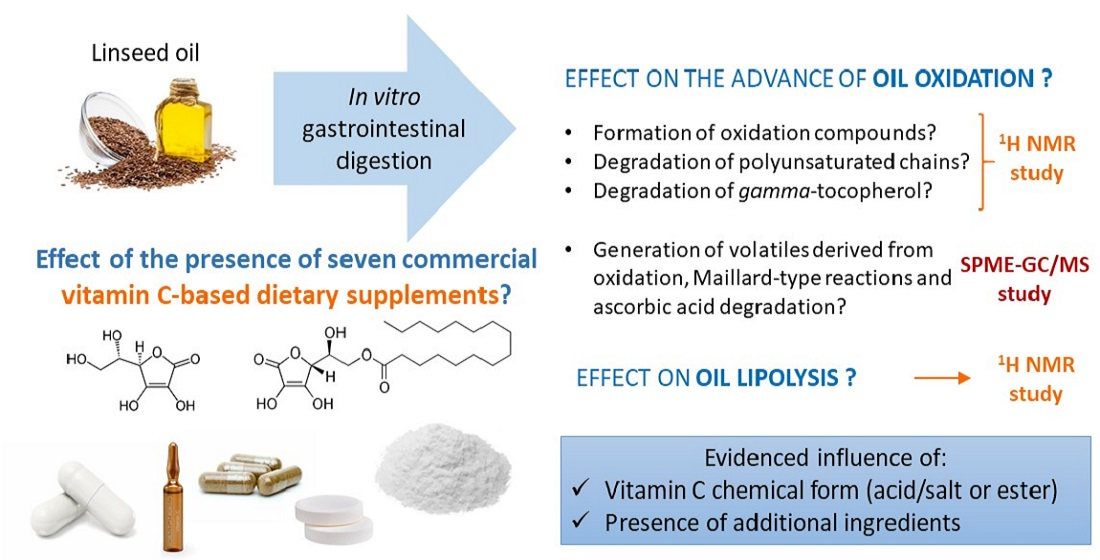
3.1.1. Effect on the Formation of Oxidation Compounds That Can Be Detected through 1H NMR
Figure 1 shows certain regions of the multisuppressed
1H NMR spectra of linseed oil (L) together with those of the lipid extracts obtained after its in vitro digestion in the absence (DL) and in the presence of either ascorbic acid/salt (DLAsc1–2 and DLVC1–6) or ascorbyl palmitate (DLAP and DLVC7), in which proton signals of primary and secondary oxidation products can be observed. The assignment of these signals is provided in
Table 2, and the estimated concentrations of the corresponding oxidation products detected in several lipidic samples, given in μmol/mol AG + FA, are shown in
Table 3.
Figure 1 reveals that the oil subject of study (L) already showed a very slight degree of oxidation before being submitted to digestion. This is deduced from the presence in its multisuppressed
1H NMR spectrum of signals
a and
h, attributable to
Z,E-conjugated dienes supported in octadecatrienoic fatty acyl chains also comprising a hydroxy group (
Z,E-CD-OH) and to the aldehydic proton of alkanals, respectively. Their estimated concentrations (see
Table 3) can be considered very low and similar to those found in other fresh edible oils rich in polyunsaturated acyl groups [
17].
During the in vitro digestion of this highly polyunsaturated oil, not only the concentrations of these types of oxidation compounds increased (
p > 0.05) slightly (see DL sample in
Figure 1 and
Table 3), but the generation of hydroperoxy-
Z,
E-conjugated dienes (
Z,
E-CD-OOH) derived from linolenic groups was also noticed (see signal
b in DL spectrum), the latter being oxidation products formed in the highest proportion, in agreement with previous results [
17,
18].
The mixture of linseed oil with ascorbic acid, either pure (Asc1–2) or in the form of supplements containing basically this compound (VC1–4), provoked a clear enhancement of linseed oil oxidation during in vitro digestion. Thus, in the spectra corresponding to the lipids of the in vitro digests of the previously mentioned samples (DLAsc1–2 and DLVC1–4), increased intensities of the signals due to
Z,E-CD-OH,
Z,E-CD-OOH and alkanals (signals
a,
b and
h, respectively) in comparison with DL spectrum were observed (see also concentrations in
Table 3). Moreover, new signals attributable to protons of
Z,
E/
E,
Z-conjugated dienes of keto-octadecatrienoic structures were also detected (see signal
c in
Figure 1). In addition, the generation of several kinds of alpha,beta-unsaturated aldehydes was also favored, inferred from the appearance of doublets
d,
e,
f and
g due to the aldehydic proton of
E-2-alkenals,
E,
E-2,4-alkadienals, 4,5-epoxy-
E-2-alkenals and
E,
Z-2,4-alkadienals, respectively. Among them, the high reactivity of the oxygenated 4,5-epoxy-
E-2-alkenals should be noticed [
41]. These results are in line with those of Lee et al. [
42], according to which Vitamin C (in the form of ascorbic acid) was very effective in decomposing linoleic acid-derived hydroperoxides to genotoxins, among which 4,5-epoxy-
E-2-decenal was present.
Overall, as shown in
Table 3, the estimated concentration of total conjugated dienic structures present in long chains in linseed oil digests was approximately 1.5–2-fold higher in DLAsc1–2 and DLVC1–4 than in DL, and that of total aldehydes from approximately 4-fold higher in DLVC2 up to ≈18-fold higher in DLAsc2. The ability of ascorbic acid to enhance lipid oxidation under certain conditions has been related to its capacity to reduce transition metal ions, which would in turn promote the formation of reactive free radicals that would favor lipid oxidation [
21]. In this regard, it should be noticed that the presence of traces of metal ions in the reaction medium cannot be discarded, since previous studies have already reported their presence in edible vegetable oils, especially in the case of such virgin oils as the one employed in this study, as well as in porcine enzymes and extracts employed for preparing digestive juices [
43,
44]. Although the prooxidant behavior of ascorbic acid has been reported to be of relevance when this compound is present at low concentrations [
45], the results of this study suggest that increased lipid oxidation also takes place in the presence of high concentrations of ascorbic acid in relation to that of lipids (170–2500 mg ascorbic acid/salt plus 0.5 g oil submitted to in vitro digestion).
It is also worth noticing that, whereas similar effects on conjugated dienes generation were observed for all doses of ascorbic acid, variations in the levels of aldehydes were found among digests, those coming from the mixtures of linseed oil with pure ascorbic acid (DLAsc1–2) showing the highest concentrations. However, no relationship was found between the amount of ascorbic acid and the level of oxidation products in the digests. As an example, despite a great difference between the amount of the ascorbic acid-based supplement VC1 added to the samples submitted to digestion (2500 mg) and that contained in VC4 (500 mg), the generation of oxidation products was of similar order in their corresponding digests (see DLVC1 and DLVC4 samples in
Table 3).
The results obtained with the other two supplements based on ascorbic acid/salt, but also containing citric acid, carotenes and other components (VC5 and VC6), were completely different. It can be observed in DLVC5–6 spectra in
Figure 1 that signal
b of
Z,
E-CD-OOH is absent. In fact, the proton signals detected in DLVC5–6 spectra are the same ones detected in the starting oil spectrum and show similar intensities (see signals
a and
h in L spectrum). In contrast with that previously observed, the small advance of oxidation occurring during linseed oil digestion is avoided in the presence of VC5–6 supplements (see
Figure 1). Quantitative data reported in
Table 3 confirm these observations. As detailed in
Table 1 of
Section 2, VC5 and VC6 supplements are mixtures of ascorbic acid with citric acid and other components, such as carotenes, neohesperidin DC or orange juice concentrate, which could contribute to oxidation inhibition [
46,
47]. In this sense, citric acid is known to effectively delay lipid oxidation processes by chelating metal ions that could be involved in the mechanism of ascorbic acid to provoke oxidation. Moreover, Osborn-Barnes and Akoh [
48] reported increased efficacy of citric acid in slowing down the generation of hydroperoxides and secondary oxidation products at acidic pH, such as that existing in the digestive tract.
It is noteworthy that similarly to that commented for DLVC5–6, in the linseed oil samples digested together with ascorbyl palmitate (DLAP, DLVC7), oxidation of linseed oil during digestion did not occur either, even though
E-2-alkenals were generated in very small amounts (see
Figure 1 and
Table 3). These findings related to the effect of the presence of ascorbyl palmitate on the oxidation extent of linseed oil during digestion greatly contrast with those observed for ascorbic acid, both pure (DLAsc1–2) and in VC1–4 supplements (DLVC1–4). When trying to explain these differences, the polar paradox theory arises, even though factors other than polarity are known to influence prooxidant and antioxidant behaviors [
49]. In contrast to ascorbic acid, which is highly soluble in water, the amphiphilic nature of ascorbyl palmitate would make these molecules locate at the interface of the lipid droplet, inside of which linseed oil glycerides would be found to be isolated from the aqueous media, where hydrophilic prooxidant species (such as metal ions or small reactive oxygen species) can be found. When ascorbyl palmitate is present at high concentrations, the contact between linseed oil polyunsaturated acyl chains and pro-oxidant species would be significantly hampered and would thus protect against oxidative degradation. Indeed, differences between ascorbic acid and ascorbyl palmitate regarding oxidation were observed in bulk oil by Frankel et al. [
50], in such a way that ascorbic acid showed an antioxidant behavior, whilst ascorbyl palmitate acted as a prooxidant; however, this trend was inverted when studying oil/water emulsions, as occurs in this study.
3.1.2. Effect on the Degradation of Polyunsaturated Acyl Groups during Digestion
As it is well known, fatty acyl chains presenting the highest number of double bonds are those showing the highest susceptibility toward oxidation. Thus, the higher the advance of oxidation, the higher degradation could be expected to affect polyunsaturated fatty acyl chains. It is worth reminding that, as indicated in
Section 2.4.2, the only samples considered for studying the changes in the molar percentages of polyunsaturated chains as a result of digestion were DL, DLAsc1–2, DLVC1 and DLVC5. The molar percentages of linolenate and linoleate chains, before and after digestion, are reported in
Table 4.
Linseed oil is especially rich in polyunsaturated acyl groups, the n-3 linolenate chains being the most abundant ones (≈52.0%). After in vitro digestion, the percentages of both linolenic and linoleic chains showed only slightly (
p > 0.05) decreased values (≈51.3% and ≈17.5% in DL, respectively), indicating that they were degraded to a low extent during digestion. This is in accordance with previous studies on digestion of highly polyunsaturated oils [
17,
18,
30,
51].
When linseed oil is digested in the presence of ascorbic acid, clear differences can be observed among digests. Thus,
Table 4 reveals that DLAsc1–2 and DLVC1 samples showed significantly (
p < 0.05) lower molar proportion of linolenic AG + FA than DL (≈48% versus 51% in DL), whereas the molar percentage of linolenic AG + FA in DLVC5 was very similar to that of linseed oil before being subjected to in vitro digestion (L). These findings are in line with the outcomes of the study of the oxidation product profile of the above-referred samples (
Section 3.1.1). This confirms that the presence of other components in addition to ascorbic acid in supplement VC5 seems to inhibit to a great extent the occurrence of oxidation during digestion of linseed oil.
3.1.3. Effect on Gamma-Tocopherol Degradation during Digestion
The occurrence of oxidation during digestion can also lead to a loss of minor components naturally present in linseed oil to which antioxidant properties have been attributed, such as gamma-tocopherol. Taking this into account, the concentration of gamma-tocopherol was estimated in the oil before digestion and in the lipid extracts of the various in vitro digests, by measuring the area of singlet
i at 6.36 ppm in the multisuppressed
1H NMR spectra (see
Figure 1 and
Table 2 for signal assignment).
As can be observed in
Table 5, the concentration of gamma-tocopherol was greatly affected by the in vitro digestion process, exhibiting a value almost 3-fold lower in the digested lipid extract (DL) than in the starting oil (L). The decrease in the concentration of gamma-tocopherol naturally present in linseed and walnut oils during in vitro digestion was also observed in previous works [
18,
30,
51].
When it comes to the digests corresponding to linseed oil digestion with only ascorbic acid (DLAsc1–2 and DLVC1–4), a total degradation of gamma-tocopherol took place during digestion (see the absence of singlet
i in
Figure 1). This could be expected, bearing in mind that these samples showed a higher oxidation level than DL. By contrast, in the presence of complex mixtures containing ascorbic acid/salt and citric acid, among other components, as is the case of VC5–6 supplements, degradation of gamma-tocopherol occurred to a very low extent, to the point that significant differences between the starting oil (L) and samples DLVC5–6 were not found (see
Table 5 and signal
i in
Figure 1). Indeed, the mean concentration of gamma-tocopherol in DLVC5 and DLVC6 lipid extracts (413.0 and 406.4 μmol/mol AG + FA, respectively) was similar to that found in L (422.7 μmol/mol AG + FA). This indicates that, if oxidation reactions took place in these in vitro digests, their extent was not enough to cause a significant degradation of gamma-tocopherol. Similar results were obtained in the case of the samples digested with ascorbyl palmitate (DLAP and DLVC7). The slightly lower values (
p > 0.05) for gamma-tocopherol concentration found in DLAP and DLVC7 (≈337 and 356 μmol/mol AG + FA) than in L could be explained to a certain extent by the increase in the total AG + FA in the digests due to the presence of the palmitate chains, rather than by the involvement of gamma-tocopherol in oxidation reactions.
In summary, the results concerning the analysis of gamma-tocopherol concentrations before and after in vitro digestion correspond with those related to the oxidation product profile of the various linseed oil digests (
Section 3.1.1).
3.1.4. Effect on the Formation of Volatile Oxidation Compounds Detected through SPME-GC/MS
The study and comparison of the profile of volatile components of several in vitro digests confirm the results obtained through
1H NMR on the different oxidation levels of the digests, regarding the generation of different kinds of oxidation compounds and the degradation of polyunsaturated chains and gamma-tocopherol. It must be highlighted that SPME-GC/MS can provide information on the specific nature of the individual volatile and semi-volatile lipid oxidation products present in the digests, which is important considering that some of them have been related to certain degenerative diseases [
41].
Table 6 shows the abundance in the headspace of the different digests of selected volatile compounds known to be derived from linseed oil oxidation (aldehydes and some furan derivatives) from Maillard-type reactions (two alkyl-pyridines) and potentially from the degradation of ascorbic acid (furfural). It can be observed that the number and abundance of studied volatiles were far higher in DLAsc1–2 and DLVC1–4 than in DL, DLVC5–6, DLAP and DLVC7, these latter samples showing, in general, abundances of similar order. Thus, in line with the results provided through
1H NMR study, the oxidation level of linseed oil digests containing only ascorbic acid, added as pure compound (DLAsc1–2) or as supplement (DLVC1–4), was markedly higher than that reached by the rest.
Among the volatiles coming from linseed oil oxidation during digestion, it can be observed that alkadienals showed the highest abundances in digested samples, followed by alkanals and alkenals, which is in accordance with
1H NMR data (see
Table 3). The main aldehydes detected were reactive 2,4-heptadienals [
34,
52,
53]. Moreover, the oxygenated alpha,beta-unsaturated 4,5-epoxy-2-heptenals and 4,5-epoxy-2-decenals, whose potential toxicity has been described [
41,
52], were only found in DLAsc1–2 and DLVC1–4. As expected, considering linseed oil composition, the main furan derivatives detected in the digest headspaces were 2-pentyl-furan and 2-ethyl-furan [
53]. The detection in DLAsc1–2 and DLVC1–4 of linolenic-derived alkenyl derivatives 2-(2-propenyl)- and 2-(2-pentenyl)-furan [
54], and of two lactones 5-ethyl-2(5
H)- and 5-pentyl-2(5
H)-furanone, is noteworthy.
In addition to lipid oxidation, it is known that during in vitro digestion Maillard-type reactions can also take place. The highest abundances of volatiles coming from these reactions are expected to be present in the digests showing the highest oxidation level due to the higher amounts of carbonyl compounds derived from lipid oxidation. Indeed, in the headspace of DLAsc1–2 and DLVC1–4 samples, significantly increased concentrations of 3-ethyl-pyridine were found, together with 2-ethyl-pyridine, the latter not being detected in the rest of digests. Both compounds are known to be derived from reactions between 2,4-heptadienals and amino compounds [
55].
Although Maillard-type reactions can also give rise to furfural, this volatile is considered to be the main product of ascorbic acid degradation [
56,
57]. Finholt et al. [
57] even reported that under acidic and anaerobic conditions (such as those that could exist during the gastric step of the in vitro procedure), one furfural molecule appears to be formed from one ascorbic acid molecule. The extent of ascorbic acid degradation is expected to be in line with the extent of lipid oxidation. This is evidenced by the detection of significantly (
p < 0.05) higher abundances of furfural in the headspace of DLVC1–3 samples in comparison with the others. It must be noted that the former are the digests of supplements that contain the highest amount of ascorbic acid (2500, 1500 and 1000 mg, respectively) and also show increased oxidation level.





{kind=link}
{kind=link}
