
Structure and Anti-Inflammation Potential of Lipoteichoic Acids Isolated from Lactobacillus Strains

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Adhesion Ability of Lactobacillus Strains
2.3. Extraction of LTA
2.4. Structural Analysis of LTA
2.5. Cell Viability Assay
2.6. Cytokine Measurements
2.7. Western Blotting Analysis
2.8. Statistical Analysis
3. Results
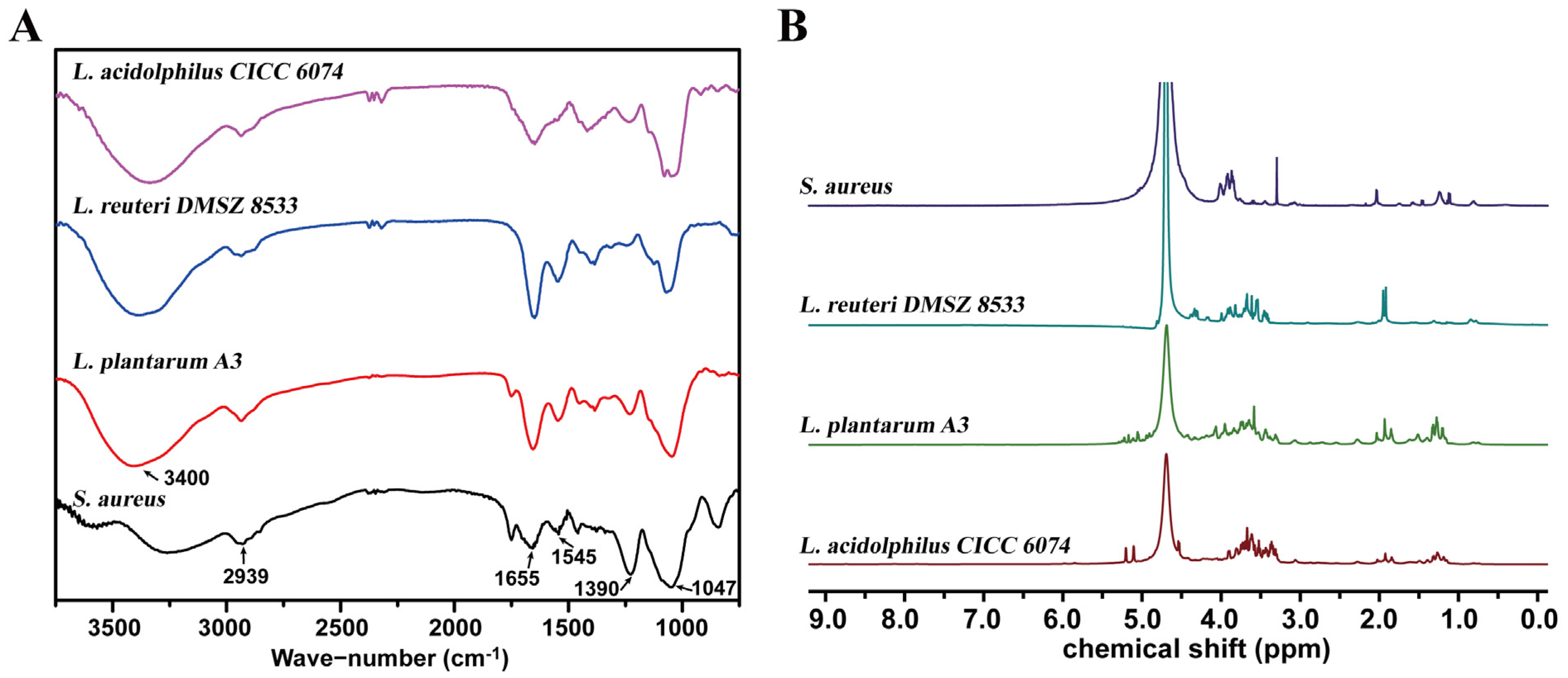
3.1. Structural Properties of Lactobacillus LTA
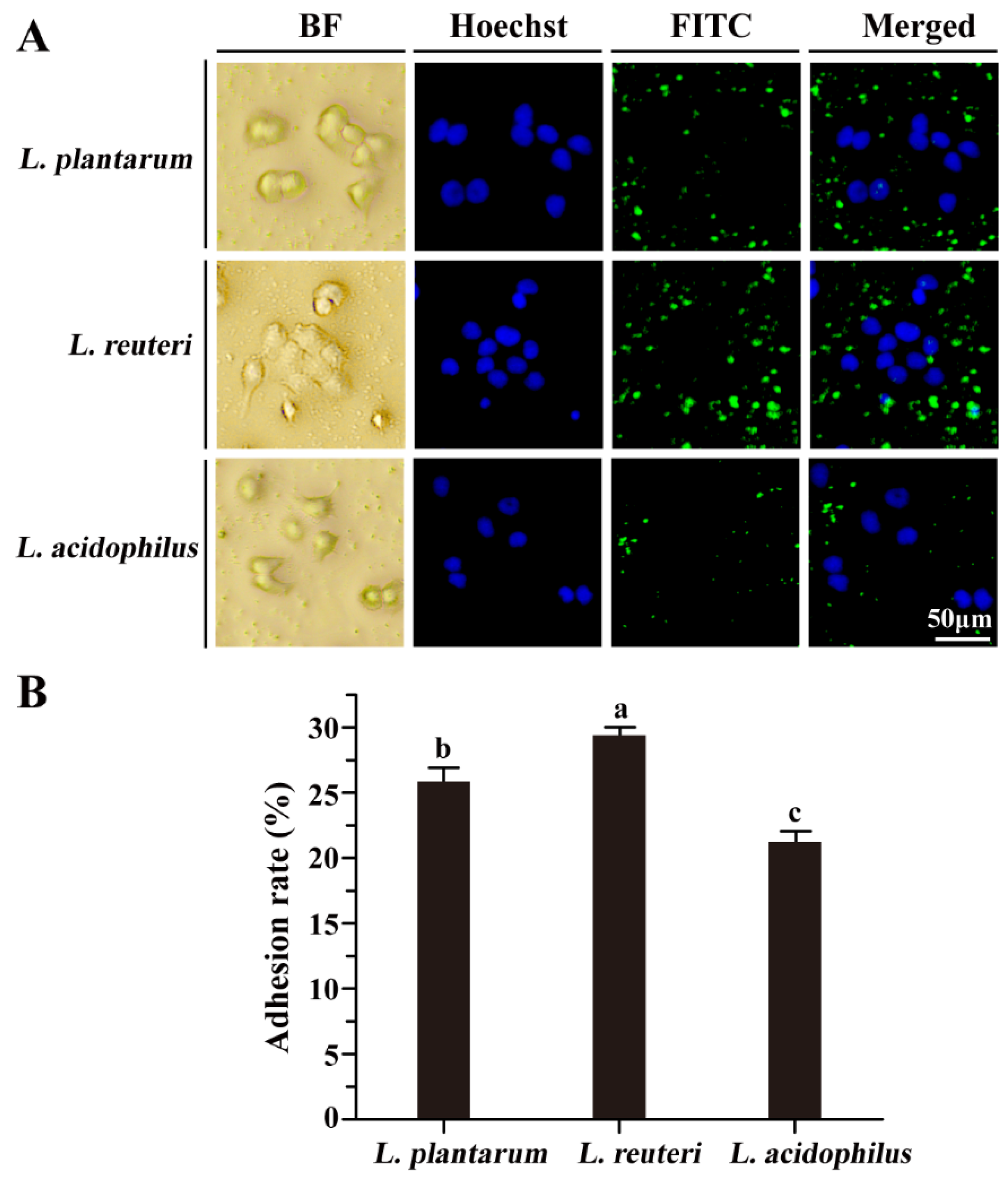
3.2. Adhesion Properties of Lactobacillus Strains to Caco-2 Cells
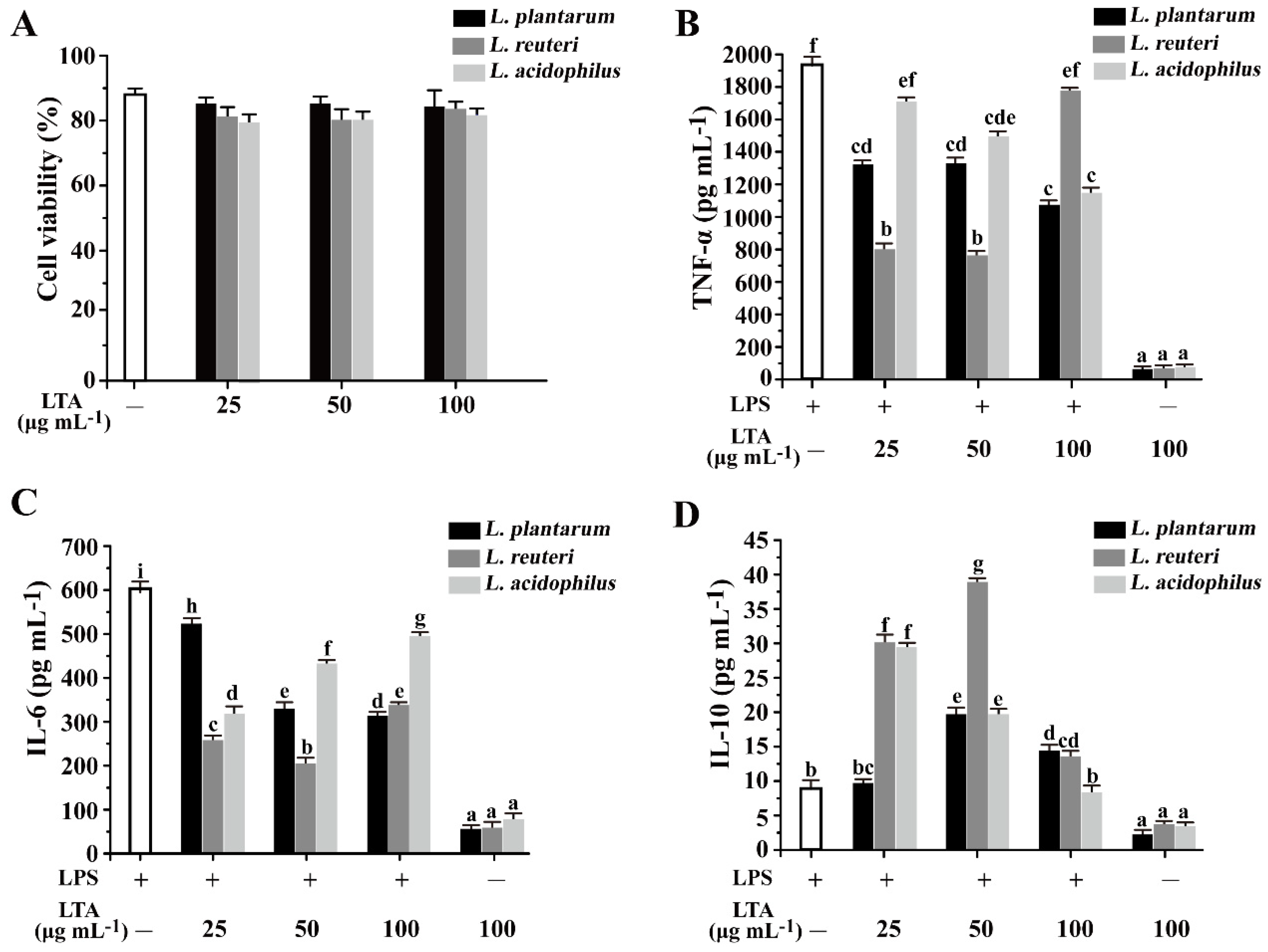
3.3. The Inhibitory Effects of LTAs on the Secretion of Inflammatory Cytokines in RAW 264.7 Cells
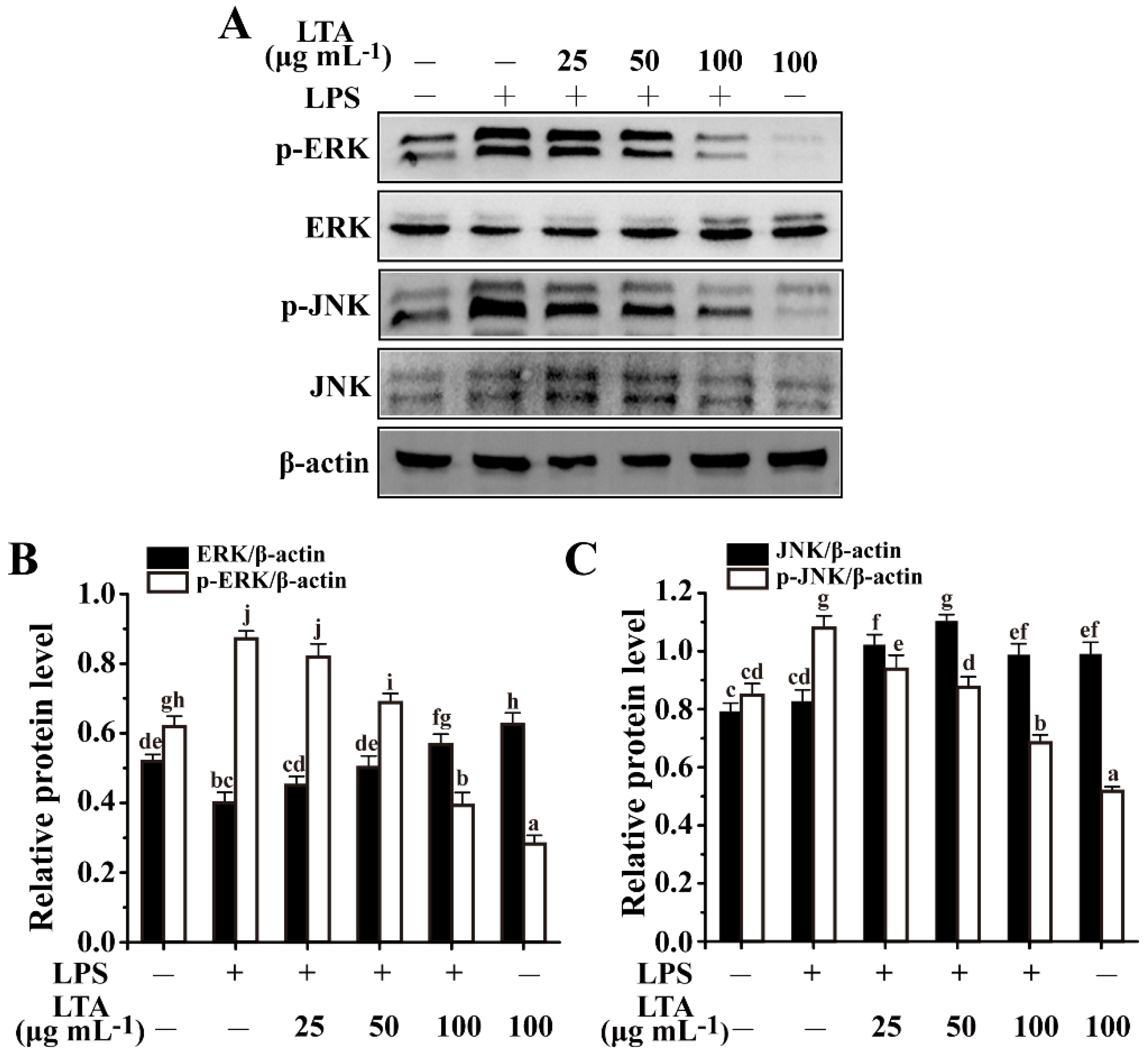
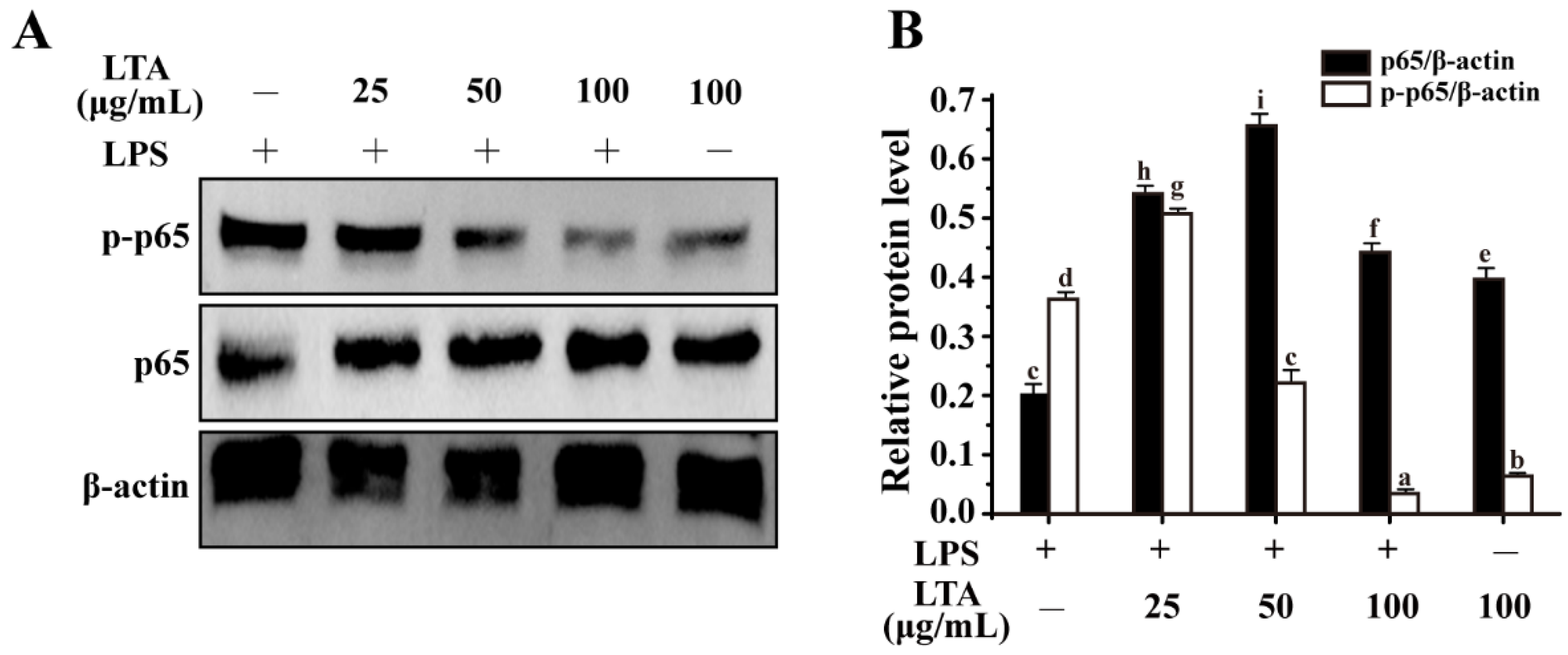
3.4. Anti-Inflammation of LTA Involved in MAPK/NF-κB Signaling Pathways
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Claes, I.J.J.; Segers, M.E.; Verhoeven, T.L.A.; Dusselier, M.; Sels, B.F.; de Keersmaecker, S.C.J.; Vanderleyden, J.; Lebeer, S. Lipoteichoic acid is an important microbe-associated molecular pattern of Lactobacillus rhamnosus GG. Microb. Cell Factories 2012, 11, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeise, K.D.; Woods, R.J.; Huffnagle, G.B. Interplay between Candida albicans and lactic acid bacteria in the gastrointestinal tract: Impact on colonization resistance, microbial carriage, opportunistic infection, and host immunity. Clin. Microbiol. Rev. 2021, 34, e0032320. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Pattern Recognition Receptors and Inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Z.D.; Xua, P.; Wu, Z.; Pan, D. Anti-inflammatory activity of surface layer protein SlpA of Lactobacillus acidophilus CICC 6074 in LPS-induced RAW 264.7 cells and DSS-induced mice colitis. J. Funct. Foods 2018, 51, 16–27. [Google Scholar] [CrossRef]
- Lebeer, S.; Vanderleyden, J.; de Keersmaecker, S.C. Genes and molecules of lactobacilli supporting probiotic action. Microbiol. Mol. Biol. Rev. 2008, 72, 728–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, K.-S.; Baik, J.E.; Han, S.H.; Chung, D.K.; Kim, B.G. Multi-spectrometric analyses of lipoteichoic acids isolated from Lactobacillus plantarum. Biochem. Biophys. Res. Commun. 2011, 407, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Lebeer, S.; Claes, I.J.J.; Vanderleyden, J. Anti-inflammatory potential of probiotics: Lipoteichoic acid makes a difference. Trends Microbiol. 2012, 20, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Matsuguchi, T.; Takagi, A.; Matsuzaki, T.; Nagaoka, M.; Ishikawa, K.; Yokokura, T.; Yoshikai, Y. Lipoteichoic acids from Lactobacillus strains elicit strong tumor necrosis factor alpha-inducing activitives in macrophages through toll-like receptor 2. Clin. Diagn. Lab. Immunol. 2003, 10, 259–266. [Google Scholar]
- Neuhaus, F.C.; Baddiley, J. A continuum of anionic charge: Structures and functions of D-alanyl-teichoic acids in gram-positive bacteria. Microbiol. Mol. Biol. Rev. 2003, 67, 686–723. [Google Scholar] [CrossRef] [Green Version]
- Ginsburg, I. Role of lipoteichoic acid in infection and inflammation. Lancet Infect. Dis. 2002, 2, 171–179. [Google Scholar] [CrossRef]
- Lee, I.T.; Lee, C.-W.; Tung, W.-H.; Wang, S.-W.; Lin, C.-C.; Shu, J.-C.; Yang, C.-M. Cooperation of TLR2 with MyD88, PI3K, and Rac1 in lipoteichoic acid-induced cPLA(2)/COX-2-dependent airway inflammatory responses. Am. J. Pathol. 2010, 176, 1671–1684. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.-S.; Sim, J.; Yun, C.; Han, S.H. Lipoteichoic acids as a major virulence factor causing inflammatory responses via Toll-like receptor 2. Arch. Pharmacal Res. 2016, 39, 1519–1529. [Google Scholar] [CrossRef] [PubMed]
- Shiraishi, T.; Yokota, S.; Fukiya, S.; Yokota, A. Structural diversity and biological significance of lipoteichoic acid in Gram-positive bacteria: Focusing on beneficial probiotic lactic acid bacteria. Biosci. Microbiota Food Health 2016, 35, 147–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.B.; Qi, L.; Mei, L.; Wu, Z.; Wang, H. C. butyricum lipoteichoic acid inhibits the inflammatory response and apoptosis in HT-29 cells induced by S. aureus lipoteichoic acid. Int. J. Biol. Macromol. 2016, 88, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Lee, Y.D.; Kim, M.; Kim, H.; Chung, D.K. Combination treatment with lipoteichoic acids isolated from Lactobacillus plantarum and Staphylococcus aureus alleviates atopic dermatitis via upregulation of CD55 and CD59. Immunol. Lett. 2019, 214, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.F.; Kim, H.; Kim, H.R.; Gim, M.G.; Chung, D.K. Different immune regulatory potential of Lactobacillus plantarum and Lactobacillus sakei isolated from kimchi. J. Microbiol. Biotechnol. 2014, 24, 1629–1635. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.W.; Kang, S.-S.; Woo, S.-J.; Park, O.-J.; Ahn, K.B.; Song, K.-D.; Lee, H.-K.; Yun, C.-H.; Han, S.H. Lipoteichoic acid of probiotic Lactobacillus plantarum attenuates poly I: C-induced IL-8 production in porcine intestinal epithelial cells. Front. Microbiol. 2017, 8, 1827. [Google Scholar] [CrossRef]
- Jung, B.-J.; Kim, H.; Chung, D.-K. Differential immunostimulatory effects of lipoteichoic acids isolated from four strains of Lactiplantibacillus plantarum. Appl. Sci. 2022, 12, 954. [Google Scholar] [CrossRef]
- Jayashree, S.; Karthikeyan, R.; Nithyalakshmi, S.; Ranjani, J.; Gunasekaran, P.; Rajendhran, J. Anti-adhesion property of the potential probiotic strain Lactobacillus fermentum 8711 against methicillin-resistant Staphylococcus aureus (MRSA). Front. Microbiol. 2018, 9, 411. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Pan, D.; Yang, Y.; Jiang, X.; Zhang, J.; Zeng, X.; Wu, Z.; Sun, Y.; Guo, Y. Effect of Lactobacillus acidophilus CICC 6074 S-layer protein on colon cancer HT-29 cell proliferation and apoptosis. J. Agric. Food Chem. 2020, 68, 2639–2647. [Google Scholar] [CrossRef]
- Ryu, Y.H.; Baik, J.E.; Yang, J.S.; Kang, S.; Im, J.; Yun, C.; Kim, D.W.; Lee, K.; Chung, D.K.; Ju, H.R.; et al. Differential immunostimulatory effects of Gram-positive bacteria due to their lipoteichoic acids. Int. Immunopharmacol. 2009, 9, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.B.; Qi, L.; Wu, Z.; Mei, L.; Wang, H. Different effects of lipoteichoic acid from C. butyricum and S. aureus on inflammatory responses of HT-29 cells. Int. J. Biol. Macromol. 2016, 87, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.S.; Ryu, Y.H.; Baik, J.E.; Yun, C.; Lee, K.; Chung, D.K.; Han, S.H. Lipoteichoic acid from Lactobacillus plantarum induces nitric oxide production in the presence of interferon-gamma in murine macrophages. Mol. Immunol. 2011, 48, 2170–2177. [Google Scholar] [CrossRef] [PubMed]
- He, M.T.; Park, S.H.; Kim, Y.S.; Lee, A.Y.; Cho, E.J. Protective effect of membrane-free stem cells against lipopolysaccharide and interferon-gamma-stimulated inflammatory responses in RAW 264.7 macrophages. Int. J. Mol. Sci. 2021, 22, 6894. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.-Y.; Ye, X.-J.; He, X.-H.; Ouyang, D.-Y. The signaling pathways regulating NLRP3 inflammasome activation. Inflammation 2021, 44, 1229–1245. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhang, D.; Qi, W.; Hong, T.; Xiong, T.; Wu, T.; Geng, F.; Xie, M.; Nie, S. Exopolysaccharides from Lactobacillus plantarum NCU116 facilitate intestinal homeostasis by modulating intestinal epithelial regeneration and microbiota. J. Agric. Food Chem. 2021, 69, 7863–7873. [Google Scholar] [CrossRef] [PubMed]
- Grangette, C.; Nutten, S.; Palumbo, E.; Mercenier, A. Enhanced antiinflammatory capacity of a Lactobacillus plantarum mutant synthesizing modified teichoic acids. Proc. Natl. Acad. Sci. USA 2005, 102, 10321–10326. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Gao, Q.; Min, M.; Zhang, C.; Peng, S.; Shi, Z. Ability of Lactobacillus plantarum lipoteichoic acid to inhibit Vibrio anguillarum-induced inflammation and apoptosis in silvery pomfret (Pampus argenteus) intestinal epithelial cells. Fish Shellfish. Immunol. 2016, 54, 573–579. [Google Scholar] [CrossRef]
- Li, H.; Yoon, J.-H.; Won, H.-J.; Ji, H.-S.; Yuk, H.J.; Park, K.H.; Park, H.-Y.; Jeong, T.-S. Isotrifoliol inhibits pro-inflammatory mediators by suppression of TLR/NF-kappa B and TLR/MAPK signaling in LPS-induced RAW264.7 cells. Int. Immunopharmacol. 2017, 45, 110–119. [Google Scholar] [CrossRef]
- Mohamadzadeh, M.; Pfeiler, E.A.; Brown, J.B.; Zadeh, M.; Gramarossa, M.; Managlia, E.; Bere, P.; Sarraj, B.; Khan, M.W.; Pakanati, K.C.; et al. Regulation of induced colonic inflammation by Lactobacillus acidophilus deficient in lipoteichoic acid. Proc. Natl. Acad. Sci. USA 2011, 108, 4623–4630. [Google Scholar] [CrossRef] [Green Version]
- Claes, I.J.J.; Lebeer, S.; Shen, C.; Verhoeven, T.L.A.; Dilissen, E.; de Hertogh, G.; Bullens, D.M.A.; Ceuppens, J.L.; van Assche, G.; Vermeire, S.; et al. Impact of lipoteichoic acid modification on the performance of the probiotic Lactobacillus rhamnosus GG in experimental colitis. Clin. Exp. Immunol. 2010, 162, 306–314. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Q.; Guo, Y.; Yang, G.; Cui, L.; Wu, Z.; Zeng, X.; Pan, D.; Cai, Z. Structure and Anti-Inflammation Potential of Lipoteichoic Acids Isolated from Lactobacillus Strains. Foods 2022, 11, 1610. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11111610
Lu Q, Guo Y, Yang G, Cui L, Wu Z, Zeng X, Pan D, Cai Z. Structure and Anti-Inflammation Potential of Lipoteichoic Acids Isolated from Lactobacillus Strains. Foods. 2022; 11(11):1610. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11111610
Chicago/Turabian StyleLu, Qianqian, Yingqi Guo, Guo Yang, Lei Cui, Zhen Wu, Xiaoqun Zeng, Daodong Pan, and Zhendong Cai. 2022. "Structure and Anti-Inflammation Potential of Lipoteichoic Acids Isolated from Lactobacillus Strains" Foods 11, no. 11: 1610. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11111610