
Whey Protein Hydrolysates Improved the Oxidative Stability and Water-Holding Capacity of Pork Patties by Reducing Protein Aggregation during Repeated Freeze–Thaw Cycles

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. Preparation of WPH
2.3. Preparation of Pork Patties
2.4. Lipid Peroxide Value
2.5. Extraction of Myofibrillar Protein (MP)
2.6. Total Sulfhydryl (SH) Content
2.7. Surface Hydrophobicity
2.8. Zeta Potential
2.9. Particle Size
2.10. Dynamic Rheological Properties
2.11. Pressure Loss
2.12. Low-Field Nuclear Magnetic Resonance (NMR) Analysis
2.13. Statistical Analysis
3. Results and Discussion
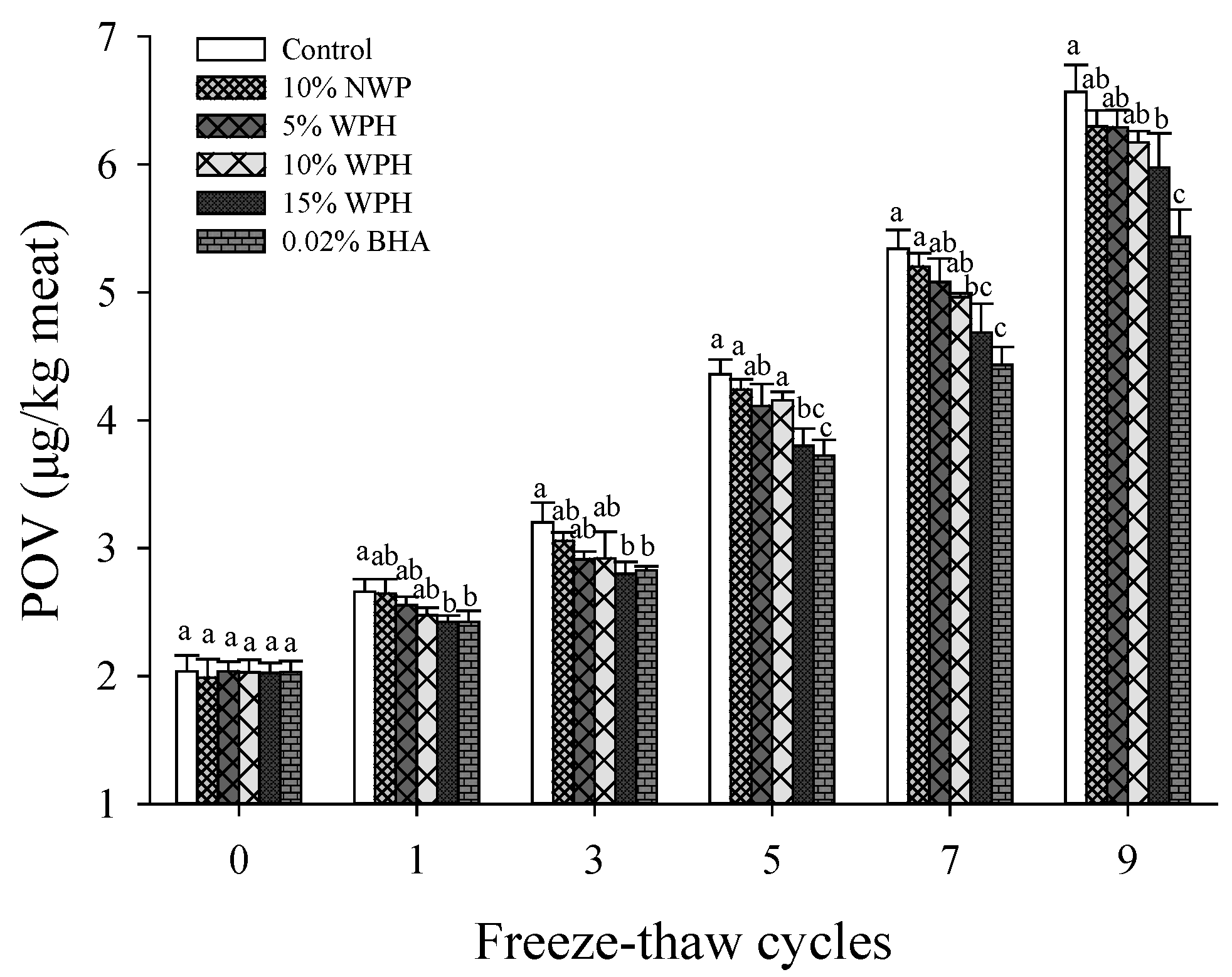
3.1. Lipid Peroxide Value
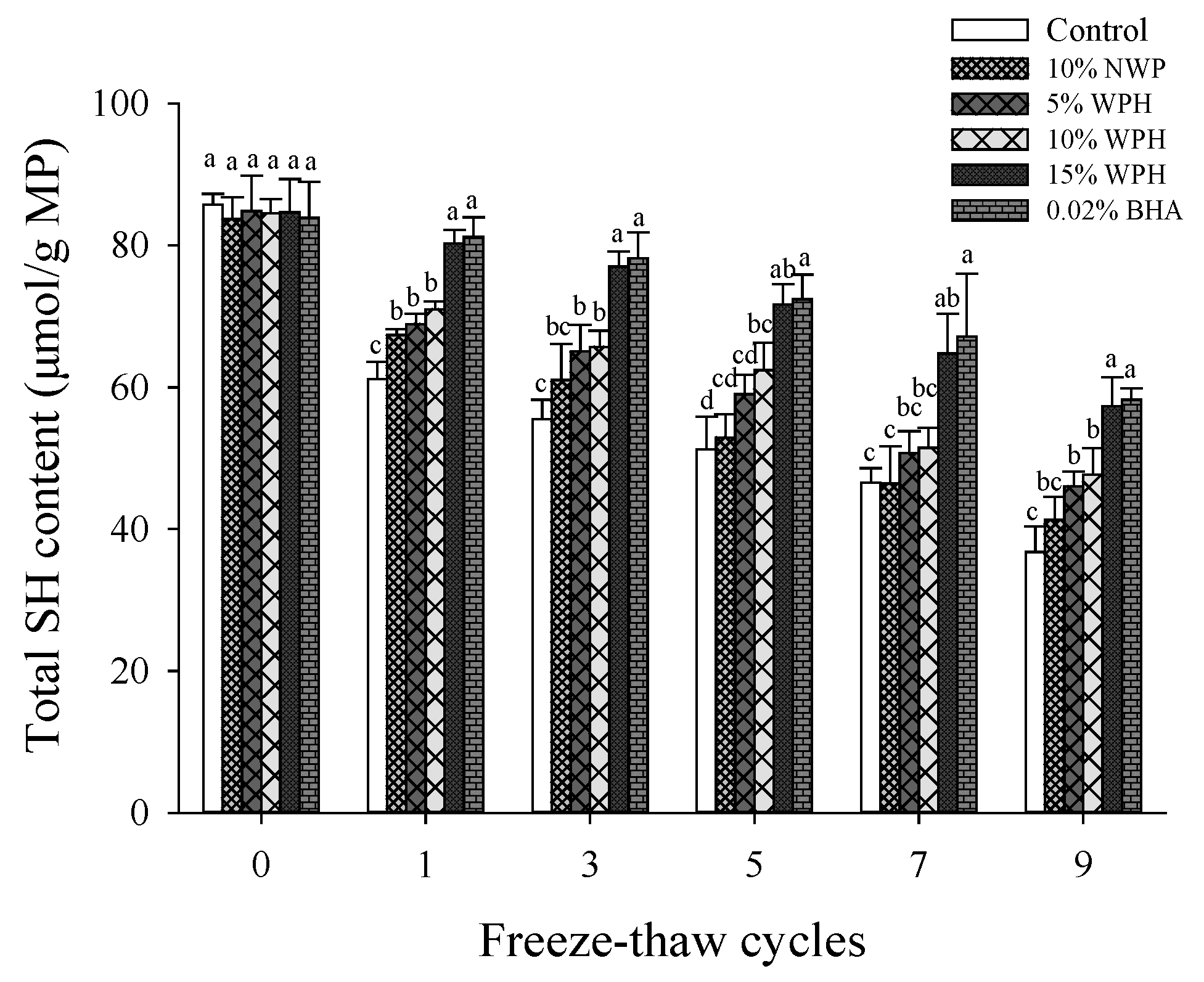
3.2. Total Sulfhydryl Content
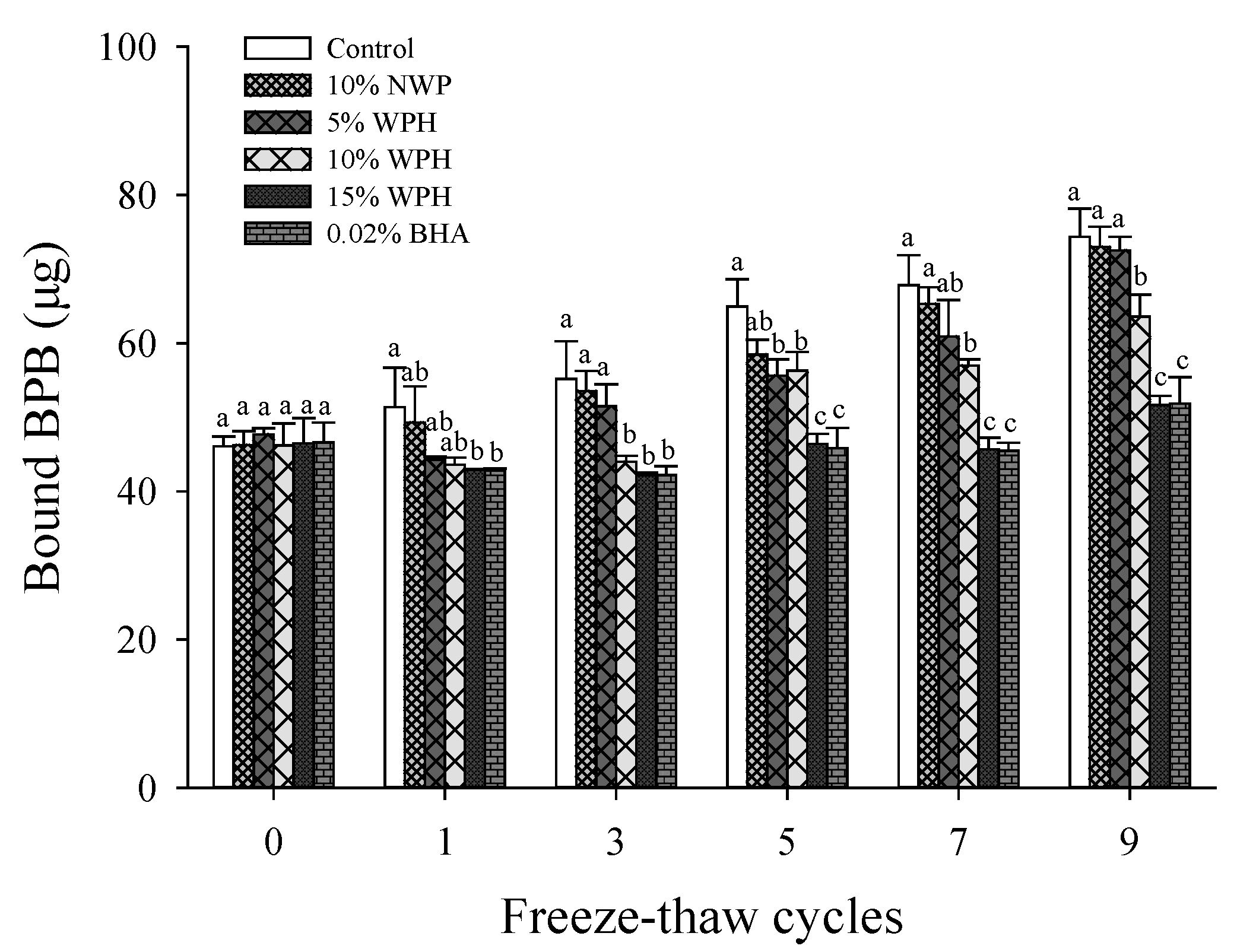
3.3. Surface Hydrophobicity
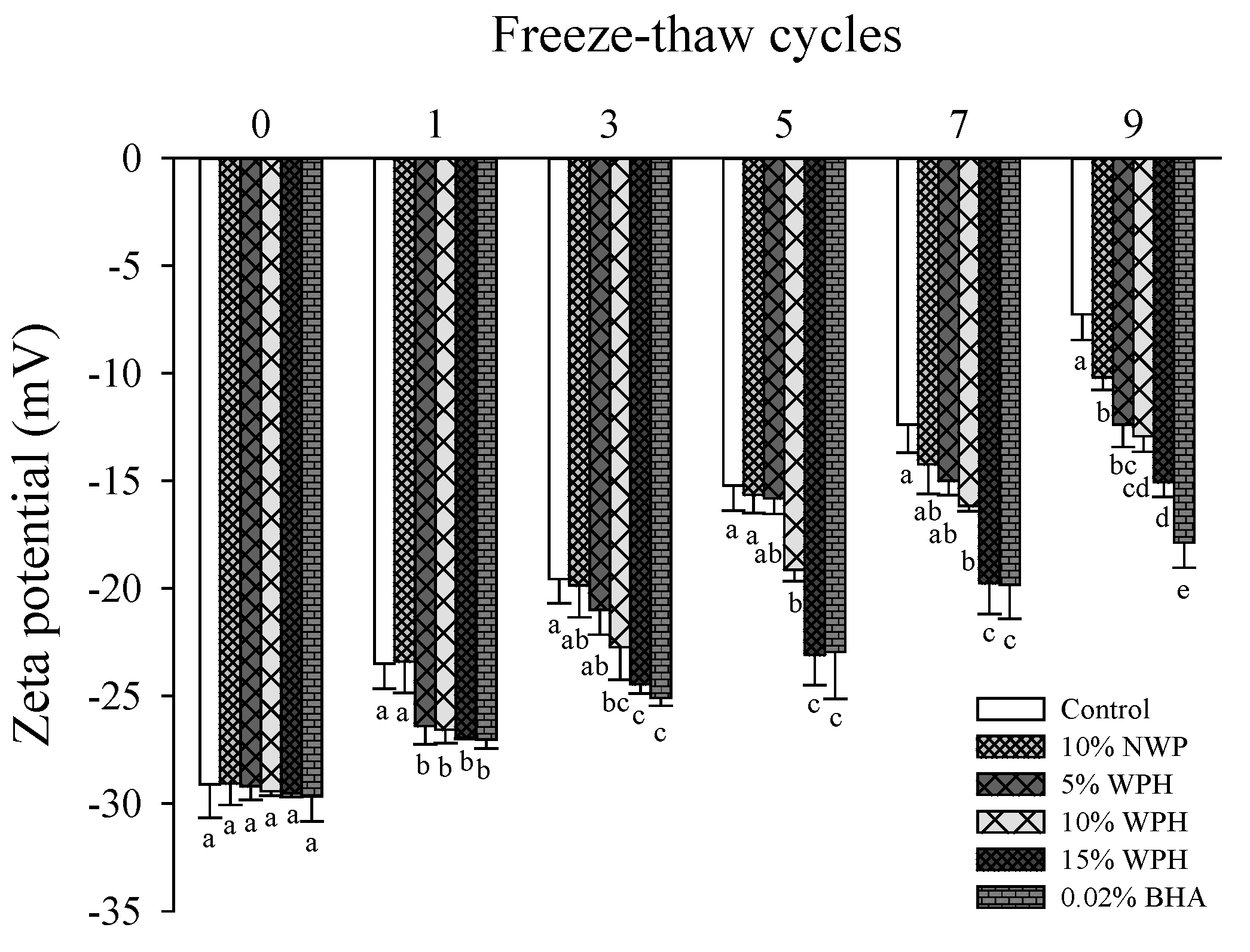
3.4. Zeta Potential
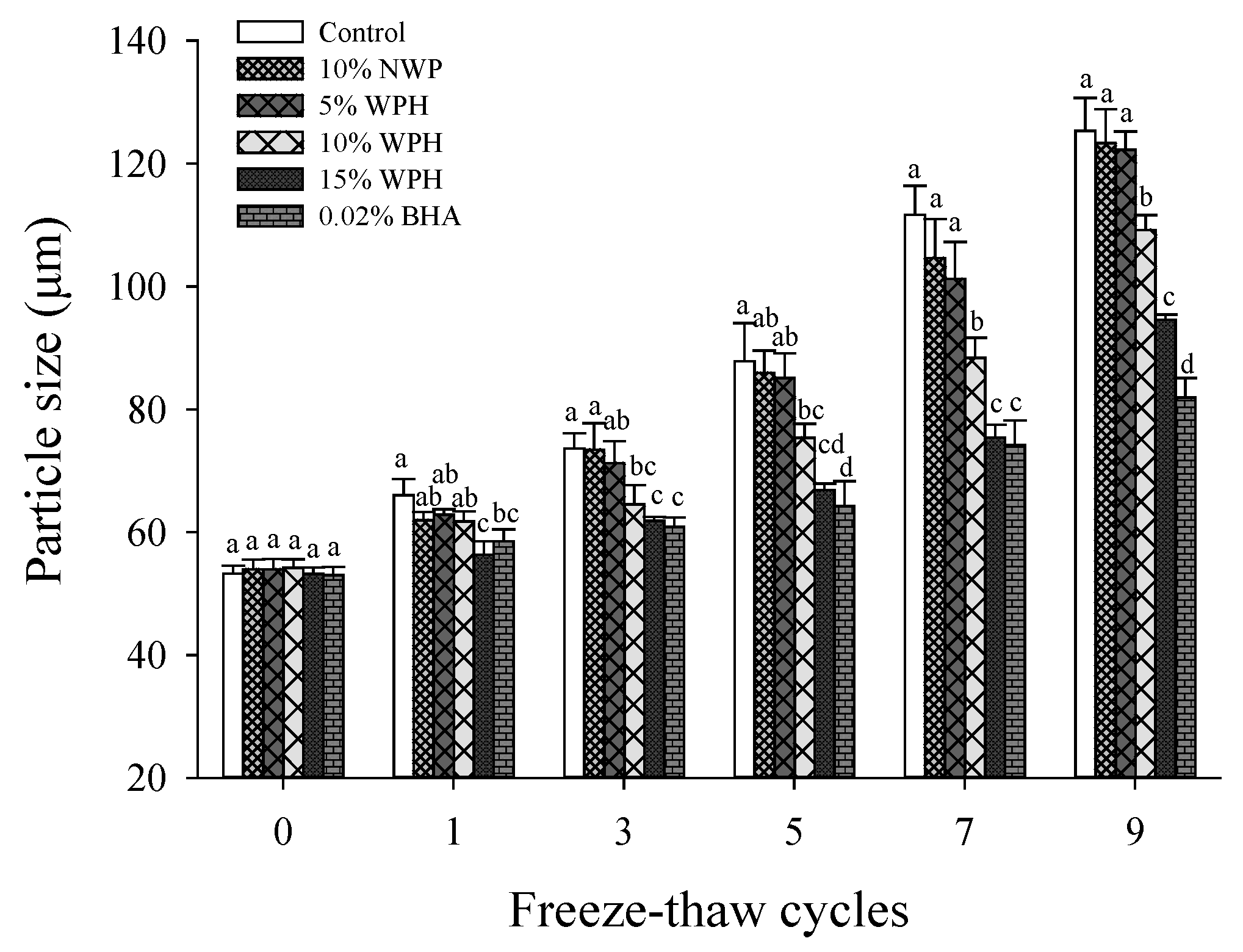
3.5. Particle Size
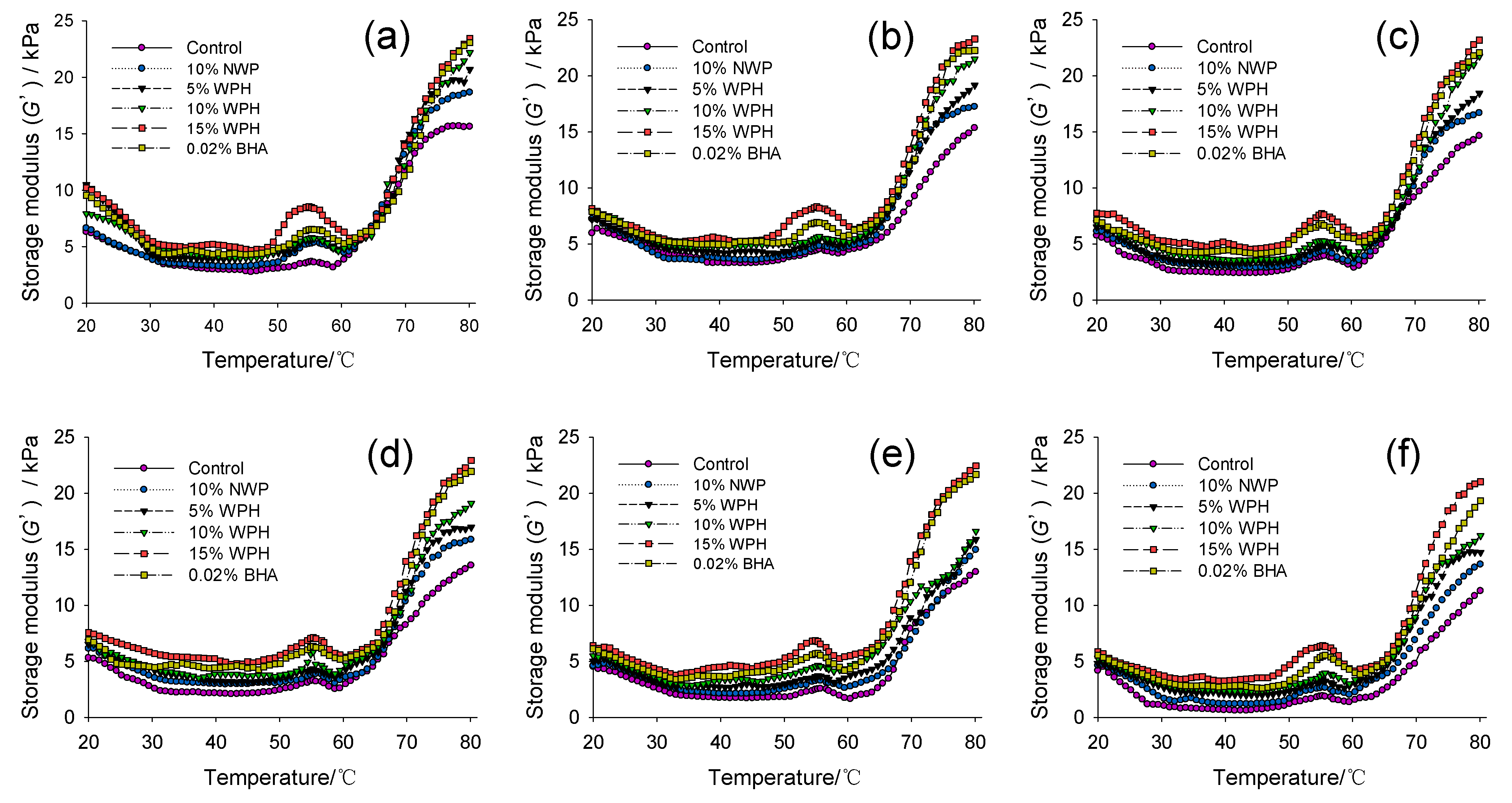
3.6. Rheological Properties
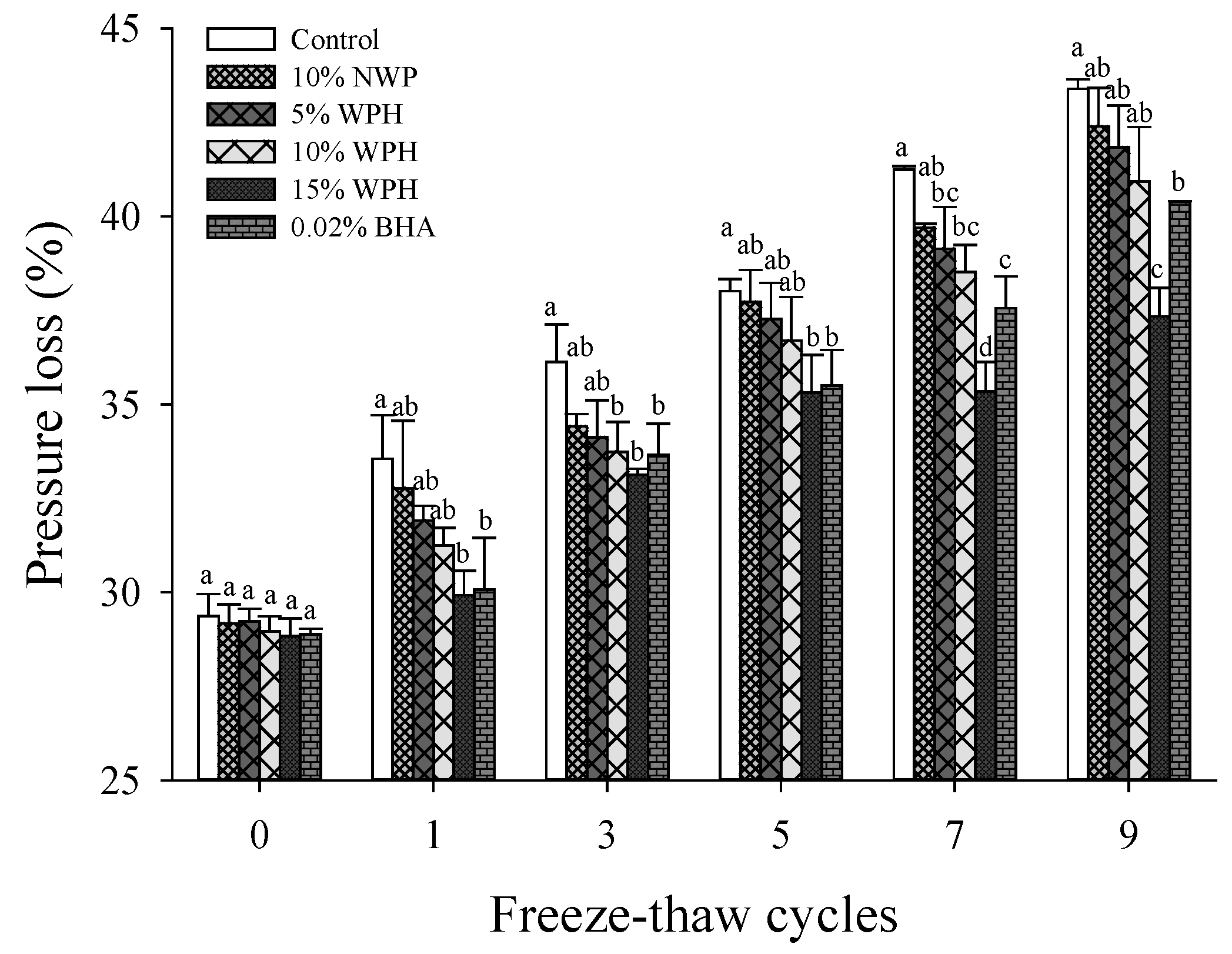
3.7. Pressure Loss
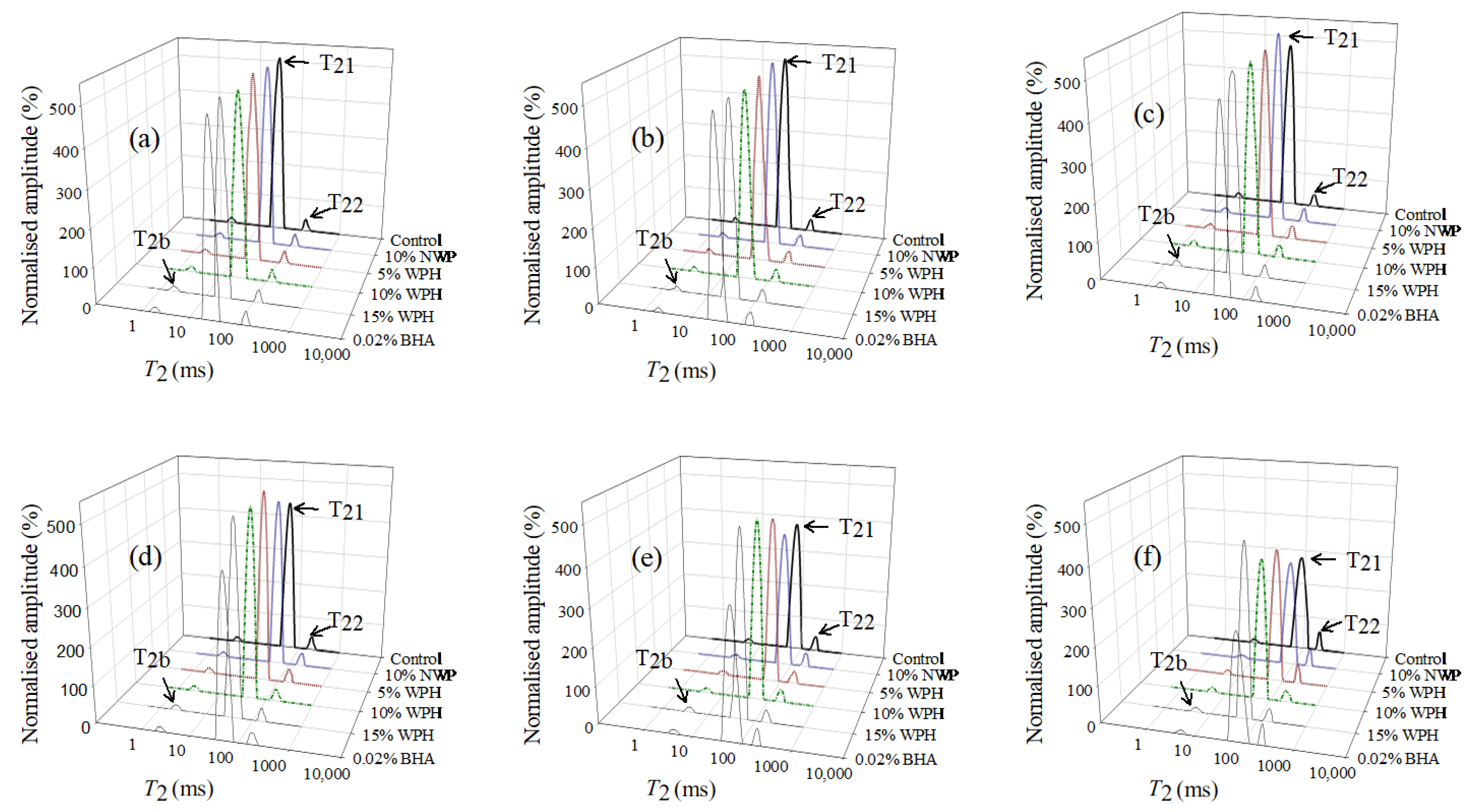
3.8. Low-Field Nuclear Magnetic Resonance Analysis
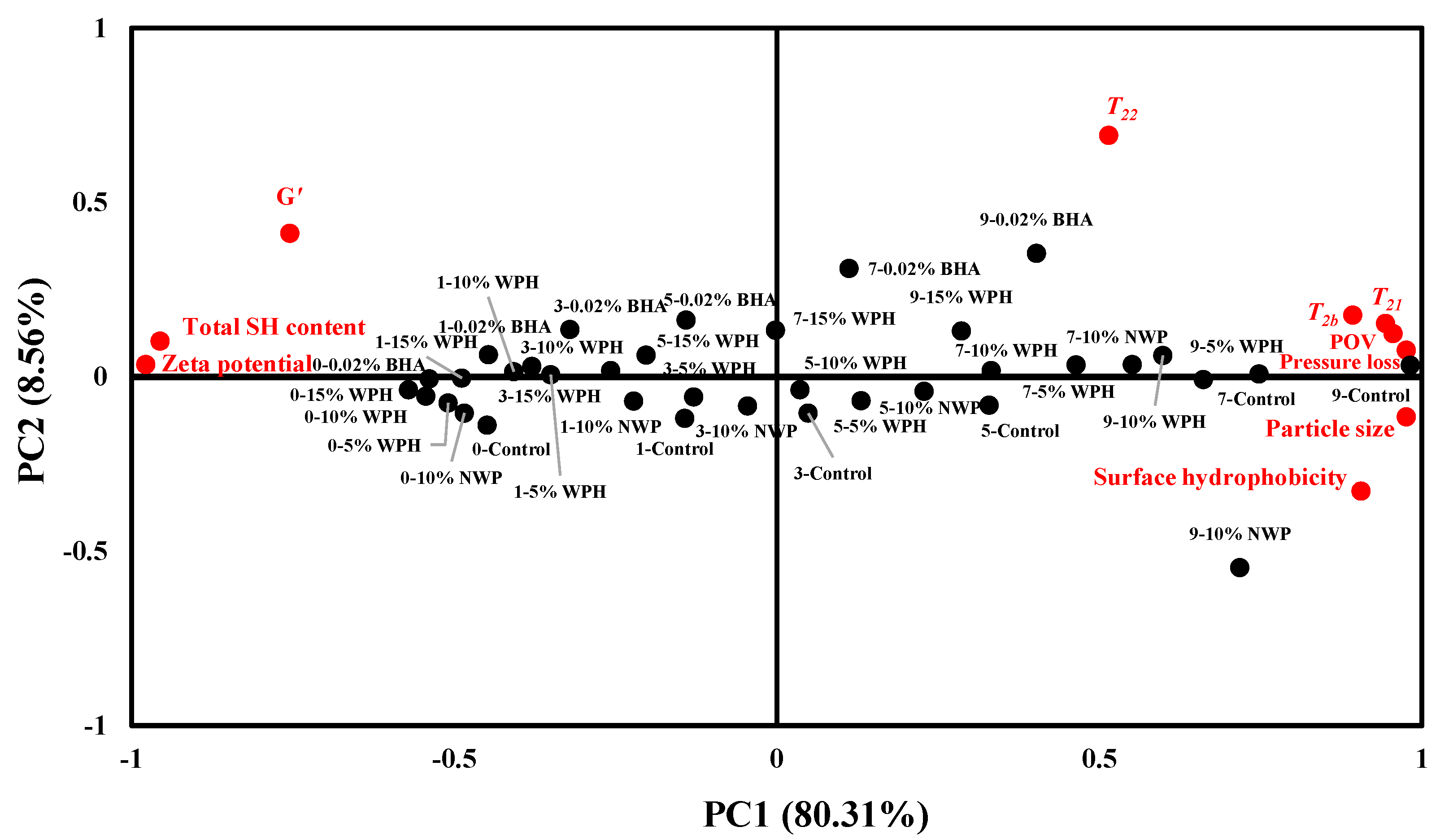
3.9. Principal Component Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Filgueras, R.S.; Gatellier, P.; Zambiazi, R.C.; Santé-Lhoutellier, V. Effect of frozen storage duration and cooking on physical and oxidative changes in M. Gastrocnemius pars interna and M. Iliofiburalis of Rhea americana. Meat Sci. 2011, 88, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, R.S.B.; Francisco, C.L.; Lino, D.M.; Borba, H. Meat quality of Santa Inês lamb chilled-then-frozen storage up to 12 months. Meat Sci. 2019, 148, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, F.; Diao, X.; Kong, B.; Xia, X. Moisture migration, microstructure damage and protein structure changes in porcine longissimus muscle as influenced by multiple freeze-thaw cycles. Meat Sci. 2017, 133, 10–18. [Google Scholar] [CrossRef]
- Leygonie, C.; Britz, T.J.; Hoffman, L.C. Impact of freezing and thawing on the quality of meat: Review. Meat Sci. 2012, 91, 93–98. [Google Scholar] [CrossRef]
- Zhang, B.; Fang, C.; Hao, G.; Zhang, Y. Effect of kappa-carrageenan oligosaccharides on myofibrillar protein oxidation in peeled shrimp (Litopenaeus vannamei) during long-term frozen storage. Food Chem. 2018, 245, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Arash, S.M.; Piret, R.; Dagmar, A.B.; Ralf, L.; Stephan, D. Dynamic rheology, microstructure and texture properties of model porcine meat batter as affected by different cold-set binding systems. Food Hydrocoll. 2018, 77, 937–944. [Google Scholar] [CrossRef]
- Huang, L.; Xiong, Y.L.; Kong, B.; Huang, X.; Li, J. Influence of storage temperature and duration on lipid and protein oxidation and flavour changes in frozen pork dumpling filler. Meat Sci. 2013, 95, 295–301. [Google Scholar] [CrossRef]
- Cai, L.; Wu, X.; Li, X.; Zhong, K.; Li, Y.; Li, J. Effects of different freezing treatments on physicochemical responses and microbial characteristics of Japanese sea bass (Lateolabrax japonicas) fillets during refrigerated storage. LWT Food Sci. Technol. 2014, 59, 122–129. [Google Scholar] [CrossRef]
- Wen, R.; Hu, Y.; Zhang, L.; Wang, Y.; Chen, Q.; Kong, B. Effect of NaCl substitutes on lipid and protein oxidation and flavor development of Harbin dry sausage. Meat Sci. 2019, 156, 33–43. [Google Scholar] [CrossRef]
- Utrera, M.; Morcuende, D.; Estévez, M. Temperature of frozen storage affects the nature and consequences of protein oxidation in beef patties. Meat Sci. 2014, 96, 1250–1257. [Google Scholar] [CrossRef]
- Zhang, Y.; Ertbjerg, P. Effects of frozen-then-chilled storage on proteolytic enzyme activity and water-holding capacity of pork loin. Meat Sci. 2018, 145, 375–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Xiong, S.; You, J.; Hu, Y.; Huang, Q.; Yin, T. Effects of vacuum chopping on physicochemical and gelation properties of myofibrillar proteins from silver carp (Hypophthalmichthys molitrix). Food Chem. 2018, 245, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Huang, F.; Huang, M.; Zhou, G. Influence of oxidation on myofibrillar proteins degradation from bovine via l-calpain. Food Chem. 2012, 134, 106–112. [Google Scholar] [CrossRef]
- Li, F.; Wang, B.; Liu, Q.; Chen, Q.; Zhang, H.; Xia, X.; Kong, B. Changes in myofibrillar protein gel quality of porcine longissimus muscle induced by its stuctural modification under different thawing methods. Meat Sci. 2019, 147, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Sun, N.; Li, D.; Cheng, S.; Song, L.; Lin, S. Enzyme-controlled hygroscopicity and proton dynamics in sea cucumber (Stichopus japonicus) ovum peptide powders. Food Res. Int. 2018, 112, 241–249. [Google Scholar] [CrossRef]
- Bao, Y.; Boeren, S.; Ertbjerg, P. Myofibrillar protein oxidation affects filament charges, aggregation and water-holding. Meat Sci. 2018, 135, 102–108. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Bai, X.; Du, X.; Pan, N.; Shi, S.; Xia, X. Comparison of Effects from Ultrasound Thawing, Vacuum Thawing and Microwave Thawing on the Quality Properties and Oxidation of Porcine Longissimus Lumborum. Foods 2022, 11, 1368. [Google Scholar] [CrossRef]
- Cheng, W.; Sun, D.W.; Pu, H.; Wei, Q. Heterospectral two-dimensional correlation analysis with near-infrared hyperspectral imaging for monitoring oxidative damage of pork myofibrils during frozen storage. Food Chem. 2018, 248, 119–127. [Google Scholar] [CrossRef]
- Lin, S.; Yang, S.; Li, X.; Chen, F.; Zhang, M. Dynamics of water mobility and distribution in soybean antioxidant peptide powders monitored by LF-NMR. Food Chem. 2016, 199, 280–286. [Google Scholar] [CrossRef]
- Wang, L.L.; Xiong, Y.L. Inhibition of oxidant-induced biochemical changes of pork myofibrillar protein by hydrolyzed potato protein. J. Food Sci. 2008, 73, 482–487. [Google Scholar] [CrossRef]
- Cheng, Y.; Xiong, Y.L.; Chen, J. Antioxidant and emulsifying properties of potato protein hydrolysate in soybean oil-in-water emulsions. Food Chem. 2010, 120, 101–108. [Google Scholar] [CrossRef]
- Li, Y.; Kong, B.; Xia, X.; Liu, Q.; Diao, X. Structural changes of the myofibrillar proteins in common carp (Cyprinus carpio) muscle exposed to a hydroxyl radical-generating system. Process Biochem. 2013, 48, 863–870. [Google Scholar] [CrossRef]
- Nikoo, M.; Benjakul, S.; Xu, X. Antioxidant and cryoprotective effects of Amur sturgeon skin gelatin hydrolysate in unwashed fish mince. Food Chem. 2015, 181, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Xiong, Y.L.; Kong, B. Antioxidant activity of peptide fractions from whey protein hydrolysates as measured by electron spin resonance. Food Chem. 2009, 113, 196–201. [Google Scholar] [CrossRef]
- Wang, Z.; He, Z.; Gan, X.; Li, H. Interrelationship among ferrous myoglobin, lipid and protein oxidations in rabbit meat during refrigerated and superchilled storage. Meat Sci. 2018, 146, 131–139. [Google Scholar] [CrossRef]
- Xue, S.; Xu, X.; Shan, H.; Wang, H.; Yang, J.; Zhou, G. Effects of high-intensity ultrasound, high-pressure processing, and high-pressure homogenization on the physicochemical and functional properties of myofibrillar proteins. Innov. Food Sci. Emerg. 2018, 45, 354–360. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, Y.; Li, P.; Wang, X.; Cai, K.; Chen, C. Combined effect of CaCl2 and high pressure processing on the solubility of chicken breast myofibrillar proteins under sodium-reduced conditions. Food Chem. 2018, 269, 236–243. [Google Scholar] [CrossRef]
- Zhao, X.; Bai, Y.; Xing, T.; Xu, X.; Zhou, G. Use of an isoelectric solubilization/precipitation process to modify the functional properties of PSE (pale, soft, exudative)-like chicken meat protein: A mechanistic approach. Food Chem. 2018, 248, 201–209. [Google Scholar] [CrossRef]
- Farouk, M.M.; Wieliczko, K.J. Effect of diet and fat content on the functional properties of thawed beef. Meat Sci. 2003, 64, 451–458. [Google Scholar] [CrossRef]
- Han, M.; Wang, P.; Xu, X.; Zhou, G. Low-field NMR study of heat-induced gelation of pork myofibrillar proteins and its relationship with microstructural characteristics. Food Res. Int. 2014, 62, 1175–1182. [Google Scholar] [CrossRef]
- Mir, S.A.; Masoodi, F.A.; Raja, J. Influence of natural antioxidants on microbial load, lipid oxidation and sensorial quality of rista—A traditional meat product of India. Food Biosci. 2017, 20, 79–87. [Google Scholar] [CrossRef]
- Chen, Q.; Xie, Y.; Xi, J.; Guo, Y.; Qian, H.; Cheng, Y.; Chen, Y.; Yao, W. Characterization of lipid oxidation process of beef during repeated freeze-thaw by electron spin resonance technology and Raman spectroscopy. Food Chem. 2018, 243, 58–64. [Google Scholar] [CrossRef]
- Du, X.; Zhao, M.N.; Pan, N.; Wang, S.P.; Xia, X.F.; Zhang, D.J. Tracking aggregation behaviour and gel properties induced by structural alterations in myofibrillar protein in mirror carp (Cyprinus carpio) under the synergistic effects of pH and heating. Food Chem. 2021, 362, 130222. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Pan, S.; Wang, H. Effect of trehalose on Lateolabrax japonicus myofibrillar protein during frozen storage. Food Chem. 2014, 160, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Sisse, J.; Tørngren, M.A.; Gunvig, A.; Skibsted, L.H.; Lund, M.N. Effect of green tea or rosemary extract on protein oxidation in Bologna type sausages prepared from oxidatively stressed pork. Meat Sci. 2013, 93, 538–546. [Google Scholar] [CrossRef]
- Wang, X.; Muhoza, B.; Wang, X.; Feng, T.; Xia, S.; Zhang, X. Comparison between microwave and traditional water bath cooking on saltiness perception, water distribution and microstructure of grass crap meat. Food Res. Int. 2019, 125, 108521. [Google Scholar] [CrossRef]
- Zhao, J.; Lv, W.; Wang, J.; Li, J.; Liu, X.; Zhu, J. Effects of tea polyphenols on the post-mortem integrity of large yellow croaker (Pseudsciaena crocea) fillet proteins. Food Chem. 2013, 141, 2666–2674. [Google Scholar] [CrossRef]
- Hwang, K.E.; Choi, Y.S.; Choi, S.M.; Kim, H.W.; Choi, J.H.; Lee, M.A.; Kim, C. Antioxidant action of ganghwayakssuk (Artemisia princeps Pamp.) in combination with ascorbic acid to increase the shelf life in raw and deep fried chicken nuggets. Meat Sci. 2013, 95, 593–602. [Google Scholar] [CrossRef]
- Liu, Q.; Kong, B.; Han, J.; Chen, Q.; He, X. Effects of superchilling and cryoprotectants on the quality of common carp (Cyprinus carpio) surimi: Microbial growth, oxidation, and physiochemical properties. LWT Food Sci. Technol. 2014, 57, 165–171. [Google Scholar] [CrossRef]
- Zhang, T.; Xu, X.; Li, Z.; Wang, Y.; Xue, Y.; Xue, C. Interactions and phase behaviors in mixed solutions of κ-carrageenan and myofibrillar protein extracted from Alaska Pollock surimi. Food Res. Int. 2018, 105, 821–827. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, Y.; Zhou, P.; Zhang, X.; Wang, J. Effects of high pressure modification on conformation and gelation properties of myofibrillar protein. Food Chem. 2017, 217, 678–686. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Zhou, C.; Fu, F.; Chen, Z.; Wu, Q. Effect of high-pressure homogenization on particle size and film properties of soy protein isolate. Ind. Crops Prod. 2013, 43, 538–544. [Google Scholar] [CrossRef]
- Soleimanpour, M.; Koocheki, A.; Kadkhodaee, R. Effect of Lepidium perfoliatum seed gum addition on whey protein concentrate stabilized emulsions stored at cold and ambient temperature. Food Hydrocoll. 2013, 30, 292–301. [Google Scholar] [CrossRef]
- Pan, N.; Wan, W.; Du, X.; Kong, B.; Liu, Q.; Lv, H.; Xia, X.; Li, F. Mechanisms of change in emulsifying capacity induced by protein denaturation and aggregation in quick-frozen pork patties with different fat levels and freeze-thaw cycles. Foods 2022, 11, 44. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Zhao, M.; Sun, W. Effect of pH on the interaction of porcine myofibrillar proteins with pyrazine compounds. Food Chem. 2019, 287, 93–99. [Google Scholar] [CrossRef]
- Zhuang, X.; Jiang, X.; Zhou, H.; Han, M.; Liu, Y.; Bai, Y.; Xu, X.; Zhou, G. The effect of insoluble dietary fiber on myofibrillar protein emulsion gels: Oil particle size and protein network microstructure. LWT Food Sci. Technol. 2019, 101, 534–542. [Google Scholar] [CrossRef]
- Gani, A.; Benjakul, S. Impact of virgin coconut oil nanoemulsion on properties of croaker surimi gel. Food Hydrocoll. 2018, 82, 34–44. [Google Scholar] [CrossRef]
- Gharibzahedi, S.M.T.; Roohinejad, S.; George, S.; Barba, F.J.; Greiner, R.; Barbosa-Cánovas, G.V.; Mallikarjunan, K. Innovative food processing technologies on the transglutaminase functionality in protein-based food products: Trends, opportunities and drawbacks. Trends Food Sci Technol. 2018, 75, 194–205. [Google Scholar] [CrossRef]
- Tornberg, E. Effects of heat on meat proteins—Implications on structure and quality of meat products. Meat Sci. 2005, 70, 493–508. [Google Scholar] [CrossRef]
- Kang, Z.; Hu, S.; Zhu, D.; Ma, H. Effect of Sodium Chloride and Processing Methods on Protein Aggregation, Physical-Chemical and Rheological Properties of Pork Batters. Int. J. Food Eng. 2018, 14, 5–6. [Google Scholar] [CrossRef]
- Zhao, S.; Liu, Y.; Yuan, X.; Zhao, Y.; Kang, Z.; Zhu, M.; Ma, H. Effect of low-frequency alternating magnetic field on the rheological properties, water distribution and microstructure of low-salt pork batters. LWT 2022, 159, 113164. [Google Scholar] [CrossRef]
- Huang, L.; Ding, B.; Zhang, H.; Kong, B.; Xiong, Y. Textural and sensorial quality protection in frozen dumplings through the inhibition of lipid and protein oxidation with clove and rosemary extracts. J. Sci. Food Agric. 2019, 99, 4739–4747. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, Y.; Chen, L.; Xu, X.; Zhou, G.; Li, Z.; Feng, X. Dose-dependent effects of rosmarinic acid on formation of oxidatively stressed myofibrillar protein emulsion gel at different NaCl concentrations. Food Chem. 2018, 243, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Ngapo, T.M.; Babare, I.H.; Reynolds, J.; Mawson, R.F. Freezing and thawing rate effects on drip loss from samples of pork. Meat Sci. 1999, 53, 149–158. [Google Scholar] [CrossRef]
- Gonçalves, E.M.; Abreu, M.; Brandão, T.R.S.; Silva, C.L.M. Degradation kinetics of colour, vitamin C and drip loss in frozen broccoli (Brassica oleracea L. ssp. Italica) during storage at isothermal and non-isothermal conditions. Int. J. Refrig. 2011, 34, 2136–2144. [Google Scholar] [CrossRef]
- Cheng, S.; Wang, X.; Li, R.; Yang, H.; Wang, H.; Wang, H.; Tan, M. Influence of multiple freeze-thaw cycles on quality characteristics of beef semimembranous muscle: With emphasis on water status and distribution by LF-NMR and MRI. Meat Sci. 2019, 147, 44–52. [Google Scholar] [CrossRef]
- Pan, N.; Dong, C.; Du, X.; Kong, B.; Sun, J.; Xia, X. Effect of freeze-thaw cycles on the quality of quick-frozen pork patty with different fat content by consumer assessment and instrument-based detection. Meat Sci. 2021, 172, 108313. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; Kong, L.; Yu, P.; Wen, R.; Yu, X.; Xu, X.; Peng, X. Whey Protein Hydrolysates Improved the Oxidative Stability and Water-Holding Capacity of Pork Patties by Reducing Protein Aggregation during Repeated Freeze–Thaw Cycles. Foods 2022, 11, 2133. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11142133
Liu C, Kong L, Yu P, Wen R, Yu X, Xu X, Peng X. Whey Protein Hydrolysates Improved the Oxidative Stability and Water-Holding Capacity of Pork Patties by Reducing Protein Aggregation during Repeated Freeze–Thaw Cycles. Foods. 2022; 11(14):2133. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11142133
Chicago/Turabian StyleLiu, Chunyun, Lingru Kong, Pengjuan Yu, Rongxin Wen, Xiaobo Yu, Xinglian Xu, and Xinyan Peng. 2022. "Whey Protein Hydrolysates Improved the Oxidative Stability and Water-Holding Capacity of Pork Patties by Reducing Protein Aggregation during Repeated Freeze–Thaw Cycles" Foods 11, no. 14: 2133. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11142133