
Screening of Regulated and Emerging Mycotoxins in Bulk Milk Samples by High-Resolution Mass Spectrometry



Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Milk Samples
2.2. Extraction Step for UHPLC-HRMS Analysis
2.3. Screening of Mycotoxins by UHPLC-HRMS Analysis
2.4. Data Processing
2.5. Multivariate Statistical Analysis
3. Results and Discussion
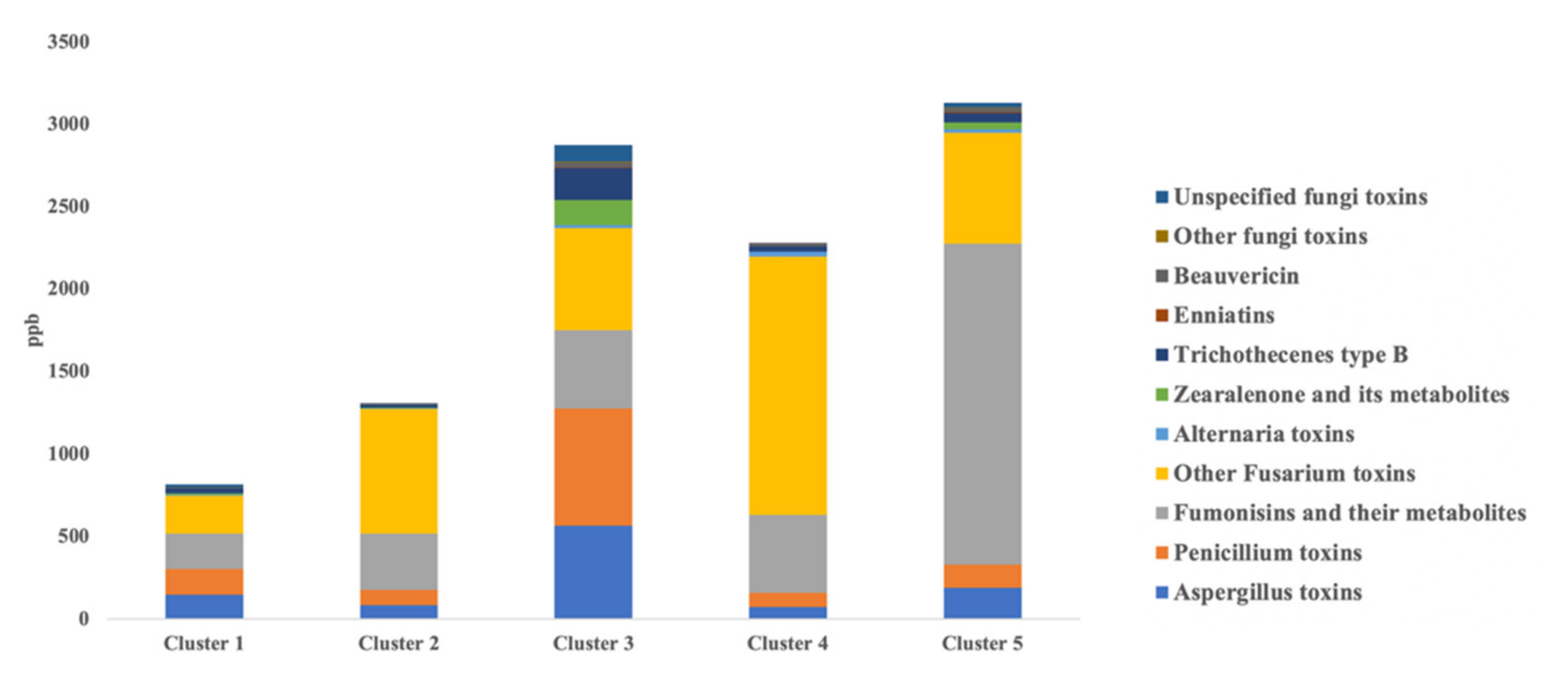
3.1. Profiling of Mycotoxins in the Different Milk Samples
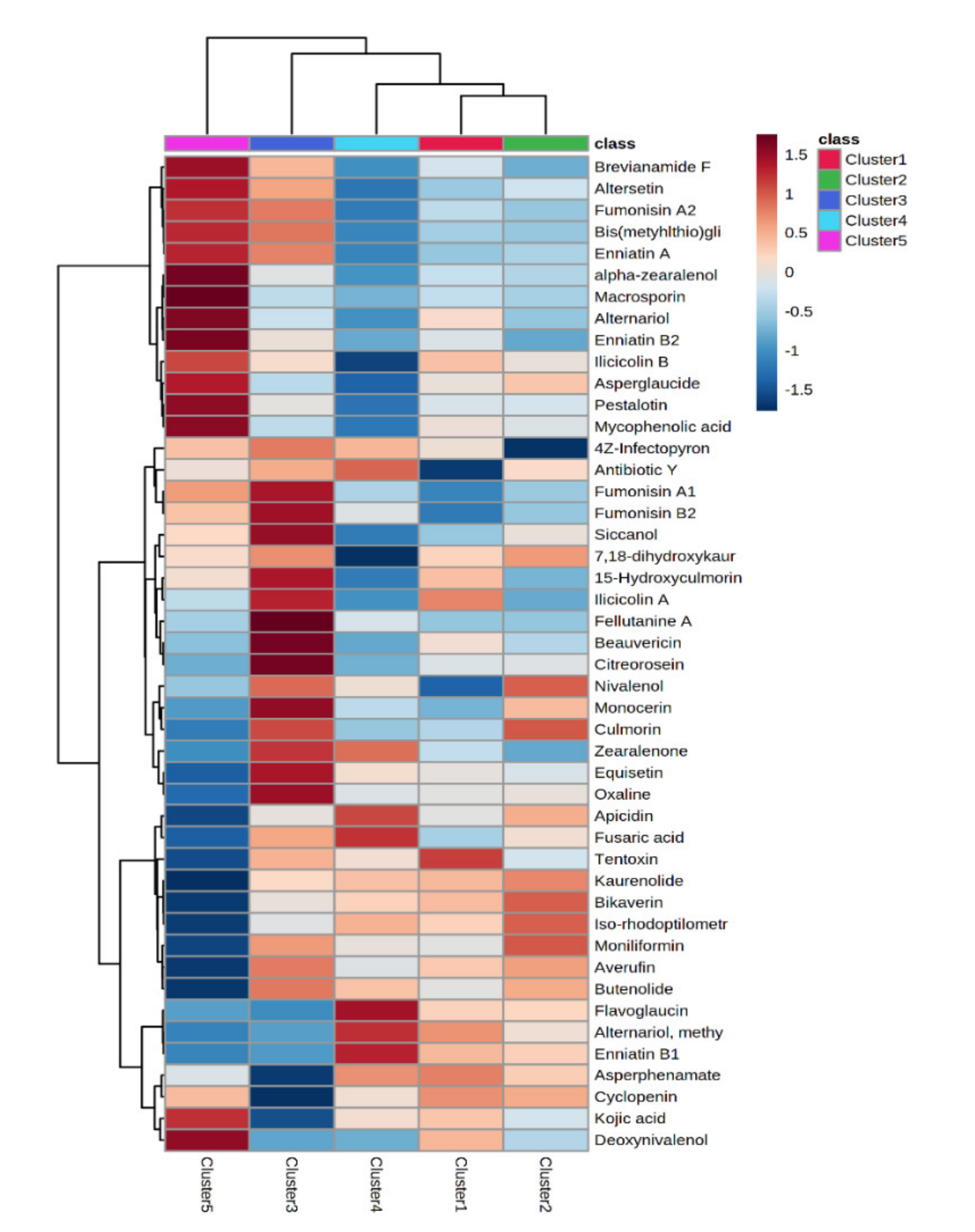
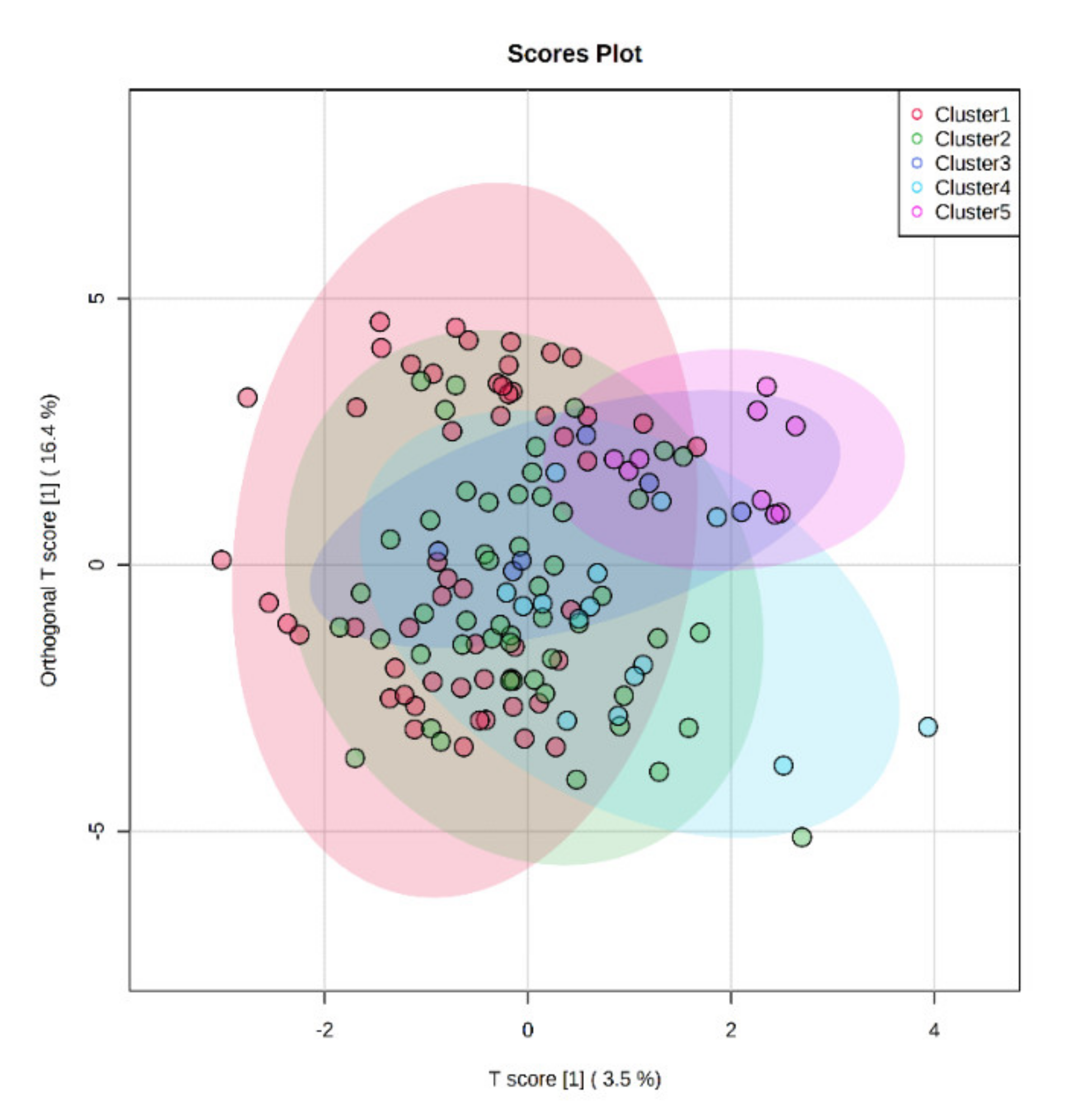
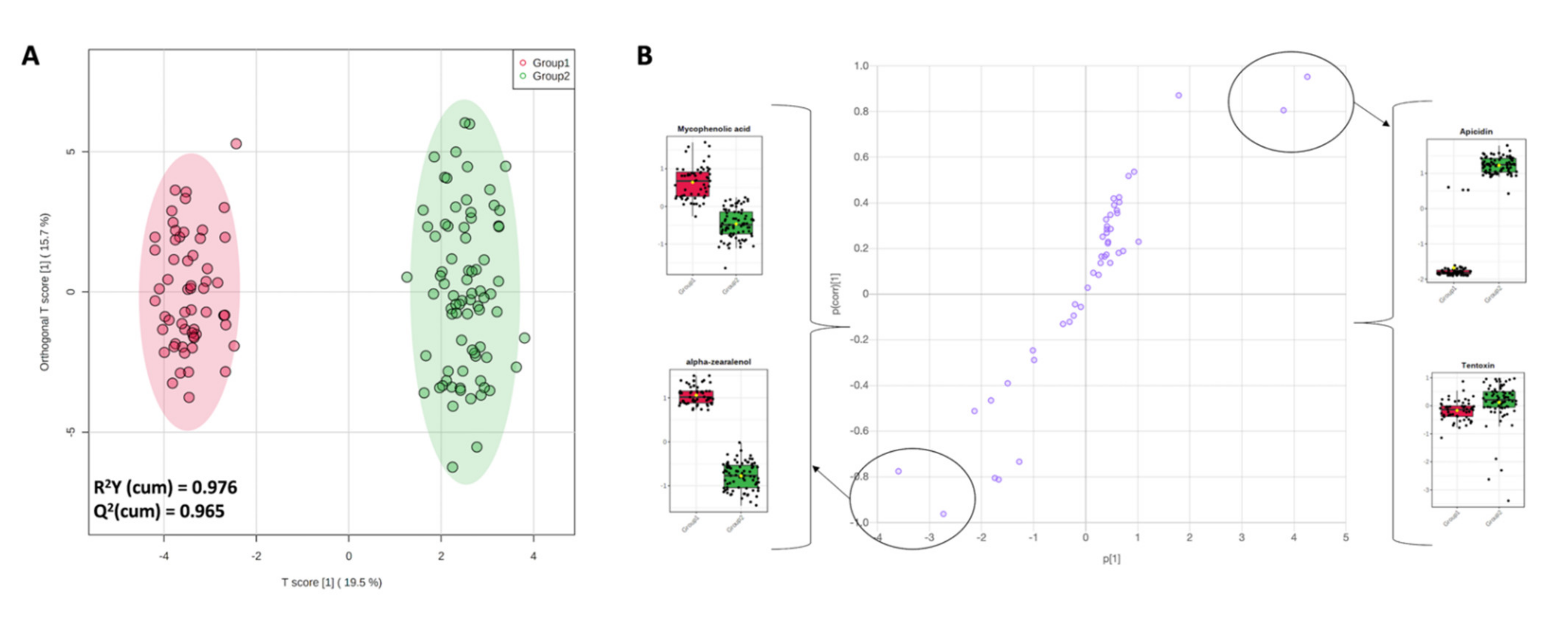
3.2. Multivariate Analysis on the Different Milk Samples and Discriminant Metabolites
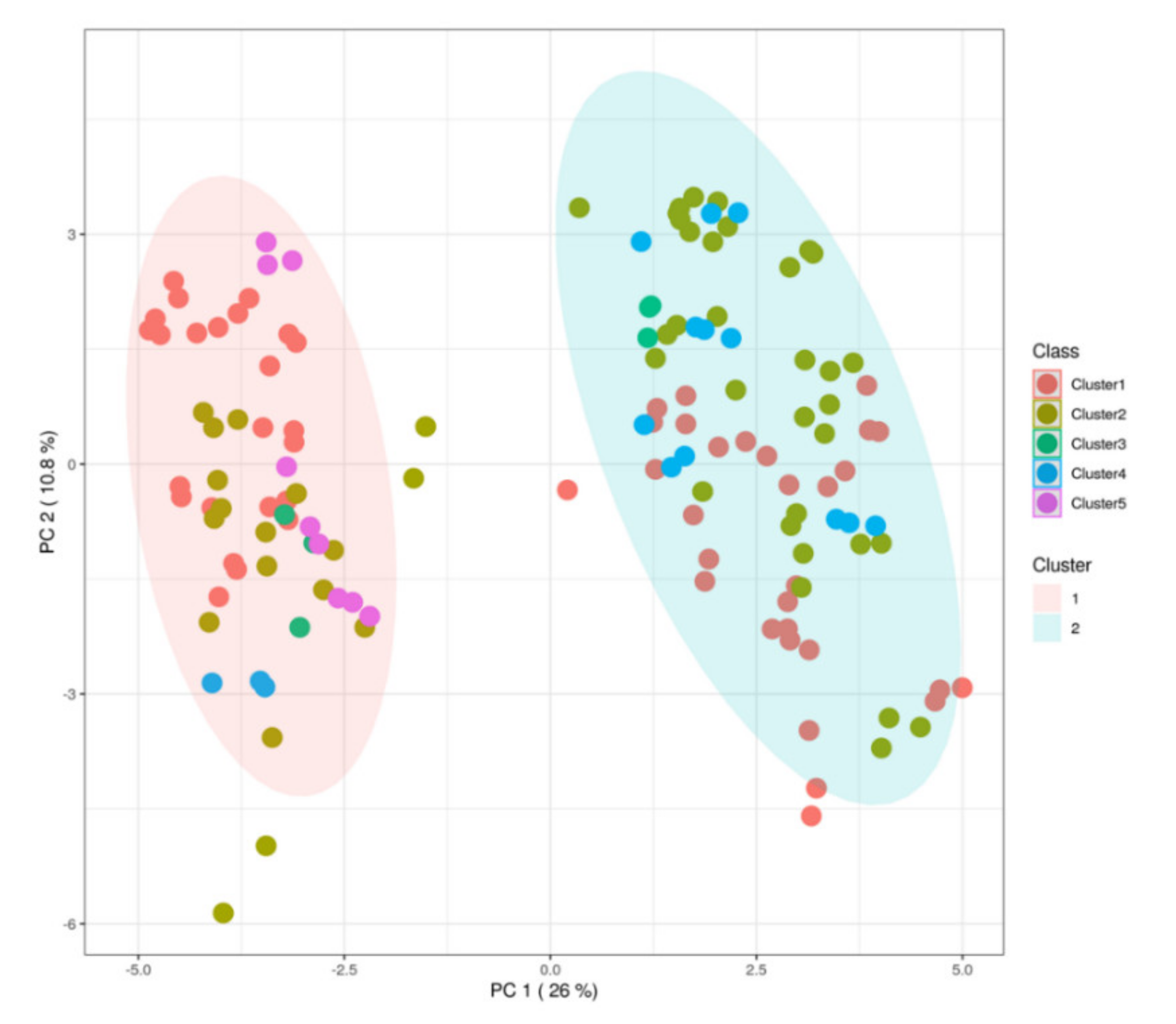
3.3. Discrimination of Milk Samples According to a k-Means Clustering Approach
3.4. Semi-Quantitative Analysis by UHPLC-HRMS of the Discriminant Markers
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, D.; Kebede, B.; McComb, K.; Hayman, A.; Chen, G.; Frew, R. Milk Biomarkers in Relation to Inherent and External Factors Based on Metabolomics. Trends Food. Sci. Technol. 2021, 109, 51–64. [Google Scholar] [CrossRef]
- Rocchetti, G.; O’Callaghan, T.F. Application of Metabolomics to Assess Milk Quality and Traceability. Curr. Opin. Food Sci. 2021, 40, 168–178. [Google Scholar] [CrossRef]
- Joubran, A.M.; Pierce, K.M.; Garvey, N.; Shalloo, L.; O’Callaghan, T.F. Invited Review: A 2020 Perspective on Pasture-Based Dairy Systems and Products. J. Dairy Sci. 2021, 104, 7364–7382. [Google Scholar] [CrossRef]
- Gallo, A.; Fancello, F.; Ghilardelli, F.; Zara, S.; Froldi, F.; Spanghero, M. Effects of Several Lactic Acid Bacteria Inoculants on Fermentation and Mycotoxins in Corn Silage. Anim. Feed Sci. Technol. 2021, 277, 114962. [Google Scholar] [CrossRef]
- Gallo, A.; Ghilardelli, F.; Atzori, A.S.; Zara, S.; Novak, B.; Faas, J.; Fancello, F. Co-Occurrence of Regulated and Emerging Mycotoxins in Corn Silage: Relationships with Fermentation Quality and Bacterial Communities. Toxins 2021, 13, 232. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Giuberti, G.; Frisvad, J.C.; Bertuzzi, T.; Nielsen, K.F. Review on Mycotoxin Issues in Ruminants: Occurrence in Forages, Effects of Mycotoxin Ingestion on Health Status and Animal Performance and Practical Strategies to Counteract Their Negative Effects. Toxins 2015, 7, 3057–3111. [Google Scholar] [CrossRef]
- Liew, W.-P.-P.; Mohd-Redzwan, S. Mycotoxin: Its Impact on Gut Health and Microbiota. Front. Cell. Infect. Microbiol. 2018, 8, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fink-Gremmels, J. Mycotoxins in Cattle Feeds and Carry-Over to Dairy Milk: A Review. Food Addit. Contam. Part A 2008, 25, 172–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemboi, D.C.; Antonissen, G.; Ochieng, P.E.; Croubels, S.; Okoth, S.; Kangethe, E.K.; Faas, J.; Lindahl, J.F.; Gathumbi, J.K. A Review of the Impact of Mycotoxins on Dairy Cattle Health: Challenges for Food Safety and Dairy Production in Sub-Saharan Africa. Toxins 2020, 12, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogunade, I.M.; Martinez-Tuppia, C.; Queiroz, O.C.M.; Jiang, Y.; Drouin, P.; Wu, F.; Vyas, D.; Adesogan, A.T. Silage Review: Mycotoxins in Silage: Occurrence, Effects, Prevention, and Mitigation. J. Dairy Sci. 2018, 101, 4034–4059. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.C.; Zheng, N.; Zheng, B.Q.; Wen, F.; Cheng, J.B.; Han, R.W.; Xu, X.M.; Li, S.L.; Wang, J.Q. Simultaneous Determination of Aflatoxin M1, Ochratoxin A, Zearalenone and α-zearalenol in Milk by UHPLC–MS/MS. Food Chem. 2014, 146, 242–249. [Google Scholar] [CrossRef]
- Izzo, L.; Rodríguez-Carrasco, Y.; Tolosa, J.; Graziani, G.; Gaspari, A.; Ritieni, A. Target Analysis and Retrospective Screening of Mycotoxins and Pharmacologically Active Substances in Milk Using an Ultra-High-Performance Liquid Chromatography/High-Resolution Mass Spectrometry Approach. J. Dairy Sci. 2020, 103, 1250–1260. [Google Scholar] [CrossRef]
- Tamura, M.; Mochizuki, N.; Nagatomi, Y.; Harayama, K.; Toriba, A.; Hayakawa, K. A Method for Simultaneous Determination of 20 Fusarium Toxins in Cereals by High-Resolution Liquid Chromatography-Orbitrap Mass Spectrometry with a Pentafluorophenyl Column. Toxins 2015, 7, 1664–1682. [Google Scholar] [CrossRef] [PubMed]
- Castaldo, L.; Graziani, G.; Gaspari, A.; Izzo, L.; Tolosa, J.; Rodríguez-Carrasco, Y.; Ritieni, A. Target Analysis and Retrospective Screening of Multiple Mycotoxins in Pet Food Using UHPLC-Q-Orbitrap HRMS. Toxins 2019, 11, 434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Dominicis, E.; Commissati, I.; Gritti, E.; Catellani, D.; Suman, M. Quantitative Targeted and Retrospective Data Analysis of Relevant Pesticides, Antibiotics and Mycotoxins in Bakery Products by Liquid Chromatography-Single-Stage Orbitrap Mass Spectrometry. Food Addit. Contam. Part A 2015, 32, 1617–1627. [Google Scholar] [CrossRef] [PubMed]
- Rocchetti, G.; Ghilardelli, F.; Bonini, P.; Lucini, L.; Masoero, F.; Gallo, A. Changes of Milk Metabolomic Profiles Resulting from a Mycotoxins-Contaminated Corn Silage Intake by Dairy Cows. Metabolites 2021, 11, 475. [Google Scholar] [CrossRef]
- Rocchetti, G.; Gallo, A.; Nocetti, M.; Lucini, L.; Masoero, F. Milk Metabolomics Based on Ultra-High-Performance Liquid Chromatography Coupled with Quadrupole Time-of-Flight Mass Spectrometry to Discriminate Different Cows Feeding Regimens. Food Res. Int. 2020, 134, 109279. [Google Scholar] [CrossRef]
- Bellassi, P.; Rocchetti, G.; Nocetti, M.; Lucini, L.; Masoero, F.; Morelli, L. A Combined Metabolomic and Metagenomic Approach to Discriminate Raw Milk for the Production of Hard Cheese. Foods 2021, 10, 109. [Google Scholar] [CrossRef]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; Vander Gheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-Independent MS/MS Deconvolution for Comprehensive Metabolome Analysis. Nat. Methods. 2015, 12, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, H.; Kind, T.; Nakabayashi, R.; Yukihira, D.; Tanaka, W.; Cajka, T.; Saito, K.; Fiehn, O.; Arita, M. Hydrogen Rearrangement Rules: Computational MS/MS Fragmentation and Structure Elucidation Using MS-FINDER Software. Anal. Chem. 2016, 88, 7946–7958. [Google Scholar] [CrossRef]
- Salek, R.M.; Steinbeck, C.; Viant, M.R.; Goodacre, R.; Dunn, W.B. The Role of Reporting Standards for Metabolite Annotation and Identification in Metabolomic Studies. GigaScience 2013, 2, 2047-217X. [Google Scholar] [CrossRef] [PubMed]
- Zachariasova, M.; Cajka, T.; Godula, M.; Malachova, A.; Veprikova, Z.; Hajslova, J. Analysis of Multiple Mycotoxins in Beer Employing (Ultra)-High-Resolution Mass Spectrometry. Rapid. Commun. Mass Spectrom. 2010, 24, 3357–3367. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the Gap Between Raw Spectra and Functional Insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Chong, J.; Li, S.; Xia, J. MetaboAnalystR 3.0: Toward an Optimized Workflow for Global Metabolomics. Metabolites 2020, 10, 186. [Google Scholar] [CrossRef]
- Escrivá, L.; Oueslati, S.; Font, G.; Manyes, L. Alternaria Mycotoxins in Food and Feed: An Overview. J. Food Qual. 2017, 2017, 1569748. [Google Scholar] [CrossRef] [Green Version]
- Arcella, D.; Eskola, M.; Gómez Ruiz, J.A. Dietary Exposure Assessment to Alternaria Toxins in the European Population. EFSA J. 2016, 14, e04654. [Google Scholar]
- Gotthardt, M.; Asam, S.; Gunkel, K.; Moghaddam, A.F.; Baumann, E.; Kietz, R.; Rychlik, M. Quantitation of Six Alternaria Toxins in Infant Foods Applying Stable Isotope Labeled Standards. Front. Microbiol. 2019, 10, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arie, T. Fusarium diseases of cultivated plants, control, diagnosis, and molecular and genetic studies. J. Pest. Sci. 2019, 44, 275–281. [Google Scholar] [CrossRef] [Green Version]
- Minervini, F.; Dell’Aquila, M.E. Zearalenone and Reproductive Function in Farm Animals. Int. J. Mol. Sci. 2008, 9, 2570–2584. [Google Scholar] [CrossRef] [Green Version]
- Keller, L.; Abrunhosa, L.; Keller, K.; Rosa, C.A.; Cavaglieri, L.; Venâncio, A. Zearalenone and Its Derivatives α-Zearalenol and β-Zearalenol Decontamination by Saccharomyces cerevisiae Strains Isolated from Bovine Forage. Toxins 2015, 7, 3297–3308. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, K.; Passeró, E.; Vallone, L.; Hymery, N.; Coton, M.; Jany, J.-L.; Mounier, J.; Coton, E. Occurrence of Roquefortine C, Mycophenolic Acid and Aflatoxin M1 Mycotoxins in Blue-Veined Cheeses. Food Cont. 2015, 47, 634–640. [Google Scholar] [CrossRef]
- Martínez-Iglesias, O.; Cacabelos, R. Chapter 12—Epigenetic Treatment of Neurodegenerative disorders. Histone Modif. Ther. 2020, 20, 311–335. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Compound | p-Value (FWER) | VIP Score (OPLS-DA) | LogFC Cluster 3 vs. Cluster 1 | LogFC Cluster 3 vs. Cluster 2 | LogFC Cluster 4 vs. Cluster 1 | LogFC Cluster 4 vs. Cluster 2 | LogFC Cluster 5 vs. Cluster 1 | LogFC Cluster 5 vs. Cluster 2 |
|---|---|---|---|---|---|---|---|---|---|
| Alternaria mycotoxins | Altersetin | p > 0.05 | <1 | 2.151 | 1.562 | −1.466 | −2.056 | 3.343 | 2.753 |
| Alternariol | p > 0.05 | <1 | −0.064 | 0.113 | −0.341 | −0.163 | −0.097 | 0.080 | |
| Alternariol, methyl-ether | 0.0011 | <1 | −0.810 | −0.433 | 0.139 | 0.516 | −1.273 | −0.897 | |
| Tentoxin | 0.0495 | 1.119 | −0.158 | 0.272 | −0.492 | −0.061 | −1.258 | −0.827 | |
| 4Z-Infectopyron | 0.0354 | <1 | 0.298 | 1.072 | 0.002 | 0.774 | −0.210 | 0.562 | |
| Aspergillus mycotoxins | Brevianamide F | 0.0065 | 1.287 | 0.307 | 0.671 | −0.584 | −0.220 | 0.501 | 0.865 |
| Kojic acid | p > 0.05 | <1 | −0.542 | −0.513 | −0.250 | −0.222 | −0.267 | −0.239 | |
| Averufin | p > 0.05 | <1 | 0.531 | 0.129 | 0.080 | −0.321 | −0.640 | −1.042 | |
| Bis(methylthio)gliotoxin | 0.0066 | 1.102 | 5.414 | 6.242 | −2.603 | −1.775 | 6.830 | 7.657 | |
| Asperphenamate | p > 0.05 | <1 | −5.604 | −4.700 | −1.351 | −0.447 | −2.126 | −1.222 | |
| Fumonisins mycotoxins | Fumonisin A1 | 0.0494 | 1.667 | 2.069 | 1.671 | 0.412 | 0.014 | 1.084 | 0.687 |
| Fumonisin A2 | 0.0024 | <1 | 3.285 | 4.041 | −4.203 | −3.447 | 4.114 | 4.870 | |
| Fumonisin B2 | 0.0294 | 1.846 | 2.444 | 1.918 | 0.858 | 0.332 | 1.096 | 0.570 | |
| Zearalenone and metabolites | Zearalenone | 0.0176 | 1.358 | 0.420 | 0.632 | 0.175 | 0.386 | −0.543 | −0.332 |
| α-Zearalenol | 0.0378 | 1.098 | 0.416 | 0.666 | −1.264 | −1.014 | 2.822 | 3.072 | |
| Trichothecenes | Deoxynivalenol | p > 0.05 | <1 | −0.396 | −0.080 | −0.530 | −0.214 | 0.00072 | 0.316 |
| Nivalenol | p > 0.05 | 1.114 | 0.198 | 0.053 | −0.029 | −0.174 | −0.265 | −0.410 | |
| Other Fusarium mycotoxins | Siccanol | p > 0.05 | <1 | 0.335 | 0.303 | −0.261 | −0.293 | −0.214 | −0.246 |
| Monocerin | p > 0.05 | <1 | 0.460 | 0.287 | −0.072 | −0.246 | −0.376 | −0.550 | |
| Moniliformin | 5.2 × 10−6 | 1.012 | 0.442 | −0.187 | −0.119 | −0.748 | −1.359 | −1.988 | |
| Equisetin | p > 0.05 | <1 | 1.166 | 2.234 | −0.060 | 1.006 | −1.493 | −0.426 | |
| Culmorin | p > 0.05 | <1 | 0.243 | 0.071 | −0.185 | −0.357 | −0.467 | −0.639 | |
| 15-Hydroxyculmorin | p > 0.05 | <1 | 0.137 | 0.352 | −0.377 | −0.161 | −0.376 | −0.161 | |
| Butenolide | p > 0.05 | <1 | 0.116 | 0.097 | −0.099 | −0.118 | −0.560 | −0.578 | |
| Bikaverin | 3.4 × 10−4 | 1.975 | −1.630 | −3.375 | −0.734 | −2.478 | −9.655 | −11.400 | |
| Apicidin | p > 0.05 | <1 | 0.104 | −1.462 | 3.163 | 1.596 | −4.700 | −6.268 | |
| Antibiotic Y | 0.0065 | 2.579 | 3.395 | 1.069 | 0.771 | −1.554 | 2.315 | −0.010 | |
| Kaurenolide | 0.0024 | 1.581 | −0.178 | −0.334 | −0.198 | −0.354 | −1.845 | −2.001 | |
| 7,1-Dihydroxykaurenolide | p > 0.05 | <1 | 0.954 | 1.572 | −1.999 | −1.381 | −0.011 | 0.606 | |
| Fusaric acid | p > 0.05 | <1 | 0.250 | 0.175 | 0.264 | 0.189 | −0.578 | −0.653 | |
| Enniatins-Beauvericin toxins | Enniatin A | p > 0.05 | <1 | 0.647 | 0.640 | −0.418 | −0.425 | 0.564 | 0.557 |
| Enniatin B1 | p > 0.05 | <1 | −1.421 | −0.724 | 0.767 | 1.464 | −1.944 | −1.248 | |
| Enniatin B2 | 0.0434 | 1.039 | 0.058 | 0.436 | −0.466 | −0.088 | 0.458 | 0.836 | |
| Beauvericin | 0.0024 | <1 | 0.438 | 0.656 | −0.429 | −0.211 | −0.586 | −0.368 | |
| Penicillium mycotoxins | Asperglaucide | p > 0.05 | <1 | −0.452 | −0.802 | −1.913 | −2.263 | 1.347 | 0.997 |
| Pestalotin | p > 0.05 | <1 | 0.088 | 0.167 | −1.029 | −0.951 | 0.981 | 1.0595 | |
| Oxaline | p > 0.05 | <1 | 0.428 | 0.475 | −0.174 | −0.127 | −0.699 | −0.652 | |
| Flavoglaucin | 0.0111 | <1 | −1.088 | −0.992 | 0.842 | 0.938 | −1.288 | −1.192 | |
| Cyclopenin | p > 0.05 | <1 | −0.515 | −0.420 | −0.281 | −0.187 | −0.393 | −0.299 | |
| Fellutanine A | p > 0.05 | <1 | 1.067 | 1.129 | 0.037 | 0.099 | −0.283 | −0.221 | |
| Mycophenolic acid | 0.0147 | <1 | −0.312 | −0.094 | −1.394 | −1.175 | 1.296 | 1.515 | |
| Other fungal metabolites | Ilicicolin A | 0.0146 | 1.706 | 2.421 | 6.086 | −4.303 | −0.639 | −2.547 | 1.117 |
| Ilicicolin B | 0.0467 | <1 | −0.086 | 0.734 | −3.767 | −2.946 | −1.029 | −0.208 | |
| Citreorosein | 0.0369 | <1 | 5.858 | 5.831 | −2.225 | −2.252 | −2.510 | −2.538 | |
| Macrosporin | 1.6 × 10−13 | <1 | −4.451 | −4.295 | −0.415 | −0.260 | −4.296 | −4.147 | |
| Iso-Rhodoptilometrin | 3.2 × 10−5 | 1.254 | −0.162 | −0.462 | −0.042 | −0.341 | −1.301 | −1.599 |
| Group (k-Means) | Milk Sample | Tentoxin (ng/mL) | α-Zearalenol (ng/mL) | Mycophenolic Acid (ng/mL) | Apicidin (ng/mL) |
|---|---|---|---|---|---|
| Group 1 (left side) | Sample 2 | 0.86 | 2.44 | 1.27 | nd |
| Sample 6 | 1.68 | 3.81 | 1.08 | nd | |
| Sample 11 | 1.71 | 2.91 | 0.95 | nd | |
| Sample 14 | 1.03 | 2.31 | 0.31 | nd | |
| Sample 16 | 0.64 | 1.79 | 0.52 | nd | |
| Sample 20 | 2.21 | 3.06 | 1.19 | nd | |
| Sample 43 | 1.08 | 2.33 | 0.35 | nd | |
| Sample 45 | 2.04 | 3.35 | 0.34 | nd | |
| Sample 10 | 1.69 | 4.94 | 0.48 | nd | |
| Sample 13 | 1.93 | 2.79 | 0.49 | nd | |
| Sample 21 | 1.18 | 5.25 | 2.68 | nd | |
| Sample 23 | 0.67 | 1.06 | 0.46 | nd | |
| Sample 26 | 0.71 | 2.13 | 0.50 | nd | |
| Sample 32 | 1.16 | 2.90 | 0.79 | nd | |
| Sample 5 | 1.07 | 2.90 | 0.30 | nd | |
| Sample 27 | 1.07 | 1.96 | 0.62 | nd | |
| Sample 4 | 0.76 | 1.75 | 0.28 | nd | |
| Sample 28 | 1.22 | 4.93 | 1.38 | <0.1 | |
| Sample 41 | 0.87 | 1.80 | 0.99 | nd | |
| Group 2 (right side) | Sample 19 | 1.01 | <0.1 | <0.1 | 0.33 |
| Sample 22 | 1.16 | <0.1 | <0.1 | 0.23 | |
| Sample 25 | 2.35 | <0.1 | 0.19 | 0.28 | |
| Sample 29 | 1.85 | <0.1 | <0.1 | 0.15 | |
| Sample 33 | 1.05 | <0.1 | 0.16 | 0.11 | |
| Sample 36 | 1.79 | <0.1 | <0.1 | 0.57 | |
| Sample 37 | 2.07 | <0.1 | <0.1 | 0.27 | |
| Sample 38 | 2.35 | <0.1 | <0.1 | 0.24 | |
| Sample 39 | 1.16 | <0.1 | 0.15 | 0.32 | |
| Sample 40 | 2.06 | <0.1 | <0.1 | <0.1 | |
| Sample 1 | 0.29 | <0.1 | <0.1 | 0.13 | |
| Sample 3 | 0.19 | <0.1 | <0.1 | 0.24 | |
| Sample 7 | 1.49 | <0.1 | <0.1 | 0.20 | |
| Sample 15 | 1.15 | <0.1 | <0.1 | 0.44 | |
| Sample 17 | 1.28 | <0.1 | <0.1 | <0.1 | |
| Sample 24 | 1.32 | <0.1 | <0.1 | 0.43 | |
| Sample 30 | 1.71 | <0.1 | <0.1 | 0.18 | |
| Sample 31 | 2.42 | <0.1 | 0.15 | 0.41 | |
| Sample 34 | 2.02 | <0.1 | <0.1 | 0.55 | |
| Sample 42 | 2.23 | <0.1 | <0.1 | 0.14 | |
| Sample 44 | 2.88 | <0.1 | <0.1 | 0.41 | |
| Sample 8 | 1.43 | <0.1 | <0.1 | 0.71 | |
| Sample 9 | <0.1 | <0.1 | <0.1 | 0.30 | |
| Sample 12 | 1.01 | <0.1 | <0.1 | 0.15 | |
| Sample 18 | 2.06 | <0.1 | <0.1 | 0.12 | |
| Sample 46 | 1.46 | <0.1 | <0.1 | 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rocchetti, G.; Ghilardelli, F.; Masoero, F.; Gallo, A. Screening of Regulated and Emerging Mycotoxins in Bulk Milk Samples by High-Resolution Mass Spectrometry. Foods 2021, 10, 2025. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10092025
Rocchetti G, Ghilardelli F, Masoero F, Gallo A. Screening of Regulated and Emerging Mycotoxins in Bulk Milk Samples by High-Resolution Mass Spectrometry. Foods. 2021; 10(9):2025. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10092025
Chicago/Turabian StyleRocchetti, Gabriele, Francesca Ghilardelli, Francesco Masoero, and Antonio Gallo. 2021. "Screening of Regulated and Emerging Mycotoxins in Bulk Milk Samples by High-Resolution Mass Spectrometry" Foods 10, no. 9: 2025. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10092025