
Development of Selection Indices for Improvement of Seed Yield and Lipid Composition in Bambara Groundnut (Vigna subterranea (L.) Verdc.)

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Composition Dataset
2.1.1. Sampling of Lines
2.1.2. Compositional Analysis Method
2.2. Selection Indices Dataset
2.3. Statistical Analysis
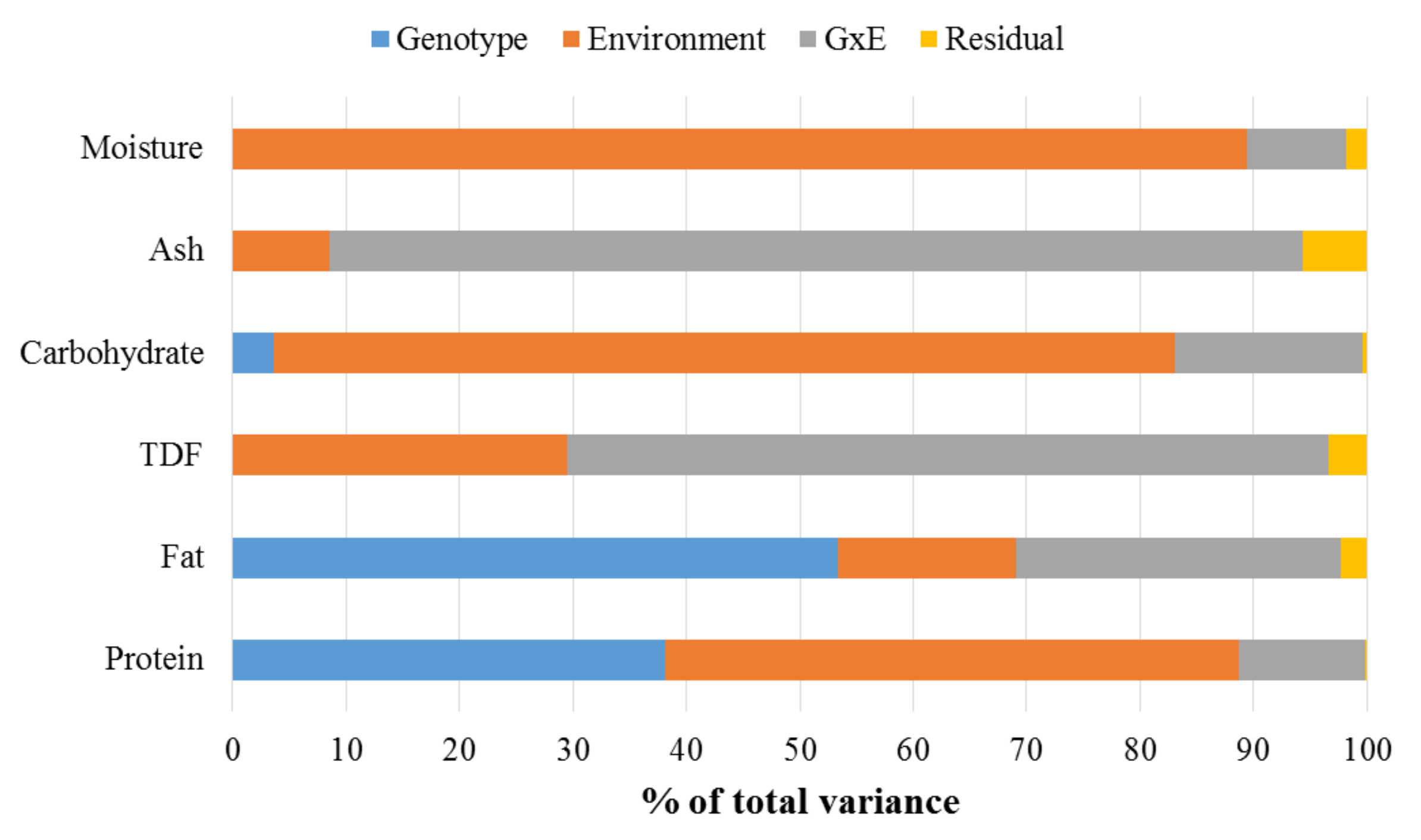
2.4. Heritability Analysis
2.5. Selection Index and Scenario Testing
3. Results
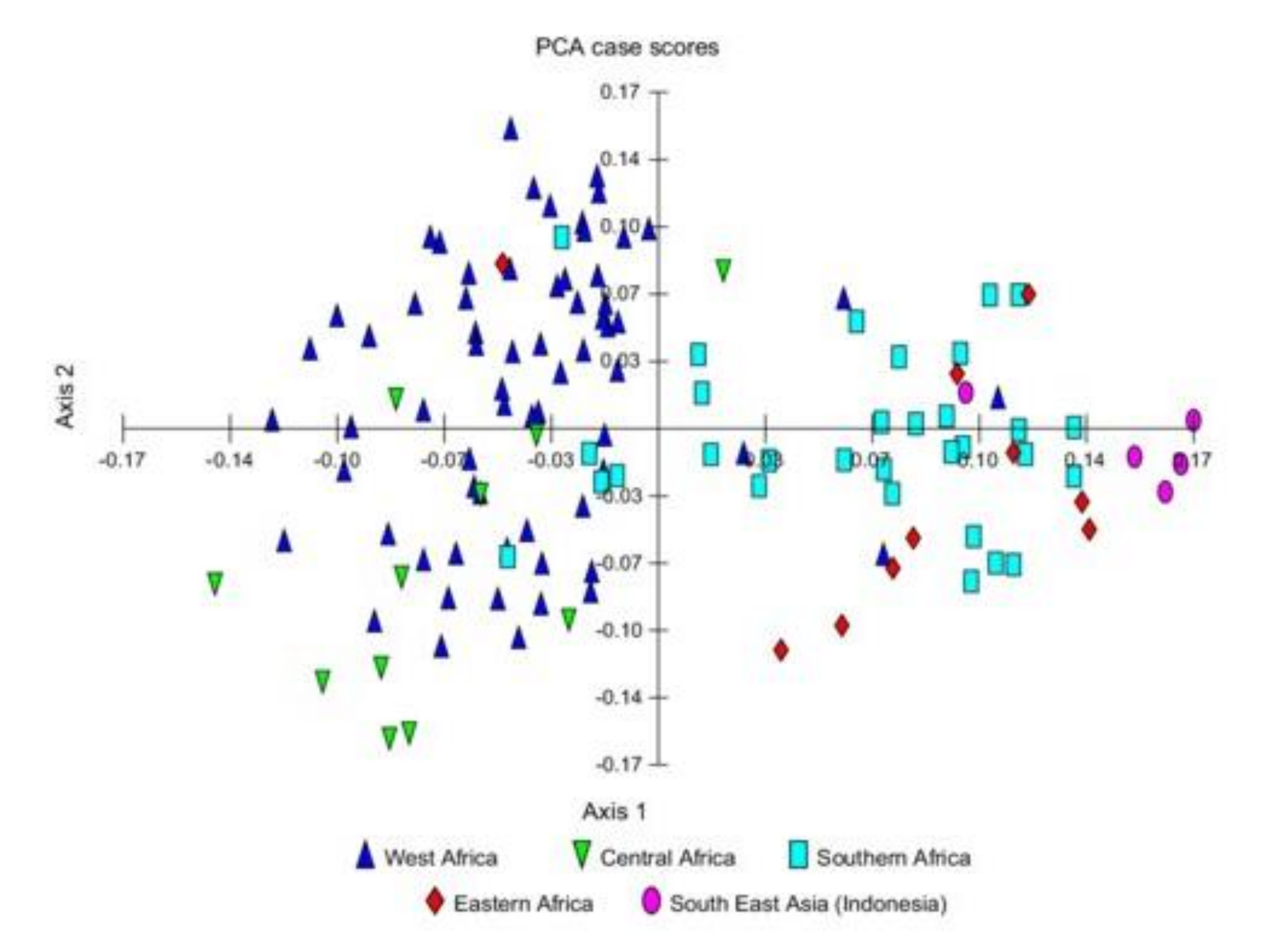
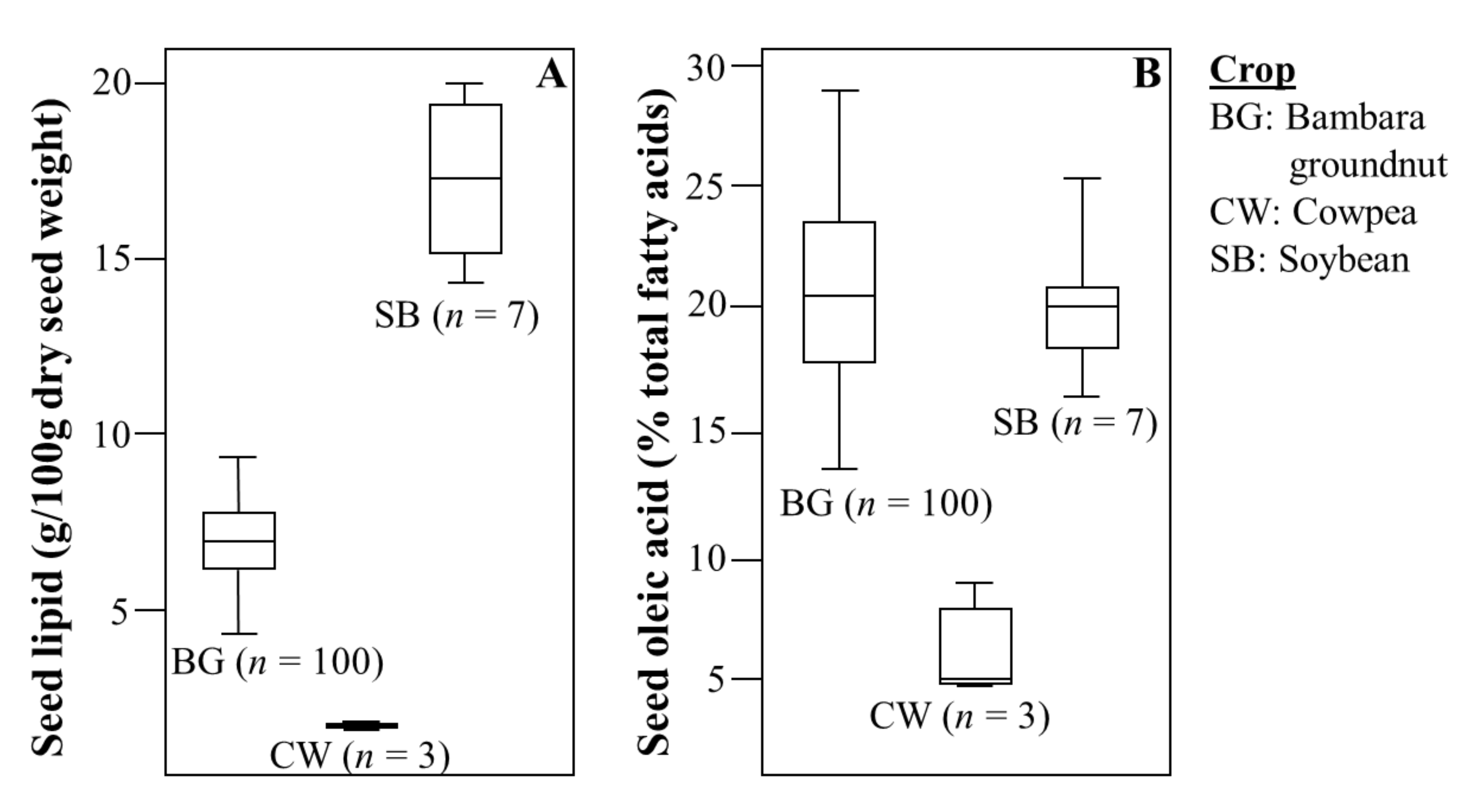
3.1. Compositional Analysis
3.2. Development of Selection Indices
4. Discussion
4.1. Compositional Analysis
4.2. Development of Selection Indices
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Céron-Rojas, J.J.; Crossa, J. (Eds.) The Linear Phenotypic Selction Index Theory. In Linear Selection Indices in Modern Plant Breeding; Springer International Publishing: Cham, Switzerland, 2018; pp. 15–42. [Google Scholar]
- Acheampong, P.P.; Owusu, V.; Nurah, G. How does Farmer Preference matter in Crop variety Adoption? The case of Improved Cassava varieties’ Adoption in Ghana. Open Agric. 2018, 3, 466. [Google Scholar] [CrossRef]
- Christinck, A.; Weltzien, E. Plant breeding for nutrition-sensitive agriculture: An appraisal of developments in plant breeding. Food Secur. 2013, 5, 693–707. [Google Scholar] [CrossRef] [Green Version]
- Gamboa, C.; Broeck, G.V.D.; Maertens, M. Smallholders’ Preferences for Improved Quinoa Varieties in the Peruvian Andes. Sustainability 2018, 10, 3735. [Google Scholar] [CrossRef] [Green Version]
- Elliot, P.; Kingwell, R.; Carter, C.; Yamamoto, M.; White, P. Western Australia’s Noodle Wheat Industry. Current Status and Future Challenges; Australian Export Grains Innovation Centre: Sidney, Australia, 2015. [Google Scholar]
- Sands, D.C.; Morris, C.E.; Dratz, E.A.; Pilgeram, A. Elevating optimal human nutrition to a central goal of plant breeding and production of plant-based foods. Plant Sci. 2009, 177, 377–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burchi, F.; Fanzo, J.; Frison, E. The Role of Food and Nutrition System Approaches in Tackling Hidden Hunger. Int. J. Environ. Res. Public Health 2011, 8, 358–373. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D.; Borelli, T.; Beltrame, D.; Oliveira, C.; Coradin, L.; Wasike, V.; Wasilwa, L.; Mwai, J.; Manjella, A.; Samarasinghe, G.; et al. The potential of neglected and underutilized species for improving diets and nutrition. Planta 2019, 250, 709–729. [Google Scholar] [CrossRef]
- van Der Merwe, J.D.; Cloete, P.C.; van Der Hoeven, M. Promoting food security through indigenous and traditional food crops. Agroecol. Sustain. Food Syst. 2016, 40, 830–847. [Google Scholar] [CrossRef]
- Mabhaudhi, T.; Chimonyo, V.G.P.; Hlahla, S.; Massawe, F.; Mayes, S.; Nhamo, L.; Modi, A.T. Prospects of orphan crops in climate change. Planta 2019, 250, 695–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khoury, C.K.; Bjorkman, A.D.; Dempewolf, H.; Ramirez-Villegas, J.; Guarino, L.; Jarvis, A.; Rieseberg, L.H.; Struik, P.C. Increasing homogeneity in global food supplies and the implications for food security. Proc. Natl. Acad. Sci. USA 2014, 111, 4001. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.N.A.; Sesay, A.; Karikari, S.K.; Massawe, F.J.; Aguilar-Manjarrez, J.; Bannayan, M.; Hampson, K.J. Assesing the Potential of An Underutilised Crop—A Case Study Using Bambara Groundnut. Exp. Agric. 2001, 37, 433–472. [Google Scholar]
- Mabhaudhi, T.; Chimonyo, V.G.P.; Chibarabada, T.P.; Modi, A.T. Developing a Roadmap for Improving Neglected and Underutilized Crops: A Case Study of South Africa. Front. Plant Sci. 2017, 8, 2143. [Google Scholar] [CrossRef] [PubMed]
- Sogbohossou, E.O.D.; Achigan-Dako, E.G.; Maundu, P.; Solberg, S.; Deguenon, E.M.S.; Mumm, R.H.; Hale, I.; Van Deynze, A.; Schranz, M.E. A roadmap for breeding orphan leafy vegetable species: A case study of Gynandropsis gynandra (Cleomaceae). Hortic. Res. 2018, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Mabhaudhi, T.; O’Reilly, P.; Walker, S.; Mwale, S. Opportunities for Underutilised Crops in Southern Africa’s Post–2015 Development Agenda. Sustainability 2016, 8, 302. [Google Scholar] [CrossRef] [Green Version]
- Benedict, O.; Uguru, M.; Aruah, C. Pollen behaviour and fertilization impairmtent in Bambara groundnut (Vigna subterranea (L.) Verdc.). J. Plant Breed. Crop. Sci. 2010, 2, 12–23. [Google Scholar]
- Ho, W.K.; Chai, H.H.; Kendabie, P.; Ahmad, N.S.; Jani, J.; Massawe, F.; Kilian, A.; Mayes, S. Integrating genetic maps in bambara groundnut [Vigna subterranea (L) Verdc.] and their syntenic relationships among closely related legumes. BMC Genom. 2017, 18, 192. [Google Scholar] [CrossRef] [Green Version]
- Mayes, S.; Ho, W.K.; Chai, H.H.; Song, B.; Chang, Y.; Massawe, F. Bambara Groundnut (Vigna Subterranea (L) Verdc)—A Climate Smart Crop for Food and Nutrition Security. In Genomic Designing of Climate-Smart Pulse Crops; Kole, C., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 397–424. [Google Scholar]
- Tsusaka, T.; Madzonga, O.; Gondwe, L.; Harry, M.; Twanje, G.; Spieldoch, A.; Mwakasungula, B.; Meier, W.; Chiwawula, A.; Cardwell, V.; et al. New Frontiers in Legume Research. Case Studies of Ten Years of Research in Malawi; The McKnight Foundation Collaborative Crop Research program (CCRP): Minneapolis, MN, USA, 2016. [Google Scholar]
- Mayes, S.; Ho, W.K.; Chai, H.H.; Gao, X.; Kundy, A.C.; Mateva, K.I.; Zahrulakmal, M.; Hahiree, M.K.I.M.; Kendabie, P.; Licea, L.C.S.; et al. Bambara groundnut: An exemplar underutilised legume for resilience under climate change. Planta 2019, 250, 803–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nono, C.T.; Gouertoumbo, W.; Wakem, G.; Foh, T.; Youmbi, E.; Ntsefong, G.N. Origin and Ecology of Bambara Groundnut (Vigna subterranea (L.) Verdc: A Review. J. Ecol. Nat. Resour. 2018, 2, 1–10. [Google Scholar]
- Ali, S.N.A.; Aguilar-Manjerrez, J.; Bannayan-Avval, M. A Global Mapping System for Bambara Groundnut Production; Food and Agriculture Organisation: Rome, Italy, 2001; pp. 1–47. [Google Scholar]
- Bamshaiye, O.M.; Adegbola, J.A.; Bamishaiye, E.I. Bambara groundnut: An Under-Utilized Nut in Africa. Adv. Agric. Biotechnol. 2011, 1, 60–72. [Google Scholar]
- Chibarabada, T.P.; Modi, A.T.; Mabhaudhi, T. Seed quality characteristics of a bambara groundnut (Vigna subterranea L.) landrace differing in seed coat colour. S. Afr. J. Plant Soil 2014, 31, 219–226. [Google Scholar] [CrossRef]
- Hillocks, R.J.; Bennett, C.; Mponda, O.M. Bambara Nut: A review of utilisation, Market potential and Crop Improvement. Afr. Crop. Sci. J. 2012, 20, 1–6. [Google Scholar]
- Mabhaudhi, T.; Modi, A.T.; Beletse, Y.G. Growth, phenological and yield responses of a bambara groundnut (Vigna subterranea L. Verdc) landrace to imposed water stress: II. Rain shelter conditions. Water SA 2013, 39, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Ho, W.K.; Muchugi, A.; Muthemba, S.; Kariba, R.; Mavenkeni, B.O.; Hendre, P.; Song, B.; Van Deynze, A.; Massawe, F.; Mayes, S. Use of microsatellite markers for the assessment of bambara groundnut breeding system and varietal purity before genome sequencing. Genome 2016, 59, 427–431. [Google Scholar] [CrossRef]
- Tan, X.L.; Azam-Ali, S.; Goh, E.V.; Mustafa, M.; Chai, H.H.; Ho, W.K.; Mayes, S.; Mabhaudhi, T.; Azam-Ali, S.; Massawe, F. Bambara Groundnut: An Underutilized Leguminous Crop for Global Food Security and Nutrition. Front. Nutr. 2020, 7, 276. [Google Scholar] [CrossRef]
- Adu-Dapaah, H.; Berchie, J.N.; Amoah, S.; Addo, J.K.; Akuamoah-Boateng, M. Progress in bambara groundnut research in Ghana: Breeding, agronomy and utilization. In Acta Horticulturae; Onus, N., Currie, A., Eds.; International Society for Horticultural Science: Leuven, Belgium, 2016; Volume 1127, pp. 1–8. [Google Scholar]
- Khan, M.M.H.; Rafii, M.Y.; Ramlee, S.I.; Jusoh, M.; Al Mamun, M. Genetic analysis and selection of Bambara groundnut (Vigna subterranea [L.] Verdc.) landraces for high yield revealed by qualitative and quantitative traits. Sci. Rep. 2021, 11, 7597. [Google Scholar] [CrossRef] [PubMed]
- Redjeki, E.; Mayes, S.; Azam-Ali, S. Evaluating the stability and adaptability of Bambara groundnut (Vigna subterranea (L.) Verd.) landraces in different agro-ecologies. Acta Hortic. 2013, 979, 389–400. [Google Scholar] [CrossRef]
- Alake, C.O.; Ayo-Vaughan, M.A.; Ariyo, J.O. Selection criteria for grain yield and stability in bambara groundnut (Vigna subterranean (L) Verdc) landraces. Acta Agric. Scand. Sect. B Soil Plant Sci. 2015, 65, 433–447. [Google Scholar] [CrossRef]
- Kendabie, P.; Jørgensen, S.T.; Massawe, F.; Fernandez, J.; Azam-Ali, S.; Mayes, S. Photoperiod control of yield and sink capacity in Bambara groundnut (Vigna subterranea) genotypes. Food Energy Secur. 2020, 9, e240. [Google Scholar] [CrossRef]
- Onwubiko, N.C.; Uguru, M.I.; Chimdi, G.O. Estimates of Genetic Parameters in Bambara Groundnut {Vigna subterranea (L.) VERDC.}. Plant Breed. Biotechnol. 2019, 7, 295–301. [Google Scholar] [CrossRef]
- Halimi, R.A.; Barkla, B.J.; Mayes, S.; King, G.J. The potential of the underutilized pulse bambara groundnut (Vigna subterranea (L.) Verdc.) for nutritional food security. J. Food Compos. Anal. 2019, 77, 47–59. [Google Scholar] [CrossRef]
- Alake, C.O. Genetic variability, gains from selection and genetic correlations for pod yield and nutritional traits in African landraces of bambara groundnut. Biol. Agric. Hortic. 2018, 34, 71–87. [Google Scholar] [CrossRef]
- Jost, E.; Ribeiro, N.D.; Maziero, S.M.; Possobom, M.T.D.F.; Rosa, D.P.; Domingues, L.D.S. Comparison among direct, indirect and index selections on agronomic traits and nutritional quality traits in common bean. J. Sci. Food Agric. 2013, 93, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, B.; Kennedy, B.W. Index versus tandem selection after repeated generations of selection. Theor. Appl. Genet. 1993, 85, 706–712. [Google Scholar] [CrossRef]
- Smith, H.F. A Discriminant Function For Plant Selection. Ann. Eugen. 1936, 7, 240–250. [Google Scholar] [CrossRef]
- Rajamani, S.; Sreekanth, M.; Naik, V.S.; Ratnam, M. Selection indices for yield attributing characters improvement in pigeon pea (Cajanus cajan L. Millspugh). Int. J. Life Sci. 2016, 2, 127–129. [Google Scholar]
- Ranjani, M.; Vanniarajan, C.; Kumar, C.S.; Saxena, R.; Sudhagar, R.; Hingane, A. Genetic variability and association studies for yield and its attributes in super-early pigeonpea (Cajanus cajan (L.) Millsp.) Genotypes. Electron J. Plant Breed. 2018, 9, 682–691. [Google Scholar] [CrossRef]
- Charrondiere, U.R.; Chevassus-Agnes, S.; Marroni, S.; Burlingame, B. Impact of different macronutrient definitions and energy conversion factors on energy supply estimations. J. Food Compos. Anal. 2004, 17, 339–360. [Google Scholar] [CrossRef]
- Zou, M.L.; Moughan, P.J.; Awati, A.; Livesey, G. Accuracy of the Atwater factors and related food energy conversion factors with low-fat, high-fiber diets when energy intake is reduced spontaneously. Am. J. Clin. Nutr. 2007, 86, 1649–1656. [Google Scholar] [CrossRef]
- FAO. Food Energy—Methods of Analysis and Conversion Factors. Report of a Technical Workshop, Rome, 3–6 December 2002; FAO: Rome, Italy, 2003. [Google Scholar]
- Medic, J.; Atkinson, C.; Hurburgh Jr., C. R. Current Knowledge in Soybean Composition. J. Am. Oil Chem. Soc. 2014, 91, 363–384. [Google Scholar] [CrossRef]
- Dyer, J.M.; Stymne, S.; Green, A.G.; Carlsson, A.S. High-value oils from plants. Plant J. 2008, 54, 640–655. [Google Scholar] [CrossRef] [PubMed]
- Redondo-Cuenca, A.; Villanueva-Suarez, M.J.; Rodriguez-Sevilla, M.D.; Mateos-Aparicio, I. Chemical composition and dietary fibre of yellow and green commercial soybeans (Glycine max). Food Chem. 2006, 101, 1216–1222. [Google Scholar] [CrossRef]
- Ijarotimi, O.S.; Esho, T.R. Comparison of nutritional composition and anti-nutrient status of fermented, germinated and roasted bambara groundnut seeds (Vigna subterranea). Br. Food J. 2009, 111, 376–386. [Google Scholar] [CrossRef]
- FAO/INFOODS. Guidelines for Converting Units, Denominators and Expressions; FAO: Rome, Italy, 2012. [Google Scholar]
- IPGRI; International Institute of Tropical (IITA). BAMNET, Descriptors for Bambara Groundnut (Vigna subterranea), International Plant Genetic Resources Institute, Rome, Italy; International Institute of Tropical Agriculture: Ibadan, Nigeria; Rome, Italy, 2000. [Google Scholar]
- Andrés-Hernández, L.; King, G.J. Compositional Dietary Nutrition Ontology (CDNO). Available online: https://obofoundry.org/ontology/cdno.html (accessed on 25 February 2021).
- Ontology, C. Bambara Groundnut Ontology. Available online: https://cropontology.org/ontology/CO_366 (accessed on 28 September 2021).
- Kirch, W. Pearson’s Correlation Coefficient. In Encyclopedia of Public Health; Kirch, W., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 1090–1091. [Google Scholar]
- Lawrence, M.J.; Marshall, D.F.; Davies, P. Genetics of genetic conservation. I. Sample size when collecting germplasm. Euphytica 1995, 84, 89–99. [Google Scholar] [CrossRef]
- Lawrence, M.J.; Marshall, D.F.; Davies, P. Genetics of genetic conservation. II. Sample size when collecting seed of cross-pollinating species and the information that can be obtained from the evaluation of material held in gene banks. Euphytica 1995, 84, 101–107. [Google Scholar] [CrossRef]
- Molosiwa, O.O.; Aliyu, S.; Stadler, F.; Mayes, K.; Massawe, F.; Kilian, A.; Mayes, S. SSR marker development, genetic diversity and population structure analysis of Bambara groundnut [Vigna subterranea (L.) Verdc.] landraces. Genet. Resour. Crop. Evol. 2015, 62, 1225–1243. [Google Scholar] [CrossRef]
- Carrera, C.S.; Reynoso, C.M.; Funes, G.J.; Martínez, M.J.; Dardanelli, J.; Resnik, S.L. Amino acid composition of soybean seeds as affected by climatic variables. Pesqui. Agropecu. Bras. 2011, 46, 1579–1587. [Google Scholar] [CrossRef]
- Qin, P.; Song, W.; Yang, X.; Sun, S.; Zhou, X.; Yang, R.; Li, N.; Hou, W.; Wu, C.; Han, T.; et al. Regional distribution of protein and oil compositions of soybean cultivars in China. Crop Sci. 2014, 54, 1139–1146. [Google Scholar] [CrossRef]
- Assefa, Y.; Purcell, L.C.; Salmeron, M.; Naeve, S.; Casteel, S.N.; Kovács, P.; Archontoulis, S.; Licht, M.; Below, F.; Kandel, H.; et al. Assessing Variation in US Soybean Seed Composition (Protein and Oil). Front. Plant Sci. 2019, 10, 298. [Google Scholar] [CrossRef]
- Oluwatosin, O.B. Genetic and environmental variability in starch, fatty acids and mineral nutrients composition in cowpea (Vigna unguiculata (L.) Walp. J. Sci. Food Agric. 1998, 78, 1–11. [Google Scholar] [CrossRef]
- Boukar, O.; Fatokun, C.A.; Huynh, B.-L.; Roberts, P.A.; Close, T.J. Genomic Tools in Cowpea Breeding Programs: Status and Perspectives. Front. Plant Sci. 2016, 7, 757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, D.; Rocha, M.D.M.; Silva, K.; Sá, F.; Lima, L.; de Resende, M. Genotypic gain with simultaneous selection of production, nutrition, and culinary traits in cowpea crosses and backcrosses using mixed models. Embrapa Meio-Norte-Artigo Periódico Indexado (ALICE) 2017, 16, 1–10. [Google Scholar] [CrossRef]
- Mudryj, A.N.; Yu, N.; Aukema, H.M. Nutritional and health benefits of pulses. Appl. Physiol. Nutr. Metab. 2014, 39, 1197. [Google Scholar] [CrossRef] [PubMed]
- Asif, M.; Rooney, L.W.; Ali, R.; Riaz, M.N. Application and opportunities of pulses in food system: A review. Crit. Rev. Food Sci. Nutr. 2013, 53, 1168–1179. [Google Scholar] [CrossRef] [PubMed]
- Singh, N. Pulses: An overview. J. Food Sci. Technol. 2017, 54, 853–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patto, M.C.V.; Amarowicz, R.; Aryee, A.N.A.; Boye, J.I.; Chung, H.-J.; Martín-Cabrejas, M.A.; Domoney, C. Achievements and Challenges in Improving the Nutritional Quality of Food Legumes. Crit. Rev. Plant Sci. 2015, 34, 105–143. [Google Scholar] [CrossRef]
- Lee, S.; Van, K.; Sung, M.; Nelson, R.; LaMantia, J.; McHale, L.K.; Mian, M.A.R. Genome-wide association study of seed protein, oil and amino acid contents in soybean from maturity groups I to IV. Theor. Appl. Genet. 2019, 132, 1639–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachleda, N.; Grey, T.; Li, Z. Effects of high oleic acid soybean on seed yield, protein and oil contents, and seed germination revealed by near-isogeneic lines. Plant Breed. 2017, 136, 539–547. [Google Scholar] [CrossRef]
- Kim, H.-J.; Ha, B.-K.; Ha, K.-S.; Chae, J.-H.; Park, J.-H.; Kim, M.-S.; Asekova, S.; Shannon, J.G.; Son, C.-K.; Lee, J.-D. Comparison of a high oleic acid soybean line to cultivated cultivars for seed yield, protein and oil concentrations. Euphytica 2015, 201, 285–292. [Google Scholar] [CrossRef]
- Nawade, B.; Mishra, G.P.; Radhakrishnan, T.; Dodia, S.M.; Ahmad, S.; Kumar, A.; Kumar, A.; Kundu, R. High oleic peanut breeding: Achievements, perspectives, and prospects. Trends Food Sci. Technol. 2018, 78, 107–119. [Google Scholar] [CrossRef]
- Bielecka, M.; Kaminski, F.; Adams, I.; Poulson, H.; Sloan, R.; Li, Y.; Larson, T.R.; Winzer, T.; Graham, I.A. Targeted mutation of Δ12 and Δ15 desaturase genes in hemp produce major alterations in seed fatty acid composition including a high oleic hemp oil. Plant Biotechnol. J. 2014, 12, 613–623. [Google Scholar] [CrossRef] [Green Version]
- Bilyeu, K.; Škrabišová, M.; Allen, D.; Rajcan, I.; Palmquist, D.E.; Gillen, A.; Mian, R.; Jo, H. The Interaction of the Soybean Seed High Oleic Acid Oil Trait With Other Fatty Acid Modifications. J. Am. Oil Chem. Soc. 2018, 95, 39–49. [Google Scholar] [CrossRef]
- Maestri, D.M.; Labuckas, D.O.; Guzmán, C.A.; Giorda, L.M. Correlation between seed size, protein and oil contents, and fatty acid composition in soybean genotypes. Grasas Aceites 1998, 49, 450–453. [Google Scholar] [CrossRef] [Green Version]
- Yol, E.; Ustun, R.; Golukcu, M.; Uzun, B. Oil Content, Oil Yield and Fatty Acid Profile of Groundnut Germplasm in Mediterranean Climates. J. Am. Oil Chem. Soc. 2017, 94, 787–804. [Google Scholar] [CrossRef]
- Samad, M.; Sarker, N.; Deb, A.J.T.P.R. Study on relationship and selection index in chickpea. Trop. Plant Res. 2014, 1, 27–35. [Google Scholar]
- Bharadwaj, C.; Patil, B.; Madan, P.; Manu, B.; Tapan, K.; Neeraj, K.; Philanim, S.; Soren, K.; Jain, P.; Supriya, S. Breeding Chickpea for Water Limited Environments: Selection Indices and Strategies. In Proceedings of the InterDrought-V, Hyderabad, India, 21–25 February 2017. [Google Scholar]
- Sellami, M.H.; Lavini, A.; Pulvento, C. Phenotypic and Quality Traits of Chickpea Genotypes under Rainfed Conditions in South Italy. Agronomy 2021, 11, 962. [Google Scholar] [CrossRef]
- Diaz, S.; Ariza-Suarez, D.; Ramdeen, R.; Aparicio, J.; Arunachalam, N.; Hernandez, C.; Diaz, H.; Ruiz, H.; Piepho, H.-P.; Raatz, B. Genetic Architecture and Genomic Prediction of Cooking Time in Common Bean (Phaseolus vulgaris L.). Front. Plant Sci. 2021, 11, 2257. [Google Scholar] [CrossRef] [PubMed]
- Lima, D.C.; Abreu, A.F.B.; Ferreira, R.A.D.C.; Ramalho, M.A.P. Breeding common bean populations for traits using selection index. Sci. Agric. 2015, 72, 132–137. [Google Scholar] [CrossRef] [Green Version]
- Batista, L.; Anjos, R.; Poersch, N.; Nalin, R.; Carneiro, P.; Carneiro, J.; de Resende, M.D. Multigeneration Index in the Selection of Common Bean Inbred Families. Crop Sci. 2017, 57, 2354–2360. [Google Scholar] [CrossRef]
- Health, A.M. Nutrient Reference Values for Australia and New Zealand; National Health and Medical Research Council: Canberra, Australia, 2014. [Google Scholar]
- Luo, H.; Pandey, M.K.; Khan, A.W.; Guo, J.; Wu, B.; Cai, Y.; Huang, L.; Zhou, X.; Chen, Y.; Chen, W.; et al. Discovery of genomic regions and candidate genes controlling shelling percentage using QTL-seq approach in cultivated peanut (Arachis hypogaea L.). Plant Biotechnol. J. 2019, 17, 1248–1260. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Ren, X.; Chen, Y.; Huang, L.; Zhou, X.; Huang, J.; Froenicke, L.; Yu, J.; Guo, B.; Liao, B.J.P.G.R. Phenotypic evaluation of the Chinese mini-mini core collection of peanut (Arachis hypogaea L.) and assessment for resistance to bacterial wilt disease caused by Ralstonia solanacearum. Plant Genet. Res. 2013, 11, 77–83. [Google Scholar] [CrossRef]
- Yol, E.; Furat, S.; Upadhyaya, H.D.; Uzun, B.J.J. Characterization of groundnut (Arachis hypogaea L.) collection using quantitative and qualitative traits in the Mediterranean Basin. J. Integr. Agric. 2018, 17, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Bamba, A.S.A.; Kundy, A.C.; Mateva, K.I.; Chai, H.H.; Ho, W.K.; Musa, M.; Mayes, S.; Massawe, F.J.A. Variation of phenotypic traits in twelve bambara groundnut (Vigna subterranea (L.) Verdc.) genotypes and two F2 bi-parental segregating populations. Agronomy 2020, 10, 1451. [Google Scholar] [CrossRef]
- Abu, H.; Saaka, S.; Buah, J. Characterization of Bambara Groundnut Landraces and Their Evaluation by Farmers in the Upper West Region of Ghana. J. Dev. Sustain. Agric. 2011, 6, 64–74. [Google Scholar]
- Mubaiwa, J.; Fogliano, V.; Chidewe, C.; Bakker, E.J.; Linnemann, A.R. Utilization of bambara groundnut (Vigna subterranea (L.) Verdc.) for sustainable food and nutrition security in semi-arid regions of Zimbabwe. PLoS ONE 2018, 13, e0204817. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, M.; Shimelis, H.; Laing, M. Farmers’ preference of bambara groundnut varieties and their breeding priority in Kano State of Nigeria. Techno Sci. Afr. 2016, 13, 1–10. [Google Scholar]
- Akpalu, M.; Atubilla, I.; Oppong-Sekyere, D. Accessing the level of cultivation and utilization of Bambara groundnut (Vigna subterranea (L.)) in the Sumbrungu community of the Upper East Region. Int. J. Plant Anim. Environ. Sci. 2013, 3, 68–75. [Google Scholar]
- Redjeki, E.S.; Ho, W.K.; Shah, N.; Molosiwa, O.O.; Ardiarini, N.R.; Mayes, S. Understanding the genetic relationships between Indonesian bambara groundnut landraces and investigating their origins. Genome 2020, 63, 319–327. [Google Scholar] [CrossRef]
- Saptadi, D.; Ardiarini, N.R.; Waluyo, B.; Kuswanto, K. Agronomic characteristic and variability of bambara groundnut [Vigna subterranea (L.) Verdc. (Fabaceae)] promising lines from Indonesia. In Transactions of Persatuan Genetik Malaysia; Persatuan Genetik Malaysia: Kuala Pilah, Malaysia, 2016; Volume 3, pp. 56–60. [Google Scholar]
- Brough, S.H.; Azam-Ali, S.N.; Taylor, A.J. The potential of bambara groundnut (Vigna subterranea) in vegetable milk production and basic protein functionality systems. Food Chem. 1993, 47, 277–283. [Google Scholar] [CrossRef]
- Ibrahim, A.R.; Dansi, A.; Salifou, M.; Ousmane, A.; Alzouma, A.; Alou, W. Farmers’ practices, utilization, conservation and marketing of Bambara groundnut (Vigna subterranea (L.) Verdc.) in Dosso Region, Western Niger. Genet. Resour. Crop Evol. 2018, 65, 1907–1914. [Google Scholar] [CrossRef] [Green Version]
- Unigwe, A.E.; Doria, E.; Adebola, P.; Gerrano, A.S.; Pillay, M. Anti-nutrient analysis of 30 Bambara groundnut (Vigna subterranea) accessions in South Africa. J. Crop. Improv. 2017, 32, 208–224. [Google Scholar] [CrossRef]
- Mubaiwa, J.; Fogliano, V.; Chidewe, C.; Linnemann, A.R. Hard-to-cook phenomenon in bambara groundnut (Vigna subterranea (L.) Verdc.) processing: Options to improve its role in providing food security. Food Rev. Int. 2016, 33, 167–194. [Google Scholar] [CrossRef] [Green Version]
- Fehr, W.R. Breeding for Modified Fatty Acid Composition in Soybean. Crop Sci. 2007, 47, S72–S87. [Google Scholar] [CrossRef]
- Wilson, R.; Burton, J.; Pantalone, V.; Dewey, R. New gene combinations governing saturated and unsaturated fatty acid composition in soybean. In Lipid Biotechnology; Gardner, H., Kuo, T.M., Eds.; CRC Press: Boca Raton, FL, USA, 2002; pp. 95–114. [Google Scholar]
- Burton, J.W.; Miranda, L. Soybean Improvement: Achievements and Challenges; Institute of Field and Vegetable Crops: Novi Sad, Serbia, 2013; Volume 50, pp. 44–51. [Google Scholar]
- Sweeney, D.W.; Carrero-Colón, M.; Hudson, K.A. Characterization of New Allelic Combinations for High-Oleic Soybeans. Crop Sci. 2017, 57, 611–616. [Google Scholar] [CrossRef]
- Pham, A.-T.; Shannon, J.G.; Bilyeu, K.D. Combinations of mutant FAD2 and FAD3 genes to produce high oleic acid and low linolenic acid soybean oil. Theor. Appl. Genet. 2012, 125, 503–515. [Google Scholar] [CrossRef]
- Antova, G.A.; Stoilova, T.D.; Ivanova, M.M. Proximate and lipid composition of cowpea (Vigna unguiculata L.) cultivated in Bulgaria. J. Food Compos. Anal. 2014, 33, 146–152. [Google Scholar] [CrossRef]
- Zia-Ul-Haq, M.; Ahmad, M.; Iqbal, S. Characteristics of oil from seeds of 4 mungbean [Vigna radiata (L.) Wilczek] cultivars grown in Pakistan. JAOCS J. Am. Oil Chem. Soc. 2008, 85, 851–856. [Google Scholar] [CrossRef]
- Vasconcelos, I.M.; Maia, F.M.M.; Farias, D.F.; Campello, C.C.; Carvalho, A.F.U.; Moreira, R.D.A.; de Oliveira, J.T.A. Protein fractions, amino acid composition and antinutritional constituents of high-yielding cowpea cultivars. J. Food Compos. Anal. 2010, 23, 54–60. [Google Scholar] [CrossRef]
- Nair, R.M.; Yang, R.Y.; Easdown, W.J.; Thavarajah, D.; Thavarajah, P.; Hughes, J.D.A.; Keatinge, J.D.H. Biofortification of mungbean (Vigna radiata) as a whole food to enhance human health. J. Sci. Food Agric. 2013, 93, 1805–1813. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Line Name | Country of Origin | Dataset | Provided by | Growing Season | Growing Location |
|---|---|---|---|---|---|
| 100SB16ANAM-C-A-16-4 | Namibia | SC and SI | CFF | 2016 | Malaysia |
| 104S-1913NAM | Namibia | SC and SI | CFF | 2015 | Malaysia |
| 109BWA1BWA-1 | Botswana | SC and SI | CFF | 2015 | Malaysia |
| 20Acc118CIV-B | Ivory Coast | SC and SI | CFF | 2016 | Malaysia |
| 56Acc89MLI-C | Mali | SC and SI | CFF | 2015 | Malaysia |
| 84ACC696ZMB-2 | Zimbabwe | SC and SI | CFF | 2015 | Malaysia |
| 99SB4-2NAM-A | Namibia | SC and SI | CFF | 2015 | Malaysia |
| 60ACC32NGA-A | Nigeria | SC and SI | CFF | 2015 | Malaysia |
| 1-76ACC390SDN-CA11 | Sudan | SC and SI | CFF | 2016 | Malaysia |
| 91UNISRSWA-B | Swaziland | SC and SI | CFF | 2015 | Malaysia |
| 92AHM968NAM-C | Namibia | SC and SI | CFF | 2015 | Malaysia |
| Ankpa-4 | Nigeria | SC only | UoN | 2016 | UK |
| BC12105 | Indonesia | SC only | CFF | 2015 | Malaysia |
| BC31107 | Indonesia | SC and SI | CFF | 2017 | Malaysia |
| BCGC12107 | Indonesia | SC only | CFF | 2015 | Malaysia |
| BCGC23107 | Indonesia | SC and SI | CFF | 2015 | Malaysia |
| BCL33107 | Indonesia | SC only | CFF | 2017 | Malaysia |
| BH16107 | Indonesia | SC and SI | CFF | 2017 | Malaysia |
| BH17107 | Indonesia | SC only | CFF | 2015 | Malaysia |
| BD | Indonesia | SC and SI | CFF | 2015 | Malaysia |
| Bogor | Indonesia | SC only | CFF | 2017 | Malaysia |
| Burkina * | Burkina Faso | SC and SI | CFF | 2015 | Malaysia |
| Cikur 2.1_CFF_2016 | Indonesia | SC and SI | CFF | 2016 | Malaysia |
| Cikur 2.3 | Indonesia | SC only | CFF | 2015 | Malaysia |
| Cikur 3.3 | Indonesia | SC only | CFF | 2015 | Malaysia |
| Cream Grey Eye-49 | Ghana | SC and SI | CFF | 2016 | Malaysia |
| Cream Light Brown Eye | Ghana | SC and SI | CFF | 2015 | Malaysia |
| Cream with black splash | Ghana | SC only | CFF | 2015 | Malaysia |
| DipC_CFF ** | Botswana | SC and SI | CFF | 2016 | Malaysia |
| DipC_UoN ** | Botswana | SC only | UoN | 2016 | UK |
| DodR_CFF ** | Tanzania | SC and SI | CFF | 2015 | Malaysia |
| DodR_UoN ** | Tanzania | SC only | UoN | 2016 | UK |
| DBB | Ghana | SC and SI | CFF | 2015 | Malaysia |
| Exsokoto * | Nigeria | SC only | CFF | 2016 | Malaysia |
| Exsokoto-39 | Nigeria | SC and SI | CFF | 2017 | Malaysia |
| Exsokoto-177 | Nigeria | SC only | CFF | 2017 | Malaysia |
| Exsokoto-183 | Nigeria | SC only | CFF | 2017 | Malaysia |
| GC11105 | Indonesia | SC only | CFF | 2015 | Malaysia |
| GC32105 | Indonesia | SC only | CFF | 2015 | Malaysia |
| GC35107 | Indonesia | SC only | CFF | 2015 | Malaysia |
| GCL13105 | Indonesia | SC only | CFF | 2015 | Malaysia |
| Getso_UoN | Indonesia | SC only | UoN | 2016 | UK |
| GH17105 | Indonesia | SC only | CFF | 2015 | Malaysia |
| GH21105 | Indonesia | SC only | CFF | 2015 | Malaysia |
| GH36107 | Indonesia | SC only | CFF | 2015 | Malaysia |
| GH37107 | Indonesia | SC and SI | CFF | 2015 | Malaysia |
| GHC36105 | Indonesia | SC and SI | CFF | 2015 | Malaysia |
| Gresik_CFF ** | Indonesia | SC only | CFF | 2016 | Malaysia |
| Gresik_UON ** | Indonesia | SC only | UoN | 2016 | UK |
| GOBRAS2.2 | Indonesia | SC only | CFF | 2015 | Malaysia |
| IITA 686_CFF **/* | Nigeria | SC and SI | CFF | 2016 | Malaysia |
| IITA-686_UoN ** | Nigeria | SC only | UoN | 2016 | UK |
| IPB Bam-1 | Indonesia | SC and SI | CFF | 2015 | Malaysia |
| IPB Bam-2 | Indonesia | SC and SI | CFF | 2015 | Malaysia |
| IPB Bam-5 | Indonesia | SC only | CFF | 2015 | Malaysia |
| IPB Bam-6_CFF2016 | Indonesia | SC and SI | CFF | 2016 | Malaysia |
| IPB Bam-10 | Indonesia | SC only | CFF | 2015 | Malaysia |
| Kaaro | Nigeria | SC only | CFF | 2016 | Malaysia |
| Kaaro-4 * | Nigeria | SC only | CFF | 2017 | Malaysia |
| Kaaro-66 | Nigeria | SC and SI | CFF | 2017 | Malaysia |
| Kaaro-74 * | Nigeria | SC only | CFF | 2017 | Malaysia |
| Kano2 | Nigeria | SC only | UoN | 2016 | UK |
| Kano | Nigeria | SC only | UoN | 2016 | UK |
| LunT | Sierra Leone | SC and SI | CFF | 2017 | Malaysia |
| LBLBE | Ghana | SC and SI | CFF | 2016 | Malaysia |
| Mottled Black_2016 | Ghana | SC and SI | CFF | 2016 | Malaysia |
| Nav4-13 | Ghana | SC and SI | CFF | 2017 | Malaysia |
| Rajap 3.2 | Indonesia | SC and SI | CFF | 2015 | Malaysia |
| S19-3_CFF ** | Namibia | SC and SI | CFF | 2017 | Malaysia |
| S19-3_UoN ** | Namibia | SC only | UoN | 2016 | UK |
| Situraja2 | Indonesia | SC only | CFF | 2016 | Malaysia |
| Songkhla-1 * | Thailand | SC and SI | CFF | 2016 | Malaysia |
| Songkhla-1-38 | Thailand | SC only | CFF | 2017 | Malaysia |
| Songkhla-1-72 | Thailand | SC only | CFF | 2017 | Malaysia |
| Songkhla-1-90 | Thailand | SC only | CFF | 2017 | Malaysia |
| Sukuraja-2 | Indonesia | SC only | CFF | 2015 | Malaysia |
| Thung yang dang | Thailand | SC only | CFF | 2015 | Malaysia |
| Tiga Nicaru | Nigeria | SC only | UoN | 2016 | UK |
| TVSU 738 | Zambia | SC only | CFF | 2015 | Malaysia |
| TVSU 89 | Mali | SC and SI | CFF | 2015 | Malaysia |
| TVSu 880 | Zambia | SC only | AGG | 2017 | Australia |
| TVSu 879 | Zambia | SC only | AGG | 2017 | Australia |
| TVSu 702 | Zambia | SC only | AGG | 2017 | Australia |
| TVSU 334 | Nigeria | SC only | AGG | 2017 | Australia |
| TVSU 922 | Zambia | SC only | AGG | 2017 | Australia |
| TVSU 924 | Zambia | SC only | AGG | 2017 | Australia |
| TVSu 323 | Nigeria | SC only | AGG | 2017 | Australia |
| TVSu 1023 | Zimbabwe | SC only | AGG | 2017 | Australia |
| TVSu 1033 | Zimbabwe | SC only | AGG | 2017 | Australia |
| TVSU 1231 | Nigeria | SC only | AGG | 2017 | Australia |
| TVSu 702-2018 | Zambia | SC only | AGG | 2018 | Australia |
| TVSu 879-2018 | Zambia | SC only | AGG | 2018 | Australia |
| TVSu 880-2018 | Zambia | SC only | AGG | 2018 | Australia |
| Uniswa Red | Swaziland | SC and SI | CFF | 2017 | Malaysia |
| Uniswa Red_AB_CFF ** | Swaziland | SC only | CFF | 2016 | Malaysia |
| Uniswa Red-Red_UoN ** | Swaziland | SC only | UoN | 2016 | UK |
| Uniswa Red-Greeen_UoN | Swaziland | SC only | UoN | 2016 | UK |
| URUG2(1) | Indonesia | SC only | CFF | 2015 | Malaysia |
| Zebra Cream-8 | Ghana | SC only | CFF | 2016 | Malaysia |
| Zebra Cream-10 (KM10) | Ghana | SC and SI | CFF | 2016 | Malaysia |
| Species | Line Name | Provided by | Accession Number | Growing Season | Growing Location |
|---|---|---|---|---|---|
| Soybean | Williams82 | AGG | AGG100180SOYB1 1 | 2017 | Australia |
| Soybean | Stuart | AGG | AGG323309SOYB1 1 | 2017 | Australia |
| Soybean | Essex | AGG | AGG104447SOYB1 1 | 2017 | Australia |
| Soybean | Peking | AGG | AGG65879SOYB4 1 | 2017 | Australia |
| Soybean | Asgrow | TR | N/A | 2017 | Australia |
| Soybean | Richmond | TR | N/A | 2017 | Australia |
| Soybean | Hayman | TR | N/A | 2017 | Australia |
| Cowpea | IT84S-2246-4 | AGG | AGG306477COWP2 1 | 2017 | Australia |
| Cowpea | Han Chui Yen | AGG | AGG306534COWP2 1 | 2017 | Australia |
| Cowpea | 524B | AGG | AGG317984COWP2 1 | 2017 | Australia |
| Trait Name | Method | Controlled Vocabulary Terms | |
|---|---|---|---|
| Crop Dietary Nutritional Ontology 1 | Crop Ontology 2 | ||
| Moisture | AOAC 925.09 | CDNO:0200002 | CO_366:0000185 |
| Ash | AOAC 942.05 | CDNO:0200004 | N/A |
| Lipid | AOAC 948.22 | CDNO:0200068 | CO_366:0000023 |
| Protein | AOAC 992.23 | CDNO:0200040 | CO_366:0000026 |
| Total carbohydrate | N/A | CDNO:0200005 | CO_366:0000309 |
| Fatty acid composition | AOAC996.06 | CDNO:0200465 | N/A |
| Oleic acid | AOAC996.06 | CDNO:0200085 | N/A |
| Lignoceric acid | AOAC996.06 | CDNO:0200081 | N/A |
| Linoleic acid | AOAC996.06 | CDNO:0200102 | N/A |
| Linolenic acid | AOAC996.06 | CDNO:0200097 | N/A |
| Seed number | IPGRI descriptor | N/A | CO_366:0000340 |
| Dry seed weight | IPGRI descriptor | N/A | CO_366:0000337 |
| Dry pod weight | IPGRI descriptor | N/A | CO_366:0000325 |
| Single seed size | IPGRI descriptor | N/A | CO_366:0000328 |
| Shelling percentage | IPGRI descriptor | N/A | CO_366:0000334 |
| Selection Index | Potential End User | Scenarios Tested | Traits Selected for Scenario | Trait Weighting |
|---|---|---|---|---|
| Seed lipid (SI-1) | Nutritionist, food industry, plant breeder | (1a) Oleic acid | (1a) Oleic acid | 3 |
| Omega 3 and 6 fatty acids (SI-2) | Nutritionist, food industry, plant breeder | (2a) Linoleic acid and Linolenic acid | (2a) Linoleic acid Linolenic acid | 3 3 |
| Seed lipid and yield (SI-3) | Plant breeder | (3a) Oleic acid and seed weight (3b) Oleic acid and single seed size (3c) Oleic acid and seed number (3d) Oleic acid, seed number, and seed weight | (3a) Oleic acid Seed weight (3b) Oleic acid Seed size (3c) Oleic acid Seed number (3d) Oleic acid Seed number Seed weight | 3 2 3 2 3 2 3 2 2 |
| Omega 3, 6 fatty acid and yield (SI-4) | Plant breeder | (4a) Linoleic acid, linolenic acid, and seed weight (4b) Linoleic acid, linolenic acid, and single seed size (4c) Linoleic acid, Linolenic acid, and seed number (4d) Linoleic acid, linolenic acid seed number, and seed weight | (4a) Linoleic acid Linolenic acid Seed weight (4b) Linoleic acid Linolenic acid Seed size (4c) Linoleic acid Linolenic acid Seed number (4d) Linoleic acid Linolenic acid Seed number Seed weight | 3 3 2 3 3 2 3 3 2 3 3 2 2 |
| Yield (SI-5) | Farmer | (5a) Seed weight (5b) Seed number (5c) Single seed size (5d) Seed number and seed weight (5e) Seed number, seed weight, and single seed size | (5a) Seed weight (5b) Seed number (5c) Seed size (5d) Seed number Seed weight (5e) Seed number Seed weight Seed size | 2 2 2 2 2 2 2 2 |
| Controlled Vocabulary 1 | Seed Number | Pod DW | Seed DW | SSS | Shelling % | Seed Protein | Seed Lipid | Seed Carb | Oleic Acid | Lignoceric Acid | Linoleic Acid | Linolenic Acid | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Seed number | CO366:0000340 | 1.00 | |||||||||||
| Pod DW | CO366:0000325 | 0.88 | 1.00 | ||||||||||
| Seed DW | CO366:0000337 | 0.94 | 0.97 | 1.00 | |||||||||
| SSS | CO366:0000328 | 0.42 | 0.65 | 0.62 | 1.00 | ||||||||
| Shelling % | CO366:0000334 | −0.30 | −0.49 | −0.37 | −0.32 | 1.00 | |||||||
| Seed Protein | CDNO:0200040 CO366:0000026 | −0.11 | 0.08 | 0.02 | 0.25 | −0.15 | 1.00 | ||||||
| Seed Lipid | CDNO:0200068 CO366:0000023 | 0.44 | 0.38 | 0.41 | 0.31 | −0.20 | 0.09 | 1.00 | |||||
| Seed Carb | CDNO:0200005 CO366:0000309 | 0.00 | −0.10 | −0.07 | −0.22 | 0.14 | −0.91 | −0.40 | 1.00 | ||||
| Oleic acid | CDNO:0200085 | 0.43 | 0.42 | 0.43 | 0.35 | −0.28 | 0.30 | 0.62 | −0.43 | 1.00 | |||
| Lignoceric acid | CDNO:0200081 | 0.48 | 0.47 | 0.51 | 0.32 | −0.26 | 0.18 | 0.38 | −0.21 | 0.55 | 1.00 | ||
| Linoleic acid | CDNO:0200102 | −0.15 | −0.16 | −0.18 | −0.16 | 0.04 | 0.07 | −0.19 | −0.04 | −0.35 | −0.28 | 1.00 | |
| Linolenic acid | CDNO:0200097 | −0.46 | −0.43 | −0.43 | −0.30 | 0.36 | −0.16 | −0.78 | 0.37 | −0.63 | −0.38 | 0.05 | 1.00 |
| Seed Number | Pod DW | Seed DW | SSS | Shelling % | Protein | Lipid | Carb | Linoleic Acid | Oleic Acid | Lignoceric Acid | Linolenic Acid | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Heritability from parent to offspring (regression) | ||||||||||||
| Generation 1 to 2 | 37 | 20 | 28 | 67 | 22 | 27 | 34 | 35 | 9 | 67 | 77 | 44 |
| Generation 2 to 3 | 41 | 6 | 12 | 16 | 7 | 72 | 45 | 58 | 36 | 79 | 62 | 32 |
| Heritability from GxE analysis | ||||||||||||
| Heritability | n/a | n/a | n/a | n/a | n/a | 77 | 63 | 17 | 0 | 0 | 69 | 81 |
| Average across three estimates of heritability | ||||||||||||
| Average | 26 | 19 | 13 | 28 | 10 | 59 | 47 | 37 | 15 | 49 | 69 | 52 |
| Bambara Groundnut Line | Geographical Origin | SI-1 Seed Lipid | SI-2 Omega 3 and 6 FA | SI-3 Seed Lipid and Yield | SI-4 Omega 3 and 6 FA and Yield | SI-5 Yield Only |
|---|---|---|---|---|---|---|
| IPB-Bam2 | Indonesia | + | + | + | + | + |
| IPB-Bam1 | Indonesia | + | + | + | + | + |
| GHC36105 | Indonesia | + | + | + | + | + |
| 99SB4-2NAM-A | Namibia | + | + | + | + | + |
| IPB Bam-6 | Indonesia | + | + | + | + | + |
| BC31107 | Indonesia | + | + | + | + | + |
| 56Acc89MLI-C | Mali | + | + | + | + | |
| 109BWA1BWA-1 | Botswana | + | + | + | + | |
| DodR_CFF | Tanzania | + | + | + | + | + |
| BD | Indonesia | + | + | + | ||
| Cikur 2.1, | Indonesia | + | + | + | ||
| 104S-1913NAM | Namibia | + | + | + | ||
| 91UNISRSWA-B | Swaziland | + | + | + | ||
| BCGC23107 | Indonesia | + | + | |||
| BH16107 | Indonesia | + | ||||
| 92AHM968NAM-C | Namibia | + | + | + | ||
| 100SB16ANMAMCA16 | Namibia | + | + | |||
| LunT | Sierra Leone | + | ||||
| Mottle Black | Ghana | + | + | |||
| Nav4-13 | Ghana | + | + | |||
| TVSU89 | Thailand | + | + | |||
| Zebra Cream-10 | Ghana | + | + | |||
| LBLBE | Ghana | + | + | |||
| DBB | Ghana | + | + | |||
| IITA686_CFF_2016 | Nigeria | + | + |
| Percent of Change to Trait Value from Mean Value (%) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Selection Index | Seed Number | Pod DW | Seed DW | SSS | Shelling % | Lipid | Protein | Carb | Linoleic Acid | Oleic Acid | Linolenic Acid |
| 1: Seed lipid | 53.49 | 51.52 | 63.44 | 13.10 | 2.21 | 8.34 | 3.71 | −1.84 | −2.52 | 8.93 | −7.54 |
| 2: Omega 3 and 6 fatty acid | 7.45 | 32.73 | 34.52 | 16.00 | 4.89 | −7.40 | 2.73 | 0.28 | 2.62 | −5.64 | 19.89 |
| 3: Seed lipid and yield | 64.88 | 67.93 | 74.86 | 12.57 | −3.24 | 7.28 | 2.39 | −1.29 | −2.70 | 9.05 | −9.26 |
| 4: Omega 3,6 fatty acid and yield | 39.74 | 40.13 | 49.99 | 11.68 | 5.49 | −2.51 | 0.25 | 0.26 | 0.57 | −1.83 | 13.89 |
| 5: Yield | 76.23 | 76.89 | 86.24 | 14.70 | −3.46 | 5.26 | −0.70 | −0.21 | −2.02 | 5.24 | −4.77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azman Halimi, R.; Raymond, C.A.; Barkla, B.J.; Mayes, S.; King, G.J. Development of Selection Indices for Improvement of Seed Yield and Lipid Composition in Bambara Groundnut (Vigna subterranea (L.) Verdc.). Foods 2022, 11, 86. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11010086
Azman Halimi R, Raymond CA, Barkla BJ, Mayes S, King GJ. Development of Selection Indices for Improvement of Seed Yield and Lipid Composition in Bambara Groundnut (Vigna subterranea (L.) Verdc.). Foods. 2022; 11(1):86. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11010086
Chicago/Turabian StyleAzman Halimi, Razlin, Carolyn A. Raymond, Bronwyn J. Barkla, Sean Mayes, and Graham J. King. 2022. "Development of Selection Indices for Improvement of Seed Yield and Lipid Composition in Bambara Groundnut (Vigna subterranea (L.) Verdc.)" Foods 11, no. 1: 86. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11010086