
Early Discrimination and Prediction of C. fimbriata-Infected Sweetpotatoes during the Asymptomatic Period Using Electronic Nose

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
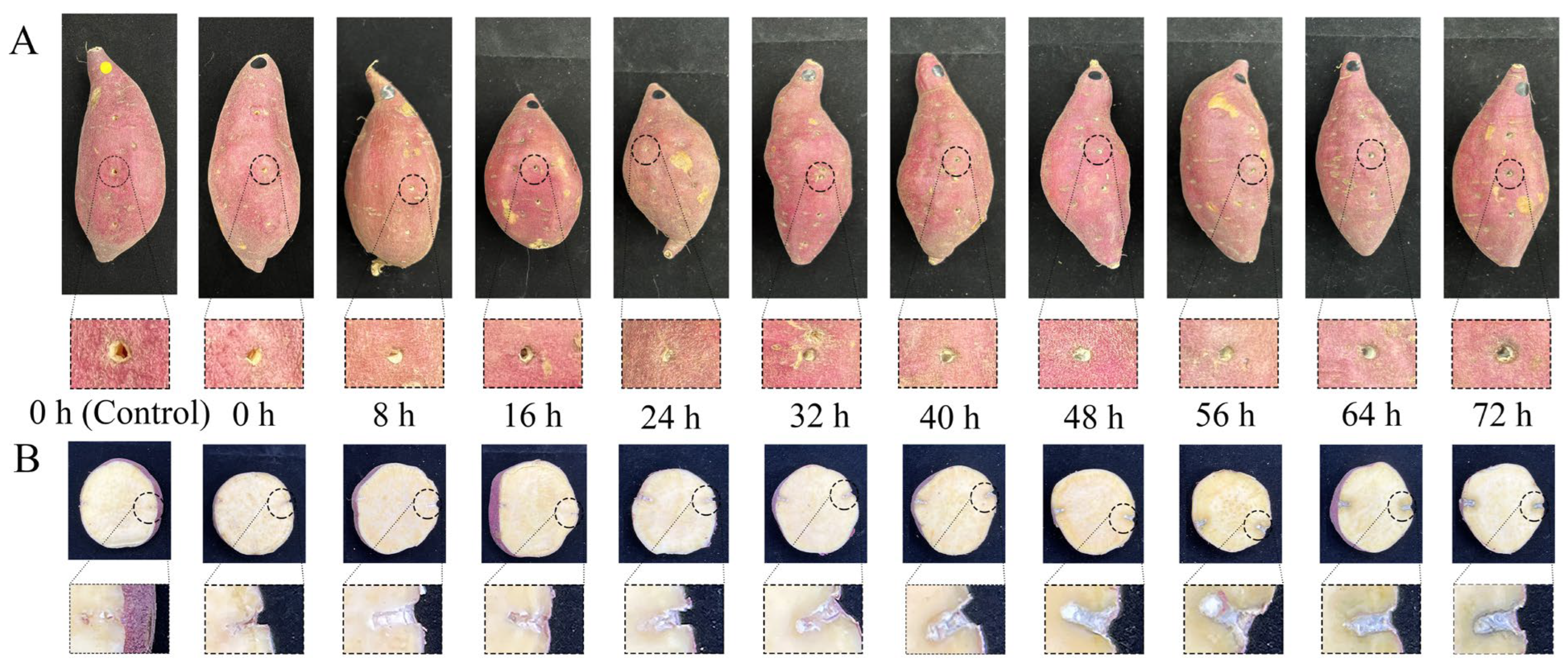
2.1. Sample Preparation
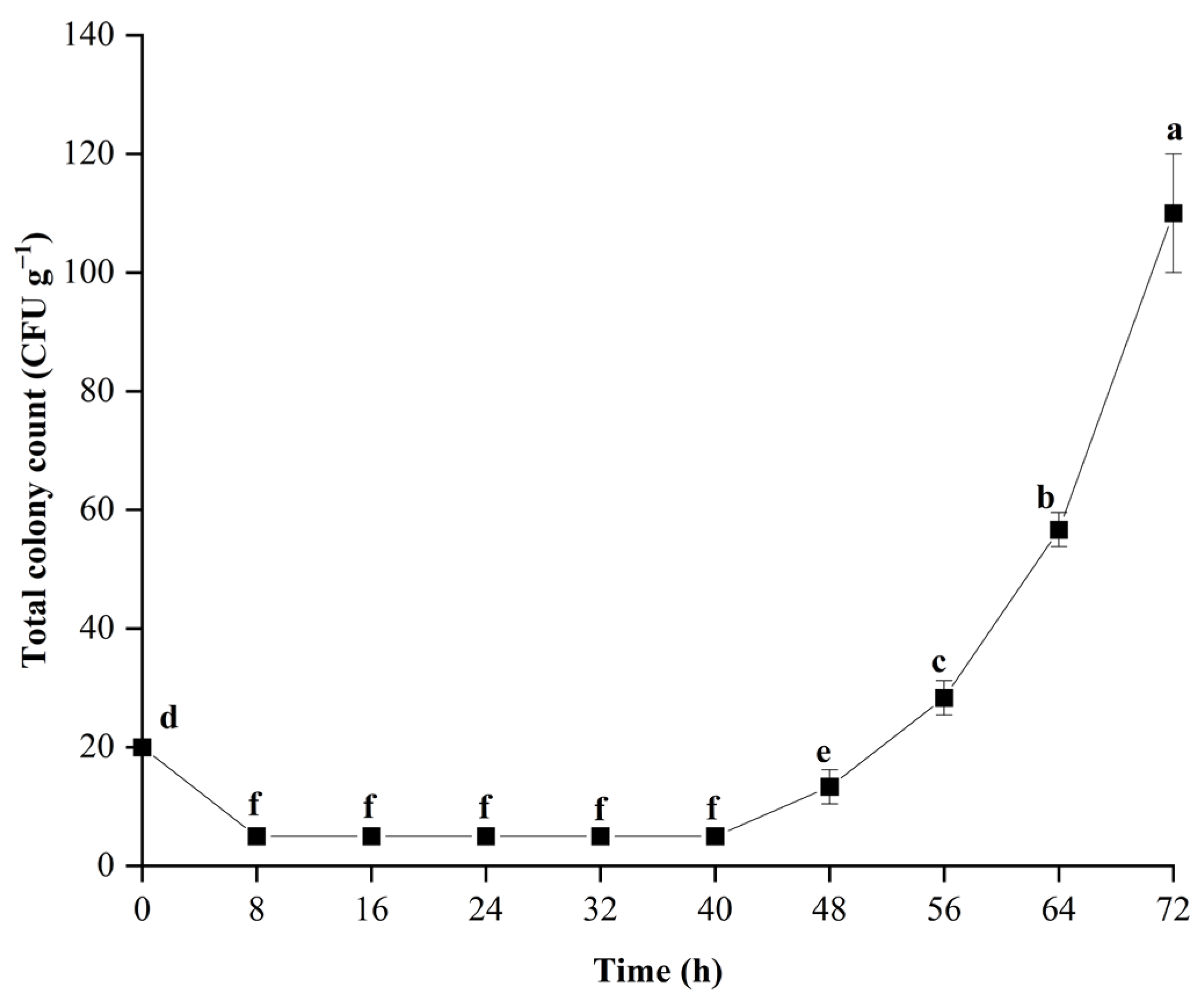
2.2. Colony Count
2.3. Headspace Solid-Phase Microextraction/Gas Chromatography–Mass Spectrometry (HS-SPME/GC–MS) Detection
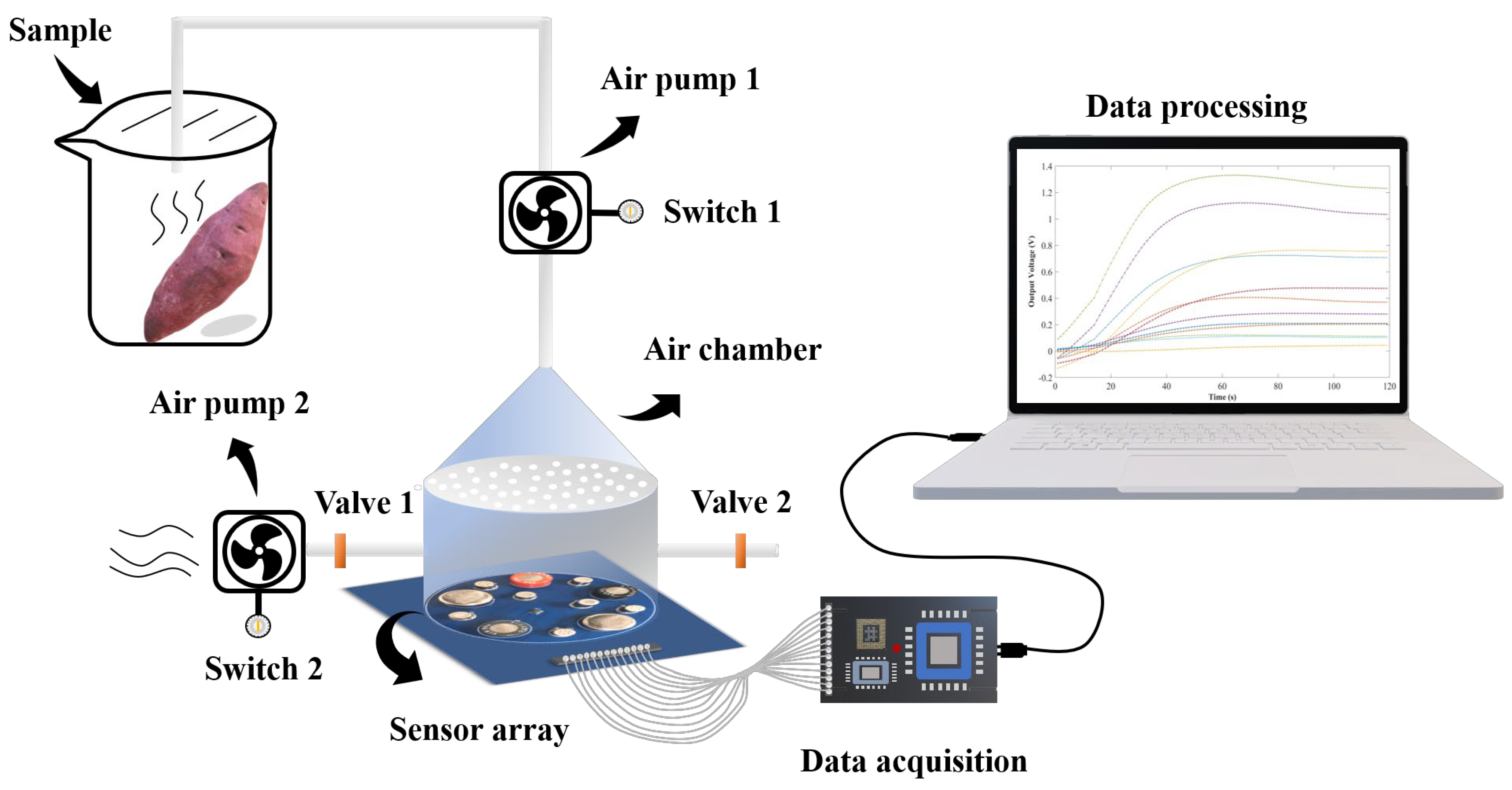
2.4. E-Nose Detection
2.5. Data Analysis
3. Results and Discussion
3.1. Growth of C. fimbriata during Infection in Sweetpotatoes
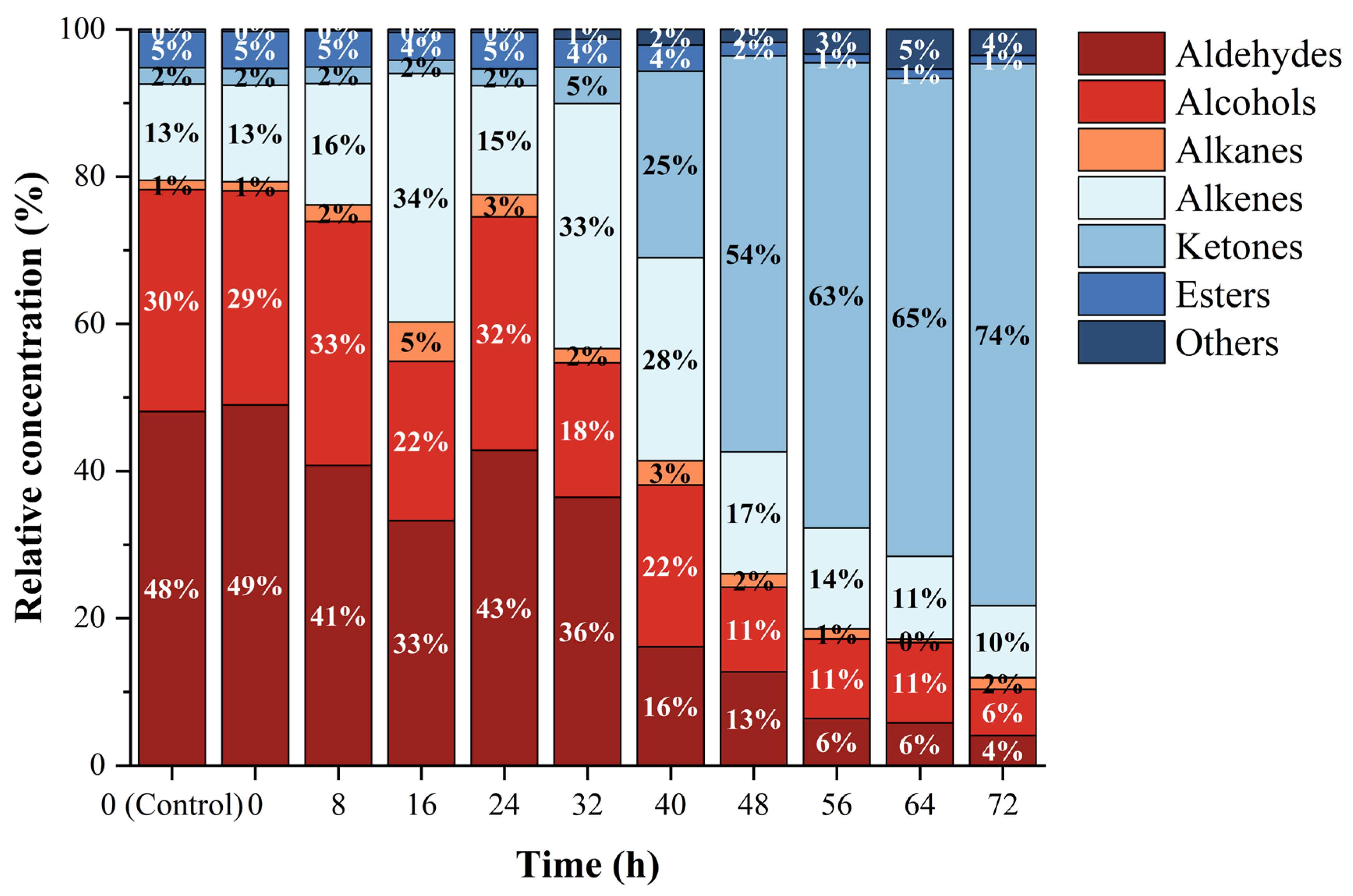
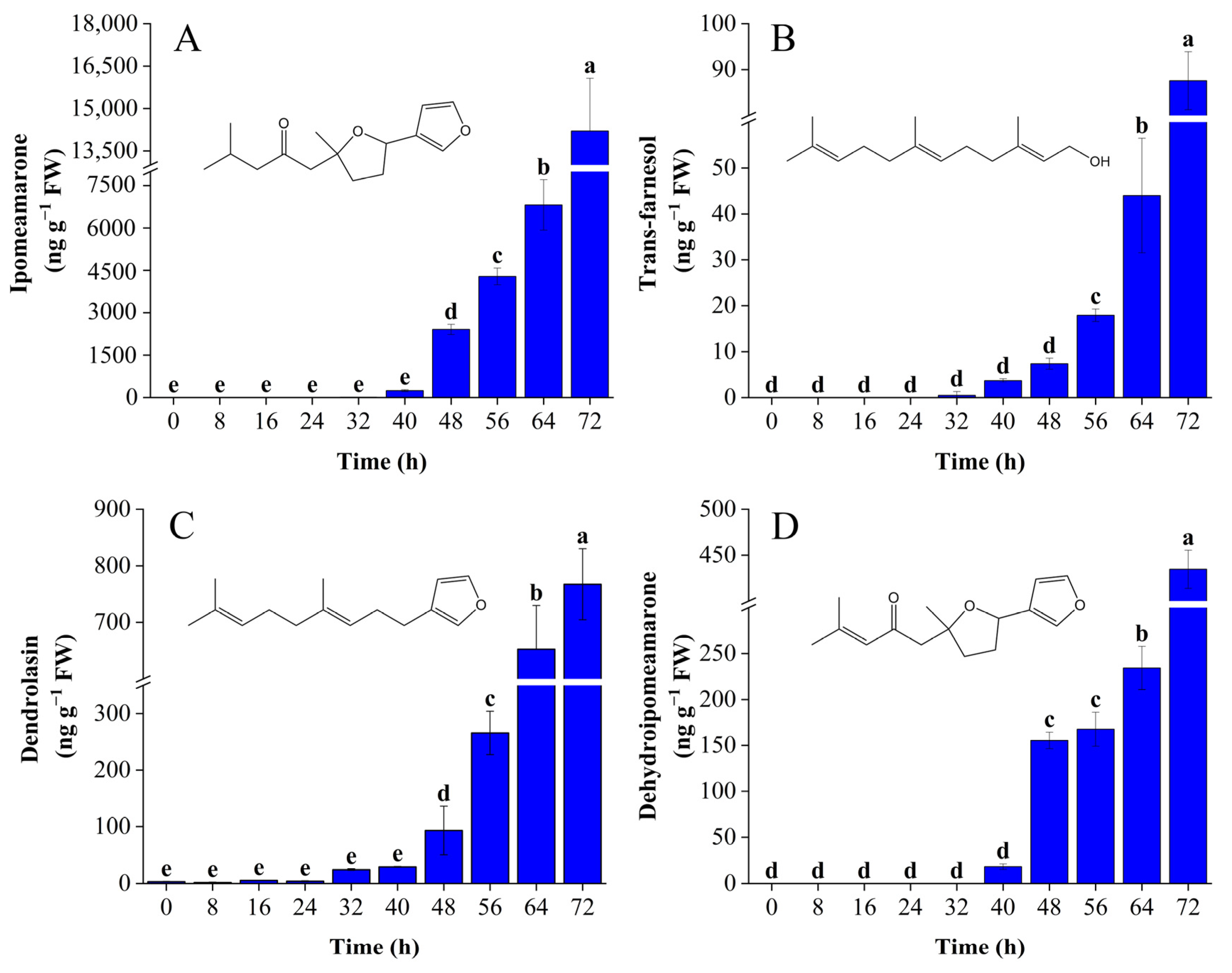
3.2. Analysis of Volatile Compounds by HS-SPME/GC–MS
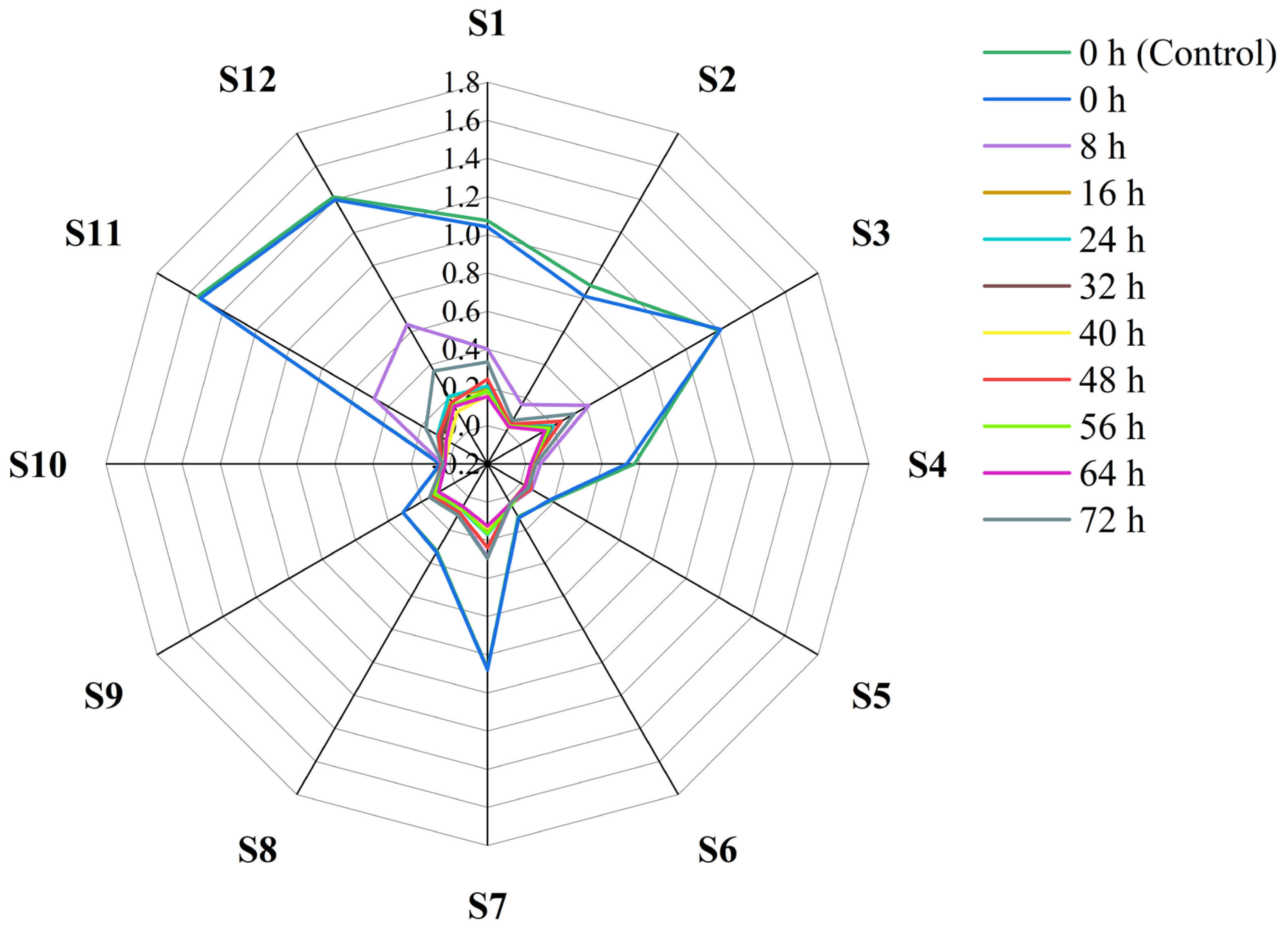
3.3. E-nose Response Signal for Volatile Compounds in Sweetpotatoes
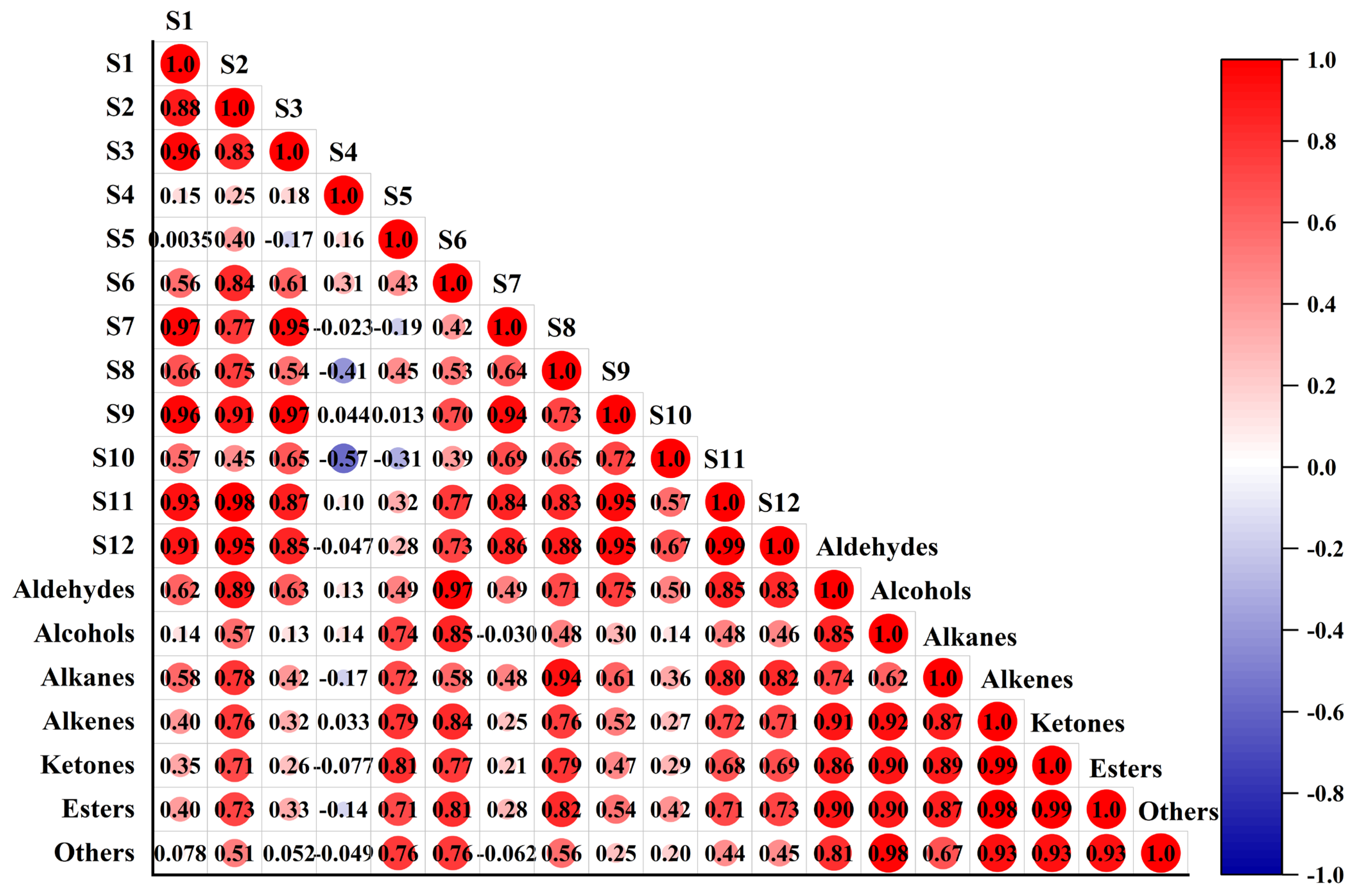
3.4. Correlation Analysis between E-Nose and HS-SPME/GC–MS
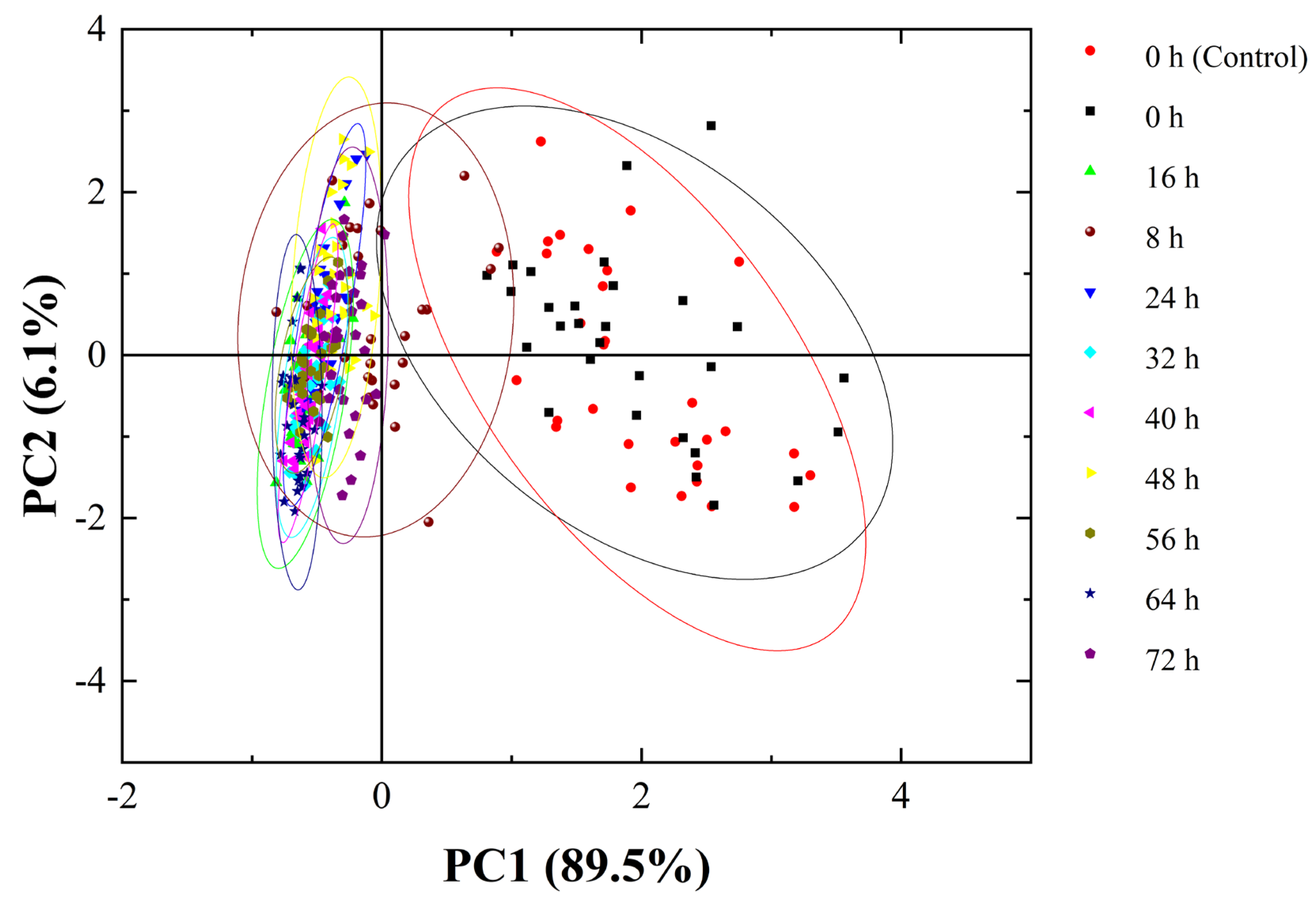
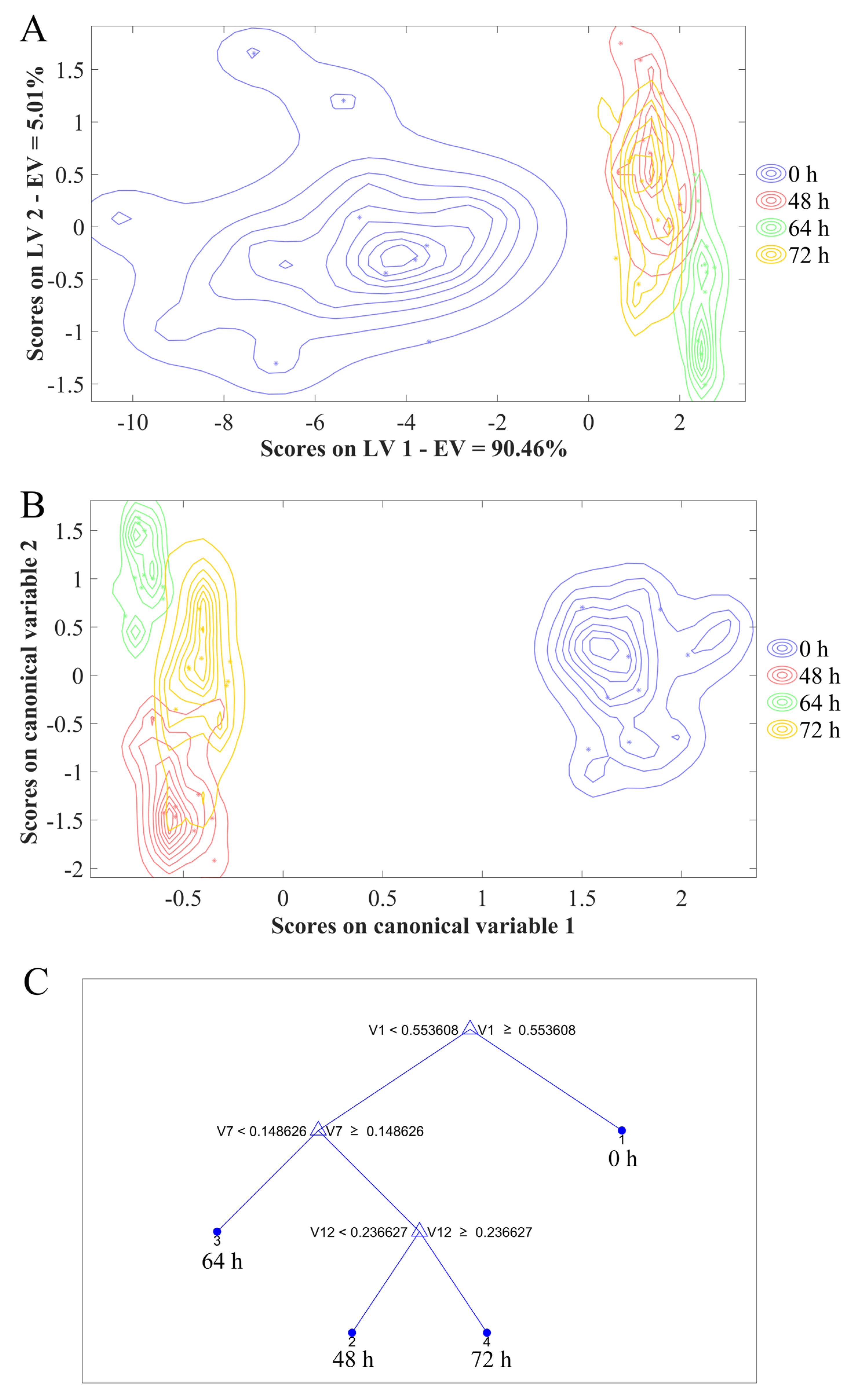
3.5. Early Discrimination of C. fimbriata-Infected Sweetpotatoes Combined with Chemometrics
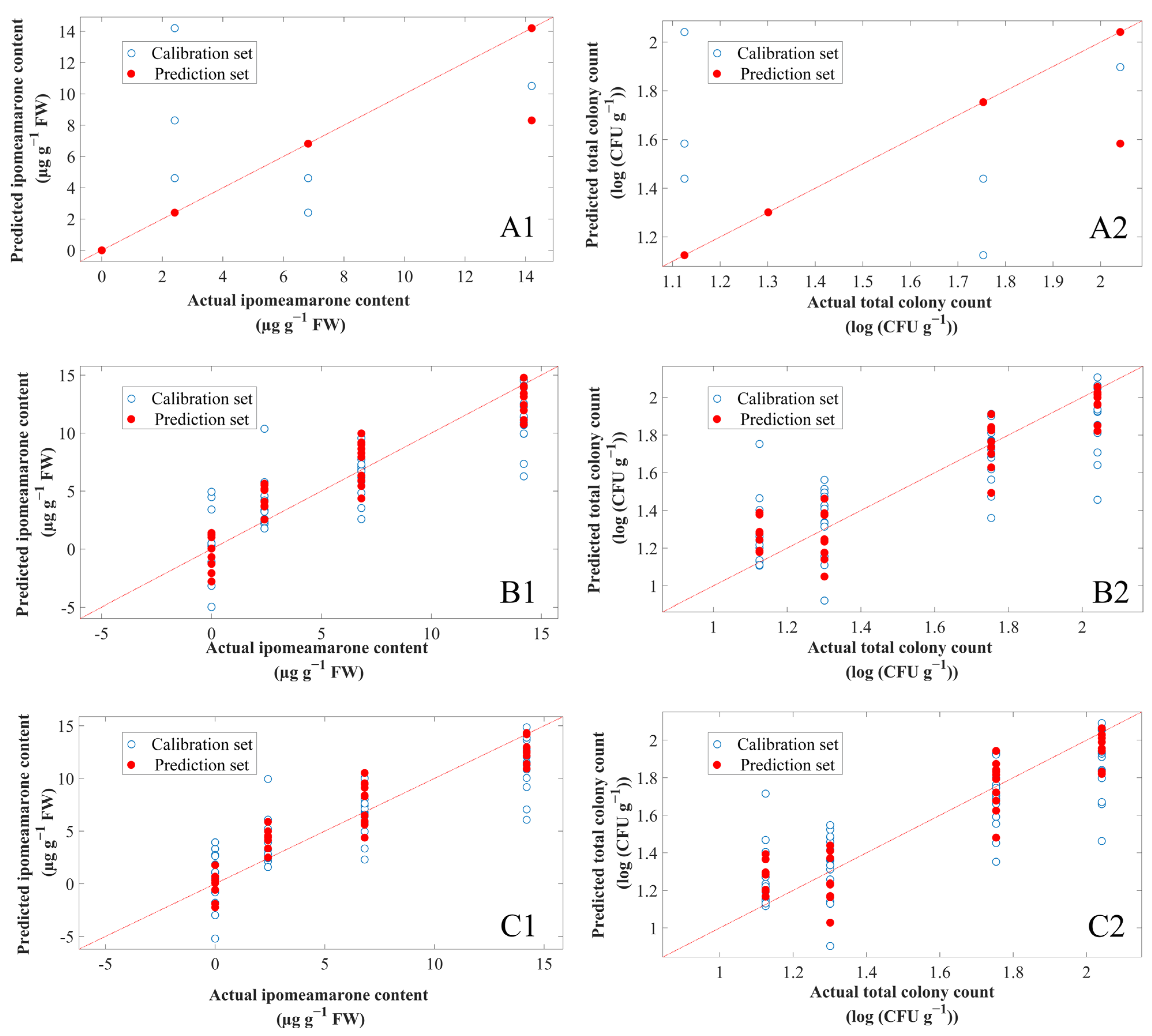
3.6. Prediction of Ipomeamarone Content and Total Colony Count Using E-Nose Combined with Chemometrics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tsai, Y.-J.; Lin, L.-Y.; Yang, K.-M.; Chiang, Y.-C.; Chen, M.-H.; Chiang, P.-Y. Effects of Roasting Sweet Potato (Ipomoea batatas L. Lam.): Quality, Volatile Compound Composition, and Sensory Evaluation. Foods 2021, 10, 2602. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Pan, Z.; Yang, C.; Jia, Z.; Guo, X. Comparative Assessment of Phenolic Profiles, Cellular Antioxidant and Antiproliferative Activities in Ten Varieties of Sweet Potato (Ipomoea batatas) Storage Roots. Molecules 2019, 24, 4476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, M.K. A comprehensive review of sweet potato (Ipomoea batatas [L.] Lam): Revisiting the associated health benefits. Trends Food Sci. Technol. 2021, 115, 512–529. [Google Scholar] [CrossRef]
- Pang, L.; Lu, G.; Cheng, J.; Lu, X.; Ma, D.; Li, Q.; Li, Z.; Zheng, J.; Zhang, C.; Pan, S. Physiological and biochemical characteristics of sweet potato (Ipomoea batatas (L.) Lam) roots treated by a high voltage alternating electric field during cold storage. Postharvest Biol. Technol. 2021, 180, 111619. [Google Scholar] [CrossRef]
- Liu, R.; Yu, Z.-L.; Sun, Y.-L.; Zhou, S.-M. The enzymatic browning reaction inhibition effect of strong acidic electrolyzed water on different parts of sweet potato slices. Food Biosci. 2021, 43, 101252. [Google Scholar] [CrossRef]
- Xu, M.; Guo, J.; Li, T.; Zhang, C.; Peng, X.; Xing, K.; Qin, S. Antibiotic Effects of Volatiles Produced by Bacillus tequilensis XK29 against the Black Spot Disease Caused by Ceratocystis fimbriata in Postharvest Sweet Potato. J. Agric. Food Chem. 2021, 69, 13045–13054. [Google Scholar] [CrossRef]
- Pang, L.-J.; Adeel, M.; Shakoor, N.; Guo, K.-R.; Ma, D.-F.; Ahmad, M.A.; Lu, G.-Q.; Zhao, M.-H.; Li, S.-E.; Rui, Y.-K. Engineered Nanomaterials Suppress the Soft Rot Disease (Rhizopus stolonifer) and Slow Down the Loss of Nutrient in Sweet Potato. Nanomaterials 2021, 11, 2572. [Google Scholar] [CrossRef]
- Wang, C.-J.; Wang, Y.-Z.; Chu, Z.-H.; Wang, P.-S.; Liu, B.-Y.; Li, B.-Y.; Yu, X.-L.; Luan, B.-H. Endophytic Bacillus amyloliquefaciens YTB1407 elicits resistance against two fungal pathogens in sweet potato (Ipomoea batatas (L.) Lam.). J. Plant Physiol. 2020, 253, 153260. [Google Scholar] [CrossRef]
- Li, X.; Liu, M.; Huang, T.; Yang, K.; Zhou, S.; Li, Y.; Tian, J. Antifungal effect of nerol via transcriptome analysis and cell growth repression in sweet potato spoilage fungi Ceratocystis fimbriata. Postharvest Biol. Technol. 2020, 171, 111343. [Google Scholar] [CrossRef]
- Mohsin, S.M.; Hasanuzzaman, M.; Parvin, K.; Morokuma, M.; Fujita, M. Effect of tebuconazole and trifloxystrobin on Ceratocystis fimbriata to control black rot of sweet potato: Processes of reactive oxygen species generation and antioxidant defense responses. World J. Microbiol. Biotechnol. 2021, 37, 1–16. [Google Scholar] [CrossRef]
- Stahr, M.; Quesada-Ocampo, L.M. Assessing the Role of Temperature, Inoculum Density, and Wounding on Disease Progression of the Fungal Pathogen Ceratocystis fimbriata Causing Black Rot in Sweetpotato. Plant Dis. 2020, 104, 930–937. [Google Scholar] [CrossRef] [PubMed]
- Paul, N.C.; Nam, S.-S.; Kachroo, A.; Kim, Y.-H.; Yang, J.-W. Characterization and pathogenicity of sweet potato (Ipomoea batatas) black rot caused by Ceratocystis fimbriata in Korea. Eur. J. Plant Pathol. 2018, 152, 833–840. [Google Scholar] [CrossRef]
- Tian, J.; Pan, C.; Zhang, M.; Gan, Y.Y.; Pan, S.Y.; Liu, M.; Li, Y.X.; Zeng, X.B. Induced cell death in Ceratocystis fimbriata by pro-apoptotic activity of a natural organic compound, perillaldehyde, through Ca2+ overload and accumulation of reactive oxygen species. Plant Pathol. 2018, 68, 344–357. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, T.; Liu, Y.; Li, X.; Zhang, C.; Feng, Z.; Peng, X.; Li, Z.; Qin, S.; Xing, K. Volatile Organic Compounds Produced by Pseudomonas chlororaphis subsp. aureofaciens SPS-41 as Biological Fumigants to Control Ceratocystis fimbriata in Postharvest Sweet Potatoes. J. Agric. Food Chem. 2019, 67, 3702–3710. [Google Scholar] [CrossRef]
- Usuki, Y.; Deguchi, T.; Iio, H. A New Concise Synthesis of (+)-Ipomeamarone, (−)-Ngaione, and Their Stereoisomers. Chem. Lett. 2014, 43, 1882–1884. [Google Scholar] [CrossRef]
- Boyd, M.R.; Burka, L.T.; Harris, T.M.; Willson, B.J. Lung-toxic furanoterpenoids produced by sweet potatoes (Ipomoea batatas) following microbial infection. Biochim. Biophys. Acta (BBA)-Lipids Lipid Metab. 1974, 337, 184–195. [Google Scholar] [CrossRef]
- Pandey, G. Acute toxicity of ipomeamarone, a phytotoxin isolated from the injured sweet potato. Pharmacogn. Mag. 2008, 4, 89–92. [Google Scholar]
- Zhang, Y.; Li, T.; Xu, M.; Guo, J.; Zhang, C.; Feng, Z.; Peng, X.; Li, Z.; Xing, K.; Qin, S. Antifungal effect of volatile organic compounds produced by Pseudomonas chlororaphis subsp. aureofaciens SPS-41 on oxidative stress and mitochondrial dysfunction of Ceratocystis fimbriata. Pestic. Biochem. Physiol. 2021, 173, 104777. [Google Scholar] [CrossRef]
- Sugimura, T.; Koguro, K.; Tai, A. Total syntheses of (+)-ipomeamarone and (−)-ngaione. Tetrahedron Lett. 1993, 34, 509–512. [Google Scholar] [CrossRef]
- Wang, A.; Luca, A.; Edelenbos, M. Emission of volatile organic compounds from yellow onion (Allium cepa L.) bulbs during storage. J. Food Sci. Technol. 2019, 56, 2940–2948. [Google Scholar] [CrossRef]
- Chalupowicz, D.; Veltman, B.; Droby, S.; Eltzov, E. Evaluating the use of biosensors for monitoring of Penicillium digitatum infection in citrus fruit. Sensors Actuators B Chem. 2020, 311, 127896. [Google Scholar] [CrossRef]
- Gong, D.; Bi, Y.; Zong, Y.; Li, Y.; Sionov, E.; Prusky, D. Characterization and sources of volatile organic compounds produced by postharvest pathogenic fungi colonized fruit. Postharvest Biol. Technol. 2022, 188, 111903. [Google Scholar] [CrossRef]
- Biancolillo, A.; Aloia, R.; Rossi, L.; D’Archivio, A.A. Organosulfur volatile profiles in Italian red garlic (Allium Sativum L.) varieties investigated by HS-SPME/GC-MS and chemometrics. Food Control 2021, 131, 108477. [Google Scholar] [CrossRef]
- Shen, X.; Wang, Y.; Ran, L.; Liu, R.; Sun, X.; Hu, L.; Xiao, Y.; Chen, F. Flavor deterioration of liquid endosperm in postharvest tender coconut revealed by LC-MS-based metabolomics, GC-IMS and E-tongue. Postharvest Biol. Technol. 2022, 187, 111866. [Google Scholar] [CrossRef]
- Li, J.; Xu, Y.; Du, W.; Jin, L.; Ren, P.; Ren, F.; Xie, J.C. Comparative analysis of aroma compounds in Chinese traditional dry-rendered fat by HS/GC-IMS, SPME/GC-MS, and SPME/GC-O. J. Food Compos. Anal. 2022, 107, 104378. [Google Scholar] [CrossRef]
- Yu, S.; Huang, X.; Wang, L.; Ren, Y.; Zhang, X.; Wang, Y. Characterization of selected Chinese soybean paste based on flavor profiles using HS-SPME-GC/MS, E-nose and E-tongue combined with chemometrics. Food Chem. 2022, 375, 131840. [Google Scholar] [CrossRef]
- Guo, Q.; Adelina, N.M.; Hu, J.; Zhang, L.; Zhao, Y. Comparative analysis of volatile profiles in four pine-mushrooms using HS-SPME/GC-MS and E-nose. Food Control 2021, 134, 108711. [Google Scholar] [CrossRef]
- Gu, S.; Chen, W.; Wang, Z.; Wang, J.; Huo, Y. Rapid detection of Aspergillus spp. infection levels on milled rice by headspace-gas chromatography ion-mobility spectrometry (HS-GC-IMS) and E-nose. LWT 2020, 132, 109758. [Google Scholar] [CrossRef]
- Ali, M.M.; Hashim, N.; Aziz, S.A.; Lasekan, O. Principles and recent advances in electronic nose for quality inspection of agricultural and food products. Trends Food Sci. Technol. 2020, 99, 1–10. [Google Scholar] [CrossRef]
- Rutolo, M.F.; Clarkson, J.P.; Covington, J.A. The use of an electronic nose to detect early signs of soft-rot infection in potatoes. Biosyst. Eng. 2018, 167, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Yu, S.; Xu, H.; Aheto, J.H.; Bonah, E.; Ma, M.; Wu, M.; Zhang, X. Rapid and nondestructive detection of freshness quality of postharvest spinaches based on machine vision and electronic nose. J. Food Saf. 2019, 39, e12708. [Google Scholar] [CrossRef]
- Wen, T.; Zheng, L.; Dong, S.; Gong, Z.; Sang, M.; Long, X.; Luo, M.; Peng, H. Rapid detection and classification of citrus fruits infestation by Bactrocera dorsalis (Hendel) based on electronic nose. Postharvest Biol. Technol. 2018, 147, 156–165. [Google Scholar] [CrossRef]
- Makarichian, A.; Chayjan, R.A.; Ahmadi, E.; Zafari, D. Early detection and classification of fungal infection in garlic (A. sativum) using electronic nose. Comput. Electron. Agric. 2021, 192, 106575. [Google Scholar] [CrossRef]
- Wang, L.; Hu, Q.; Pei, F.; Mugambi, M.A.; Yang, W. Detection and identification of fungal growth on freeze-dried Agaricus bisporus using spectra and olfactory sensors. J. Sci. Food Agric. 2020, 100, 3136–3146. [Google Scholar] [CrossRef] [PubMed]
- Leggieri, M.C.; Mazzoni, M.; Fodil, S.; Moschini, M.; Bertuzzi, T.; Prandini, A.; Battilani, P. An electronic nose supported by an artificial neural network for the rapid detection of aflatoxin B1 and fumonisins in maize. Food Control 2020, 123, 107722. [Google Scholar] [CrossRef]
- Sanaeifar, A.; Li, X.; He, Y.; Huang, Z.; Zhan, Z. A data fusion approach on confocal Raman microspectroscopy and electronic nose for quantitative evaluation of pesticide residue in tea. Biosyst. Eng. 2021, 210, 206–222. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, B.; Dong, J.; Tian, Y.; Lin, Y.; Fang, G.; Wang, S. Identification of mouldy rice using an electronic nose combined with SPME-GC/MS. J. Stored Prod. Res. 2021, 95, 101921. [Google Scholar] [CrossRef]
- Lan, T.; Gao, C.; Yuan, Q.; Wang, J.; Zhang, H.; Sun, X.; Lei, Y.; Ma, T. Analysis of the Aroma Chemical Composition of Commonly Planted Kiwifruit Cultivars in China. Foods 2021, 10, 1645. [Google Scholar] [CrossRef]
- Zhang, D.; Ji, H.; Liu, S.; Gao, J. Similarity of aroma attributes in hot-air-dried shrimp (Penaeus vannamei) and its different parts using sensory analysis and GC–MS. Food Res. Int. 2020, 137, 109517. [Google Scholar] [CrossRef]
- Pang, L.; Wang, J.; Lu, X.; Yu, H. Discrimination of Storage Age for Wheat by E-Nose. Trans. ASABE 2008, 51, 1707–1712. [Google Scholar] [CrossRef]
- Marchal, P.C.; Sanmartin, C.; Martínez, S.S.; Ortega, J.G.; Mencarelli, F.; García, J.G. Prediction of Fruity Aroma Intensity and Defect Presence in Virgin Olive Oil Using an Electronic Nose. Sensors 2021, 21, 2298. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhao, N.; Zhou, D.; Sun, Y.; Sun, K.; Pan, L.; Tu, K. Discrimination and growth tracking of fungi contamination in peaches using electronic nose. Food Chem. 2018, 262, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, J.S.; Koutsoumanis, K.P. Towards lag phase of microbial populations at growth-limiting conditions: The role of the variability in the growth limits of individual cells. Int. J. Food Microbiol. 2016, 224, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Gu, Y. A Machine Learning Method for the Detection of Brown Core in the Chinese Pear Variety Huangguan Using a MOS-Based E-Nose. Sensors 2020, 20, 4499. [Google Scholar] [CrossRef]
- Nouri, B.; Mohtasebi, S.S.; Rafiee, S. Quality detection of pomegranate fruit infected with fungal disease. Int. J. Food Prop. 2020, 23, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Wamalwa, L.N.; Cheseto, X.; Ouna, E.; Kaplan, F.; Maniania, N.K.; Machuka, J.; Torto, B.; Ghislain, M. Toxic Ipomeamarone Accumulation in Healthy Parts of Sweetpotato (Ipomoea batatas L. Lam) Storage Roots upon Infection by Rhizopus stolonifer. J. Agric. Food Chem. 2014, 63, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Schneider, J.A.; Lee, J.; Naya, Y.; Nakanishi, K.; Oba, K.; Uritani, I. The fate of the phytoalexin ipomeamarone: Furanoterpenes and butenolides from Ceratocystis fimbriata-infected sweet potatoes. Phytochemistry 1984, 23, 759–764. [Google Scholar] [CrossRef]
- Inoue, H.; Oba, K.; Ando, M.; Uritani, I. Enzymatic reduction of dehydroipomeamarone to ipomeamarone in sweet potato root tissue infected by Ceratocystis fimbriata. Physiol. Plant Pathol. 1984, 25, 1–8. [Google Scholar] [CrossRef]
- Barbosa-Pereira, L.; Rojo-Poveda, O.; Ferrocino, I.; Giordano, M.; Zeppa, G. Assessment of volatile fingerprint by HS-SPME/GC-qMS and E-nose for the classification of cocoa bean shells using chemometrics. Food Res. Int. 2019, 123, 684–696. [Google Scholar] [CrossRef] [Green Version]
- Dittrich, I.; Gertz, M.; Maassen-Francke, B.; Krudewig, K.-H.; Junge, W.; Krieter, J. Combining multivariate cumulative sum control charts with principal component analysis and partial least squares model to detect sickness behaviour in dairy cattle. Comput. Electron. Agric. 2021, 186, 106209. [Google Scholar] [CrossRef]
- Li, Y.; Fei, C.; Mao, C.; Ji, D.; Gong, J.; Qin, Y.; Qu, L.; Zhang, W.; Bian, Z.; Su, L.; et al. Physicochemical parameters combined flash GC e-nose and artificial neural network for quality and volatile characterization of vinegar with different brewing techniques. Food Chem. 2021, 374, 131658. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Sun, K.; Zhao, N.; Yang, J.; Zhang, Y.; Ma, C.; Pan, L.; Tu, K. Information fusion of hyperspectral imaging and electronic nose for evaluation of fungal contamination in strawberries during decay. Postharvest Biol. Technol. 2019, 153, 152–160. [Google Scholar] [CrossRef]
- Wang, S.; Hu, X.-Z.; Liu, Y.-Y.; Tao, N.-P.; Lu, Y.; Wang, X.-C.; Lam, W.; Lin, L.; Xu, C.-H. Direct authentication and composition quantitation of red wines based on Tri-step infrared spectroscopy and multivariate data fusion. Food Chem. 2021, 372, 131259. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, D.; Moreira, B.R.D.A.; Júnior, M.R.B.; Papa, J.P.; da Silva, R.P. Predicting on multi-target regression for the yield of sweet potato by the market class of its roots upon vegetation indices. Comput. Electron. Agric. 2021, 191, 106544. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (h) | Aldehydes | Alcohols | Alkanes | Alkenes | Ketones | Esters | Others |
|---|---|---|---|---|---|---|---|
| 0 (Control) | 496.35 ± 44.56 cA | 311.02 ± 61.60 dB | 13.34 ± 1.40 dD | 134.89 ± 7.25 fC | 22.84 ± 3.65 eD | 49.98 ± 4.35 eD | 3.75 ± 1.21 eD |
| 0 | 499.00 ± 26.34 cA | 296.54 ± 108.77 dB | 12.59 ± 1.49 dD | 133.39 ± 15.50 fC | 23.39 ± 3.18 eD | 50.95 ± 14.33 eD | 3.18 ± 0.51 eD |
| 8 | 348.02 ± 24.42 deA | 282.95 ± 15.65 dB | 19.35 ± 0.80 dDE | 140.66 ± 28.37 fC | 19.32 ± 5.43 eDE | 42.00 ± 8.12 eD | 1.68 ± 0.83 eE |
| 16 | 514.80 ± 120.50 cA | 335.73 ± 9.34 dB | 82.85 ± 18.66 bcdC | 522.37 ± 100.61 deA | 28.35 ± 8.50 eC | 58.34 ± 6.76 deC | 6.50 ± 1.59 eC |
| 24 | 411.93 ± 79.23 cdA | 305.17 ± 11.34 dB | 28.90 ± 0.82 cdD | 142.50 ± 32.01 fC | 22.05 ± 5.81 eD | 47.67 ± 7.87 eD | 4.07 ± 0.84 eD |
| 32 | 742.72 ± 50.90 bA | 372.05 ± 17.51 dB | 39.60 ± 6.09 bcdC | 678.17 ± 316.08 dA | 100.69 ± 34.94 eC | 77.71 ± 6.97 cdC | 27.22 ± 6.14 eC |
| 40 | 241.57 ± 36.99 eC | 329.20 ± 33.58 dB | 49.58 ± 7.70 bcdD | 413.32 ± 45.67 eA | 379.07 ± 45.36 eAB | 53.48 ± 16.41 deD | 32.01 ± 3.87 eD |
| 48 | 694.50 ± 125.8 bBC | 626.53 ± 95.87 cC | 100.14 ± 24.43 bcD | 903.59 ± 99.88 cB | 2932.65 ± 249.65 dA | 101.57 ± 15.67 cD | 96.02 ± 42.29 dD |
| 56 | 518.93 ± 43.41 cC | 878.19 ± 149.36 bB | 110.02 ± 6.67 bD | 1112.41 ± 97.31 cB | 5131.48 ± 406.33 cA | 94.84 ± 4.21 cD | 273.18 ± 41.65 cCD |
| 64 | 721.62 ± 28.65 bBC | 1352.64 ± 302.55 aB | 56.87 ± 46.74 bcdC | 1396.61 ± 109.82 bB | 8055.23 ± 1010.49 bA | 160.34 ± 6.00 bC | 669.30 ± 76.45 bBC |
| 72 | 963.91 ± 33.46 aC | 1473.76 ± 29.78 aBC | 376.55 ± 118.15 aC | 2304.64 ± 177.67 aB | 1,7346.42 ± 1682.93 aA | 270.25 ± 34.08 aC | 835.11 ± 67.99 aC |
| Trans-Farnesol | Dendrolasin | Dehydroipomeamarone | Ipomeamarone | |
|---|---|---|---|---|
| Total colony count | 0.978 ** | 0.937 ** | 0.940 ** | 0.968 ** |
| Trans-farnesol | 1 | 0.937 ** | 0.951 ** | 0.987 ** |
| Dendrolasin | 1 | 0.929 ** | 0.933 ** | |
| Dehydroipomeamarone | 1 | 0.973 ** |
| Data Set | Calibration | Cross-Validation | Prediction | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Model | Class | Sens(%) | Spec(%) | Prec(%) | Ac(%) | Sens(%) | Spec(%) | Prec(%) | Ac(%) | Sens(%) | Spec(%) | Prec(%) | Ac(%) |
| PLS-DA | 0 h | 100 | 100 | 100 | 97 | 100 | 100 | 100 | 97 | 100 | 100 | 100 | 100 |
| 48 h | 95 | 100 | 100 | 95 | 100 | 100 | 100 | 100 | 90 | ||||
| 64 h | 100 | 98 | 95 | 100 | 98 | 94 | 100 | 100 | 100 | ||||
| 72 h | 95 | 98 | 95 | 95 | 98 | 95 | 100 | 100 | 100 | ||||
| LDA | 0 h | 100 | 100 | 100 | 96 | 100 | 100 | 100 | 94 | 100 | 100 | 100 | 100 |
| 48 h | 95 | 98 | 95 | 95 | 97 | 91 | 100 | 100 | 100 | ||||
| 64 h | 100 | 98 | 95 | 100 | 97 | 91 | 100 | 100 | 100 | ||||
| 72 h | 90 | 98 | 95 | 81 | 98 | 94 | 100 | 100 | 100 | ||||
| CART | 0 h | 100 | 100 | 100 | 95 | 100 | 100 | 100 | 89 | 100 | 100 | 100 | 97 |
| 48 h | 86 | 98 | 95 | 76 | 95 | 84 | 88 | 100 | 100 | ||||
| 64 h | 95 | 100 | 100 | 90 | 97 | 90 | 100 | 100 | 100 | ||||
| 72 h | 100 | 95 | 88 | 90 | 94 | 83 | 100 | 96 | 90 | ||||
| Data Set | Calibration | Cross-Validation | Prediction | RPD | |||
|---|---|---|---|---|---|---|---|
| Model | RMSEC (µg g−1 FW) | RMSECV (µg g−1 FW) | RMSEP (µg g−1 FW) | ||||
| KNN | 0.836 | 2.183 | 0.834 | 2.200 | 0.932 | 1.410 | 3.852 |
| PLS | 0.771 | 2.581 | 0.723 | 2.840 | 0.875 | 1.909 | 2.845 |
| PCR | 0.776 | 2.556 | 0.724 | 2.835 | 0.877 | 1.891 | 2.872 |
| Data Set | Calibration | Cross-Validation | Prediction | RPD | |||
|---|---|---|---|---|---|---|---|
| Model | RMSEC (log (CFU g−1)) | RMSECV (log (CFU g−1)) | RMSEP (log (CFU g−1)) | ||||
| KNN | 0.744 | 0.184 | 0.757 | 0.179 | 0.909 | 0.110 | 3.306 |
| PLS | 0.741 | 0.185 | 0.644 | 0.217 | 0.867 | 0.133 | 2.734 |
| PCR | 0.735 | 0.187 | 0.661 | 0.212 | 0.857 | 0.138 | 2.635 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, J.; Pang, L.; Zhang, X.; Lu, X.; Yin, L.; Lu, G.; Cheng, J. Early Discrimination and Prediction of C. fimbriata-Infected Sweetpotatoes during the Asymptomatic Period Using Electronic Nose. Foods 2022, 11, 1919. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11131919
Wu J, Pang L, Zhang X, Lu X, Yin L, Lu G, Cheng J. Early Discrimination and Prediction of C. fimbriata-Infected Sweetpotatoes during the Asymptomatic Period Using Electronic Nose. Foods. 2022; 11(13):1919. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11131919
Chicago/Turabian StyleWu, Jiawen, Linjiang Pang, Xiaoqiong Zhang, Xinghua Lu, Liqing Yin, Guoquan Lu, and Jiyu Cheng. 2022. "Early Discrimination and Prediction of C. fimbriata-Infected Sweetpotatoes during the Asymptomatic Period Using Electronic Nose" Foods 11, no. 13: 1919. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11131919