei supports the development of starter strain combinations. Microbiol. Res. 2019, 221, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yang, B.; Ross, R.P.; Stanton, C.; Zhao, J.; Zhang, H.; Chen, W. Comparative Genomics Analysis of Lactobacillus ruminis from Different Niches. Genes 2020, 11, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broadbent, J.R.; Neeno-Eckwall, E.C.; Stahl, B.; Tandee, K.; Cai, H.; Morovic, W.; Horvath, P.; Heidenreich, J.; Perna, N.T.; Barrangou, R.; et al. Analysis of the Lactobacillus casei supragenome and its influence in species evolution and lifestyle adaptation. BMC Genom. 2012, 13, 533. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Yang, B.; Ross, R.; Stanton, C.; Zhao, J.; Zhang, H.; Chen, W. Comparative Genomics of Pediococcus pentosaceus Isolated from Different Niches Reveals Genetic Diversity in Carbohydrate Metabolism and Immune System. Front. Microbiol. 2020, 11, 253. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, L.; Ross, P.; Zhao, J.; Zhang, H.; Chen, W. Comparative Genomics of Lactobacillus crispatus from the Gut and Vagina Reveals Genetic Diversity and Lifestyle Adaptation. Genes 2020, 11, 360. [Google Scholar] [CrossRef] [Green Version]
- O’Riordan, N.; O’Callaghan, J.; Buttò, L.; Kilcoyne, M.; Joshi, L.; Hickey, R. Bovine glycomacropeptide promotes the growth of Bifidobacterium longum ssp. infantis and modulates its gene expression. J. Dairy Sci. 2018, 101, 6730–6741. [Google Scholar] [CrossRef]
- Patryk, Z.; iley-Elkin, B.B.; Miriam, D.; Mark, B.L.; Permyakov, E. Structural and Biochemical Insights into the Peptidoglycan Hydrolase Domain of FlgJ from Salmonella typhimurium. PLoS ONE 2016, 11, e0149204. [Google Scholar] [CrossRef] [Green Version]
- Kameshwar, A.K.S.; Qin, W. Understanding the structural and functional properties of carbohydrate esterases with a special focus on hemicellulose deacetylating acetyl xylan esterases. Mycology 2018, 9, 273–295. [Google Scholar] [CrossRef] [Green Version]
- Salminen, S. Demonstration of safety of probiotics–A review. Int. J. Food Microbiol. 1998, 44, 93–106. [Google Scholar] [CrossRef]
- Georgieva, R.; Yocheva, L.; Tserovska, L.; Zhelezova, G.; Stefanova, N.; Atanasova, A.; Danguleva, A.; Ivanova, G.; Karapetkov, N.; Rumyan, N.; et al. Antimicrobial activity and antibiotic susceptibility ofLactobacillusandBifidobacteriumspp. intended for use as starter and probiotic cultures. Biotechnol. Biotechnol. Equip. 2014, 29, 84–91. [Google Scholar] [CrossRef]
- Botta, C.; Langerholc, T.; Cencic, A.; Cocolin, L. In Vitro Selection and Characterization of New Probiotic Candidates from Table Olive Microbiota. PLoS ONE 2014, 9, e94457. [Google Scholar] [CrossRef]
- Roberts, M.C.; Schwarz, S. Tetracycline and Phenicol Resistance Genes and Mechanisms: Importance for Agriculture, the Environment, and Humans. J. Environ. Qual. 2016, 45, 576–592. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, A.; Ono, N.; Akasaka, T.; Noumi, T.; Sawai, T.J.J.o.B.C. Metal-tetracycline/H+ antiporter of Escherichia coli encoded by a transposon, Tn10. The role of the conserved dipeptide, Ser65-Asp66, in tetracycline transport. J. Biol. Chem. 1990, 265, 15525–15530. [Google Scholar] [CrossRef]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR Provides Acquired Resistance Against Viruses in Prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Tyson, G.; Banfield, J. Rapidly evolving CRISPRs implicated in acquired resistance of microorganisms to viruses. Environ. Microbiol. 2007, 10, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Mougiakos, I.; Bosma, E.F.; de Vos, W.M.; van Kranenburg, R.; van der Oost, J. Next Generation Prokaryotic Engineering: The CRISPR-Cas Toolkit. Trends Biotechnol. 2016, 34, 575–587. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Source | Genome Size (Mb) | GC (%) | CDS No. | Reference |
|---|---|---|---|---|---|
| FAHWH17-L1 | Human feces | 1.90 | 41.82 | 1860 | This work |
| FJLHD1M1 | Human feces | 1.98 | 41.81 | 1969 | This work |
| FJLHD26M6 | Human feces | 1.98 | 41.79 | 1950 | This work |
| FXJWS8M1 | Human feces | 2.11 | 41.00 | 2142 | This work |
| FZJHZ2M8 | Human feces | 2.18 | 41.61 | 2085 | This work |
| GD179 | Human feces | 1.88 | 41.83 | 1856 | This work |
| FLEC01 | Human feces | 1.96 | 41.24 | 1919 | [25] |
| QYNXSBNJH59L1 | Sour cowhide | 1.94 | 41.77 | 1928 | This work |
| QJSSZ1L4 | Air-dried sausage | 1.90 | 41.92 | 1876 | This work |
| QJSNT1L10 | Air-dried sausage | 1.85 | 41.86 | 1829 | This work |
| QAHLA3L8 | Air-dried sausage | 1.90 | 42.03 | 1858 | This work |
| DSM15831 | Meat | 1.99 | 41.00 | 1924 | [26] |
| RI-496 | Meat | 2.02 | 40.90 | 1934 | [26] |
| RI-499 | Meat | 1.91 | 41.00 | 1838 | [26] |
| RI-500 | Meat | 2.03 | 40.90 | 1992 | [26] |
| RI-394 | Meat | 1.94 | 41.00 | 1963 | [27] |
| 23K | French sausage | 1.88 | 41.30 | 1779 | [20] |
| J64 | Dry sausage | 2.05 | 41.06 | 2102 | [25] |
| J156 | Dry sausage | 1.88 | 41.07 | 1817 | [25] |
| J112 | Dry sausage | 1.98 | 41.04 | 1913 | [25] |
| J54 | Dry sausage | 2.01 | 41.31 | 1982 | [25] |
| J18 | Dry sausage | 1.82 | 41.00 | 1826 | [25] |
| MFPB16A1401 | Beef carpaccio | 2.04 | 41.09 | 1960 | [25] |
| MFPB19 | Beef carpaccio | 2.06 | 41.05 | 1977 | [25] |
| J160 × 1 | Horse meat | 1.90 | 41.07 | 1857 | [25] |
| FAM18311 | Fermented meat products | 1.95 | 41.3 | 1896 | [28] |
| JXJ41 | Kimchi water | 2.04 | 41.80 | 2010 | This work |
| RS704 | Kimchi water | 1.96 | 41.86 | 1945 | This work |
| DS4 | Kimchi | 2.10 | 41.41 | 1970 | [26] |
| Probio65 | Kimchi | 2.08 | 41.18 | 1984 | [26] |
| WiKim0063 | Kimchi | 2.08 | 41.17 | 1955 | [26] |
| WiKim0072 | Kimchi | 2.03 | 41.17 | 1936 | [26] |
| WiKim0073 | Kimchi | 2.04 | 41.16 | 1913 | [26] |
| WiKim0074 | Kimchi | 2.04 | 41.03 | 1916 | [26] |
| WiKim22 | Kimchi | 2.19 | 40.61 | 1869 | [29] |
| CBA3614 | kimchi | 2.02 | 41.13 | 1910 | [30] |
| MBEL1397 | Kimchi | 1.99 | 41.04 | 1847 | [31] |
| FBL1 | Mukeunji | 2.03 | 41.20 | 1936 | [32] |
| QJSSZ4L10 | Pickled green vegetable heart | 2.05 | 41.70 | 2075 | This work |
| QJLYJ4L4 | Pickled radish | 1.97 | 41.76 | 1928 | This work |
| CECT9267 | Fermented potatos | 2.03 | 40.90 | 1972 | [26] |
| CECT9624 | Fermented potatos | 2.06 | 40.80 | 1986 | [33] |
| ZFM229 | Fermented vegetables | 2.02 | 41.22 | 1907 | [30] |
| LZ217 | Fermented vegetables | 2.02 | 41.22 | 1908 | [30] |
| RI-516 | Cacao bean | 1.91 | 41.00 | 1847 | [26] |
| RI-517 | Cacao bean | 1.95 | 41.00 | 1878 | [26] |
| LS25 | Commercial product Bitec LS-25 | 2.02 | 40.90 | 1972 | [34] |
| DSM20017 | Sake | 1.91 | 41.1 | 1820 | [26] |
| TMW1.411 | Starter culture | 1.94 | 41.06 | 1859 | [33] |
| LK-145 | Japanese sake cellar | 1.99 | 41.15 | 1901 | [35] |
| NBRC3541 | Raw moto-shu | 1.94 | 41.10 | 1855 | [36] |
| NBRC15893 | Kimoto | 1.90 | 41.10 | 1822 | [37] |
| ATCC15521 | Moto, starter of sake | 1.94 | 41.10 | 1835 | Unknown |
| ZFM220 | Raw cow milk | 2.02 | 41.22 | 1909 | [30] |
| ZFM225 | Raw cow milk | 2.02 | 41.22 | 1909 | [30] |
| AMBR8 | Nasopharyngeal samples | 2.00 | 41.00 | 1923 | [38] |
| RI-493 | Unknown | 1.97 | 40.90 | 1890 | [26] |
| CBA3635 | Unknown | 2.06 | 41.10 | 2002 | [30] |
| L15 | Unknown | 1.98 | 41.00 | 1915 | [30] |
| ye2 | Unknown | 1.99 | 41.10 | 1907 | [30] |
| RI-403 | Unknown | 2.00 | 41.00 | 2032 | [27] |
| RI-404 | Unknown | 1.95 | 40.90 | 1977 | [27] |
| RI-409 | Unknown | 1.99 | 41.00 | 2022 | [27] |
| RI-410 | Unknown | 1.93 | 41.10 | 1949 | [27] |
| RI-412 | Unknown | 1.92 | 41.10 | 1934 | [27] |
| MGYG-HGUT-02389 | Unknown | 1.96 | 41.20 | 1813 | Unknown |
| ERR260134-bin.14 | Unknown | 1.52 | 41.20 | - | Unknown |
| NBRC107868 | Unknown | 1.97 | 41.00 | 1920 | Unknown |
| Strain | Concentration (μg/mL) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| RD | S | K | CN | TE | DA | N | E | CIP | VA | C | |
| FJLHD1M1 | 1 | 256 | 256 | 1024 | 2 | 0.5 | 256 | 1 | 128 | 128 | 8 |
| GD17-9 | 1 | 256 | 128 | 1024 | 2 | 0.5 | 256 | 1 | 64 | 128 | 8 |
| FAHWH17-L1 | 0.5 | 512 | 256 | 1024 | 16 | 2 | 256 | 4 | 128 | 128 | 16 |
| JXJ4-1 | 1 | 512 | 128 | 1024 | 64 | 0.5 | 64 | 4 | 128 | 128 | 128 |
| QYNXSBNJH59L1 | 0.25 | 512 | 256 | 1024 | 4 | 0.5 | 256 | 2 | 128 | 128 | 32 |
| QJSSZ4L10 | 0.5 | 512 | 512 | 1024 | 8 | 0.25 | 128 | 4 | 128 | 128 | 32 |
| QJLYJ4L4 | 0.5 | 256 | 128 | 256 | 0 | 0.25 | 256 | 8 | 128 | 128 | 16 |
| QJSSZ1L4 | 1 | 512 | 256 | 1024 | 32 | 0.5 | 256 | 1 | 128 | 128 | 16 |
| QAHLA3L8 | 1 | 256 | 256 | 1024 | 32 | 0.5 | 256 | 2 | 64 | 128 | 32 |
| FJLHD26M6 | 0.25 | 256 | 256 | 512 | 4 | 0.125 | 32 | 2 | 128 | 128 | 8 |
| FXJWS8M1 | 8 | 256 | 256 | 1024 | 2 | 0.25 | 128 | 1 | 64 | 128 | 8 |
| FZJHZ2M8 | 1 | 256 | 512 | 256 | 64 | 4 | 256 | 2 | 128 | 128 | 16 |
| RS704 | 2 | 256 | 128 | 512 | 64 | 0.5 | 256 | 2 | 64 | 128 | 8 |
| QJSNT1L10 | 0.125 | 128 | 128 | 1024 | 64 | 0.0625 | 128 | 0.5 | 32 | 128 | 128 |
| Strains | Repeat Sequence (5’-3’) | CAS_ Subtype | Repeat Length | No. Spacer | Cas1 | Cas2 | Cas9 | Csn2 |
|---|---|---|---|---|---|---|---|---|
| FZJHZ2M8 | GTTTTAGAAGAGTATCAAATCAATGAGTAGTTCAAC | IIA | 36 | 16 | Y | Y | Y | Y |
| ATTTCATCTTAACCGAATGTTGAAGGAAACAATAGC | IIC | 36 | 71 | Y | N | Y | N | |
| QJSNT1L10 | GTTTTAGAAGAGTATCAAATCAATGAGTAGTTCAAC | IIA | 36 | 18 | N | N | N | Y |
| QAHLA3L8 | GTTGAACTACTCATTGATTTGATACTCTTCTAAAAC | IIA | 36 | 33 | Y | N | Y | Y |
| DS4 | GCTATTGTTTCCTTCAACATTCGGTTAAGATGAAAT | IIC | 36 | 26 | Y | Y | Y | N |
| J18 | GCTATTGTTTCCTTCAACATTCGGTTAAGATGAAAC | IIC | 36 | 92 | Y | Y | Y | N |
| CECT9624 | GTTTTAGAAGAGTATCAAATCAATGAGTAGTTCAAC | IIA | 36 | 8 | Y | Y | Y | Y |
| ATTTCATCTTAACCGAATGTTGAAGGAAACA | IIC | 31 | 2 | Y | Y | Y | N | |
| J54 | GTTGAACTACTCATTGATTTGATACTCTTCTAAAAC | IIA | 36 | 29 | Y | Y | Y | Y |
| J112 | GTTTTAGAAGAGTATCAAATCAATGAGTAGTTCAAC | IIA | 36 | 14 | Y | Y | Y | N |
| FLEC01 | GTTGAACTACTCATTGATTTGATACTCTTCTAAAAC | IIA | 36 | 35 | Y | Y | Y | Y |
| J160 × 1 | GTTGAACTACTCATTGATTTGATACTCTTCTAAAAC | IIA | 36 | 24 | Y | Y | Y | Y |
| MGYG-HGUT-02389 | GTTGAACTACTCATTGATTTGATACTCTTCTAAAAC | IIA | 36 | 35 | Y | Y | Y | Y |
| DSM15831 | AGTTGAACCACTCATTGATTTGATACTCTTCTAAAAC | IIA | 37 | 11 | N | N | N | Y |
| RI-403 | GTTTTAGAAGAGTATCAAATCAATGAGTGGTTCAAC | IIA | 36 | 40 | N | N | N | Y |
| RI-404 | GTTTTAGAAGAGTATCAAATCAATGAGTGGTTCAAC | IIA | 36 | 37 | Y | Y | Y | Y |
| AMBR8 | GTTGAACCACTCATTGATTTGATACTCTTCTAAAAC | IIA | 36 | 23 | Y | Y | Y | Y |
| NBRC107868 | GTTTTAGAAGAGTATCAAATCAATGAGTGGTTCAACT | IIA | 37 | 11 | N | N | N | Y |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Li, N.; Zhao, S.; Zhang, C.; Qiao, N.; Duan, H.; Xiao, Y.; Yan, B.; Zhao, J.; Tian, F.; et al. Integrated Phenotypic–Genotypic Analysis of Latilactobacillus sakei from Different Niches. Foods 2021, 10, 1717. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10081717
Chen Y, Li N, Zhao S, Zhang C, Qiao N, Duan H, Xiao Y, Yan B, Zhao J, Tian F, et al. Integrated Phenotypic–Genotypic Analysis of Latilactobacillus sakei from Different Niches. Foods. 2021; 10(8):1717. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10081717
Chicago/Turabian StyleChen, Ying, Nan Li, Shenxi Zhao, Chuan Zhang, Nanzhen Qiao, Hui Duan, Yue Xiao, Bowen Yan, Jianxin Zhao, Fengwei Tian, and et al. 2021. "Integrated Phenotypic–Genotypic Analysis of Latilactobacillus sakei from Different Niches" Foods 10, no. 8: 1717. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10081717