
Does Finishing at Pasture Influence the Colour of Muscle from Suckler Bulls and Can Colour Be Used to Authenticate Their Pre-Slaughter Diet?

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Management and Post-Slaughter Measurements and Sampling
2.2. Colour and Chemical Analysis
2.3. Statistical Analysis
3. Results
3.1. Animal Growth and Carcass Characteristics
3.2. Muscle Ultimate pH and Colour
3.3. Muscle Composition
3.4. Correlations between Carcass or Muscle Characteristics and Muscle Colour
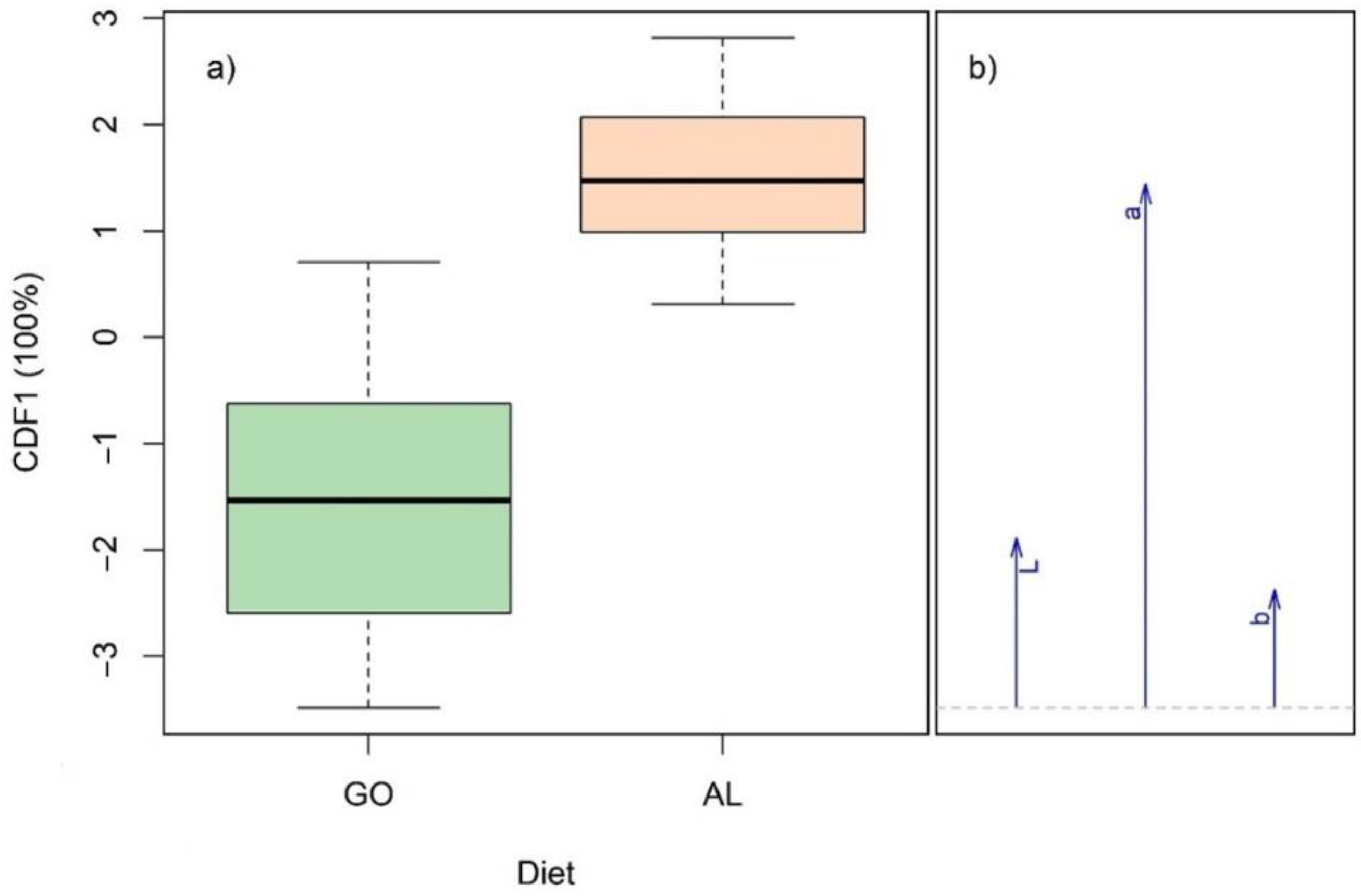
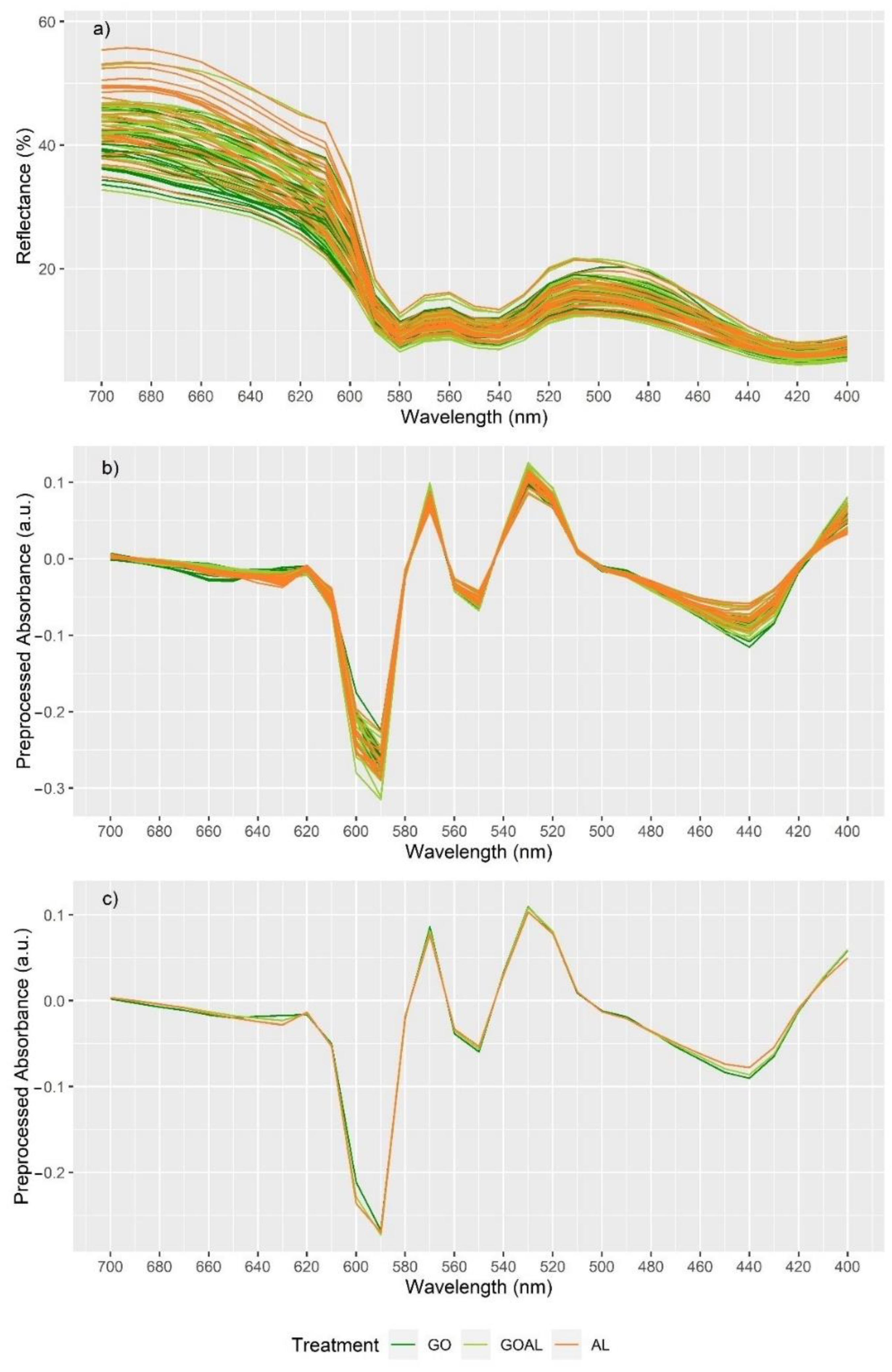
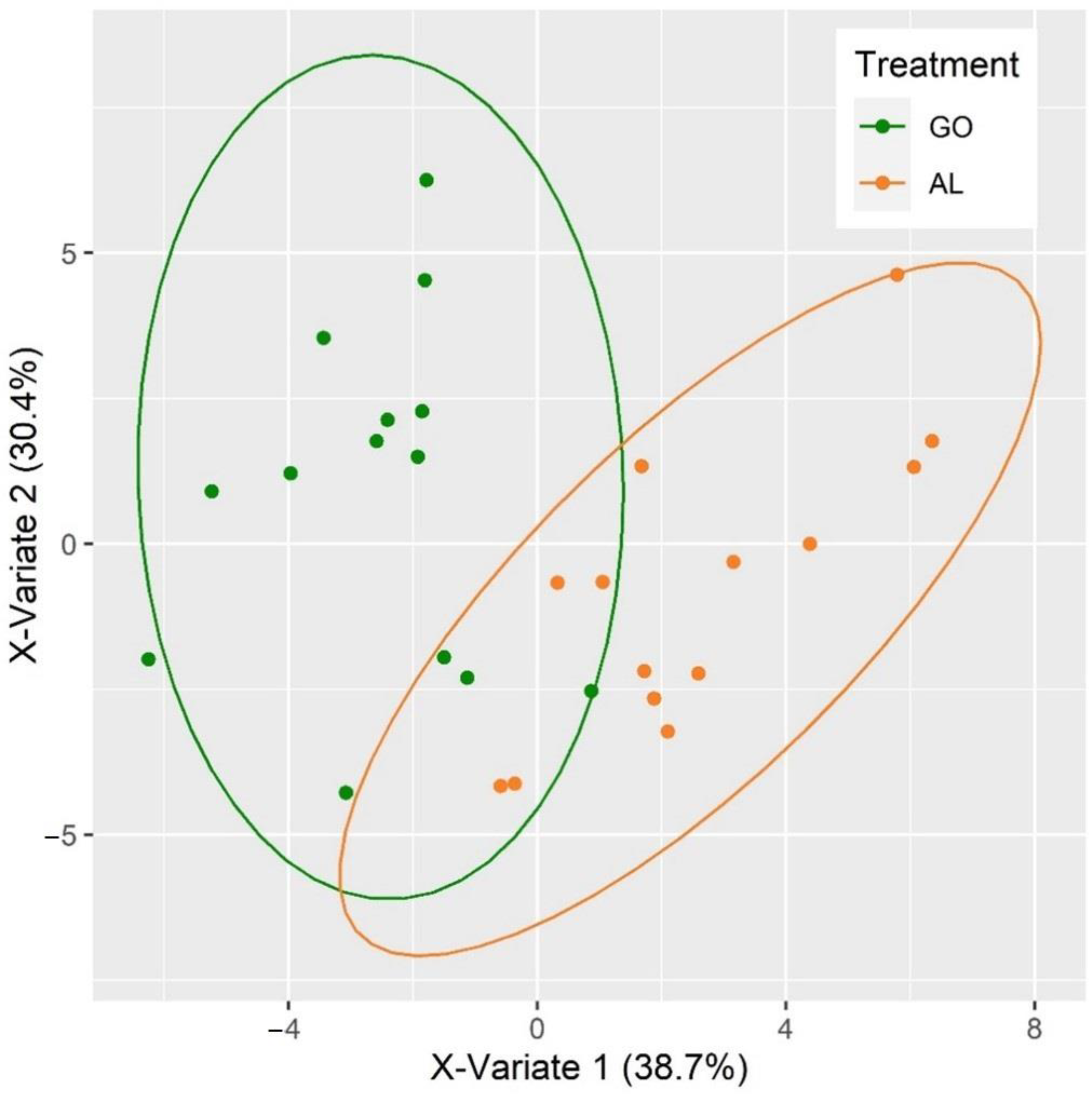
3.5. Classification of Beef According to Pre-Slaughter Cattle Diet
4. Discussion
4.1. Animal Growth and Carcass Characteristics
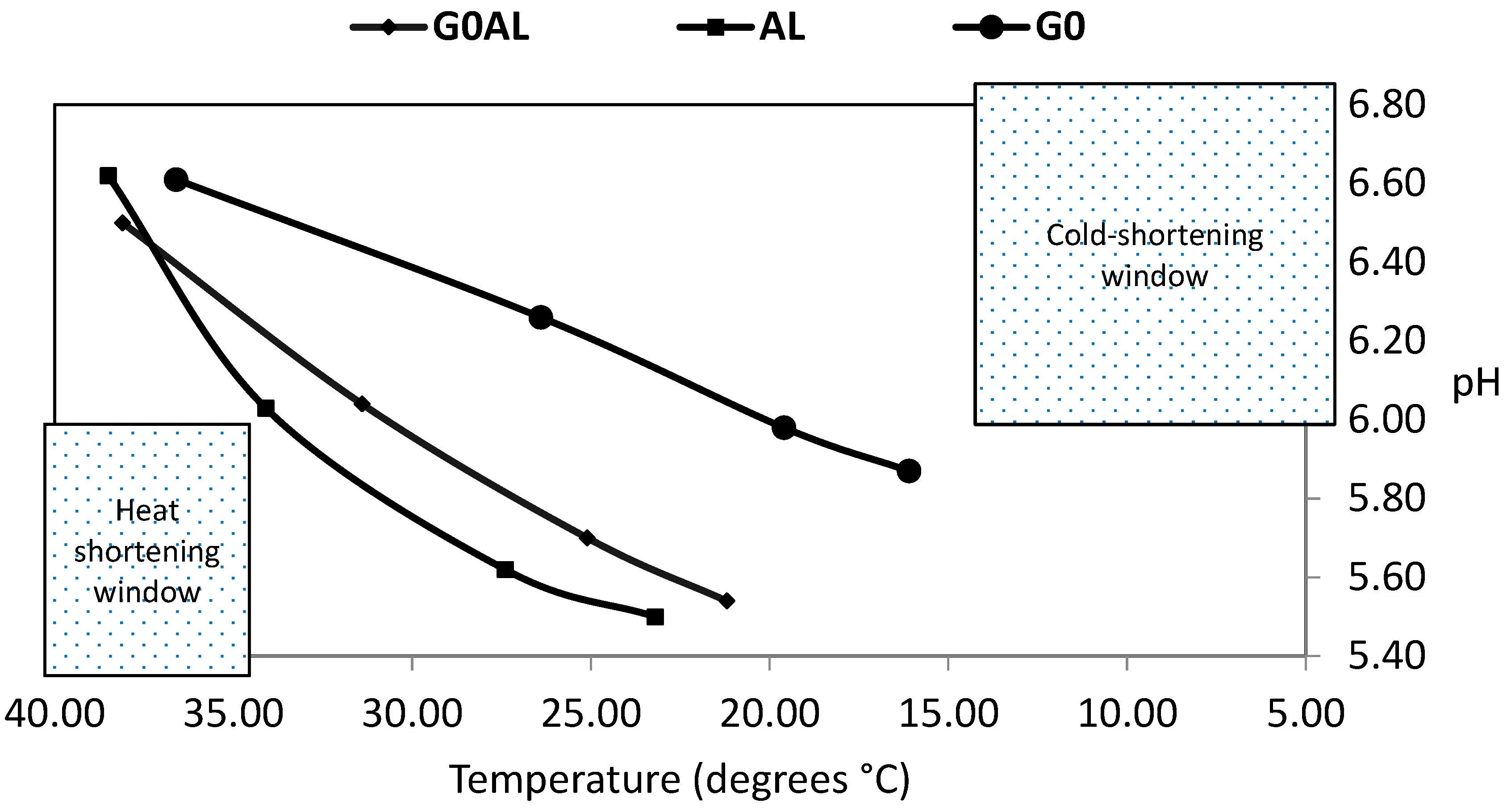
4.2. Ultimate pH
4.3. Muscle Colour
4.4. Possible Explanations for the Differences in Muscle Colour
4.5. Discrimination of Pre-Slaughter Diet of Bulls
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holman, B.W.B.; van de Ven, R.J.; Mao, Y.; Coombs, C.E.O.; Hopkins, D.L. Using instrumental (CIE and reflectance) measures to predict consumers’ acceptance of beef colour. Meat Sci. 2017, 127, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Dunne, P.G.; Monahan, F.J.; Moloney, A.P. Current perspectives on the darker beef often reported from extensively managed cattle: Does physical activity play a significant role. Livest. Sci. 2011, 142, 1–22. [Google Scholar] [CrossRef]
- O’Riordan, E.G.; Crosson, P.; McGee, M. Finishing male cattle from the beef suckler herd. Ir. Grassl. Assoc. J. 2011, 45, 131–146. [Google Scholar]
- Moran, L.; Wilson, S.S.; McElhinney, C.K.; Monahan, F.J.; McGee, M.; O’Sullivan, M.G.; O’Riordan, E.G.; Kerry, J.P.; Moloney, A.P. Suckler bulls slaughtered at 15 months of age: Effect of different production systems on the fatty acid profile and selected quality characteristics of Longissimus thoracis. Foods 2019, 8, 264. [Google Scholar] [CrossRef] [Green Version]
- Monahan, F.J.; Schmidt, O.; Moloney, A.P. Meat provenance: Authentication of geographical origin and dietary background of meat. Meat Sci. 2018, 144, 2–14. [Google Scholar] [CrossRef]
- Huang, Y.; Andueza, D.; de Oliveira, L.; Zawadzki, F.; Prache, S. Visible spectroscopy on carcass fat combined with chemometrics to distinguish pasture-fed, concentrate-fed and concentrate-finished pasture-fed lambs. Meat Sci. 2015, 101, 5–12. [Google Scholar] [CrossRef]
- Prieto, N.; Lopez-Campos, O.; Zijlstra, R.T.; Uttaro, B.; Aalhus, J.L. Discrimination of beef dark cutters using visible and near infrared reflectance spectroscopy. Can. J. Anim. Sci. 2014, 94, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Barragan, W.; Aalhus, J.L.; Penner, G.; Dugan, M.E.R.; Juarez, M.; Lopez-Campos, O.; Vahmani, P.J.; Segura, J.; Angulo, J.; Prieto, N. Authentication of barley finished beef using visible and near infrared spectroscopy (Vis-NIRS) and different discrimination approaches. Meat Sci. 2021, 172, 108342. [Google Scholar] [CrossRef]
- Lenehan, C.; Moloney, A.P.; O’Riordan, E.G.; Kelly, A.; McGee, M. Pasture-based finishing of early-maturing sired suckler beef bulls at 15 or 19 months of age. Adv. Anim. Biosci. 2017, 8, s28–s32. [Google Scholar] [CrossRef]
- Kenny, D.; Murphy, C.P.; Sleator, R.D.; Judge, M.M.; Evans, R.D.; Berry, D.P. Animal-level factors associated with the achievement of desirable specifications in Irish beef carcasses graded using the EUROP classification system. J. Anim. Sci. 2020, 98, 1–12. [Google Scholar] [CrossRef]
- Moran, L.; O’Sullivan, M.G.; Kerry, J.P.; Picard, B.; McGee, M.; O’Riordan, E.G.; Moloney, A.P. Effect of a grazing period prior to finishing on a high concentrate diet on meat quality from bulls and steers. Meat Sci. 2017, 125, 76–83. [Google Scholar] [CrossRef]
- Jurie, C.; Ortigues-Marty, I.; Picard, B.; Micol, D.; Hocquette, J.-F. The separate effects of the nature of diet and grazing mobility on metabolic potential of muscles from Charolais steers. Livest. Sci. 2006, 104, 182–192. [Google Scholar] [CrossRef]
- Picard, B.; Barboiron, C.; Chadeyron, D.; Jurie, C. Protocol for high-resolution electrophoresis separation of myosin heavy chain isoforms in bovine skeletal muscle. Electrophoresis 2011, 32, 1804–1806. [Google Scholar] [CrossRef]
- Krzywicki, K. Assessment of relative content of myoglobin, oxymyoglobin and metmyoglobin at the surface of beef. Meat Sci. 1979, 3, 1–10. [Google Scholar] [CrossRef]
- Hunt, M.C.; King, A.; Barbut, S.; Clause, J.; Cornforth, D.; Hanson, D.; Lindahl, G.; Mancini, R.; Milkowski, A.; Mohan, A.; et al. AMSA Meat Color Measurement Guidelines; American Meat Science Association: Champaign, IL, USA, 2012. [Google Scholar]
- Krzywicki, K. The determination of haem pigments in meat. Meat Sci. 1982, 7, 29–36. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: http://www.r-project.org/ (accessed on 24 May 2022).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002; ISBN 0-387-95457-0. [Google Scholar]
- Friendly, M.; Fox, J. Candisc: Visualizing Generalized Canonical Discriminant and Canonical Correlation Analysis. R Package Version 0.8-6. 2021. Available online: https://CRAN.R-project.org/package=heplots (accessed on 30 May 2022).
- Rohart, F.; Gautier, B.; Singh, A.; Le Cao, K.-A. mixOmics: An R package for ’omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. Available online: http://www.mixOmics.org (accessed on 24 May 2022).
- Andersen, C.M.; Bro, R. Variable selection in regression—a tutorial. J. Chemom. 2010, 24, 728–737. [Google Scholar] [CrossRef]
- Bord Bia. Grass Fed Beef Standard. 2020. Available online: https://www.bordbia.ie/globalassets/bordbia2020/farmers--growers/grass-fed-standard/grass-fed-beef-standard-revision-02-draft1.pdf (accessed on 1 May 2022).
- Ashmore, C.R.; Carroll, F.; Doerr, L.; Tompkins, G.; Stokes, H.; Parker, W. Experimental prevention of dark-cutting meat. J. Anim. Sci. 1973, 36, 33–36. [Google Scholar] [CrossRef]
- Viljoen, H.; De Kock, H.; Webb, E. Consumer acceptability of dark, firm and dry (DFD) and normal pH beef steaks. Meat Sci. 2002, 61, 181–185. [Google Scholar] [CrossRef]
- Liu, J.; Pogorzelski, G.; Neveu, A.; Legrand, I.; Pethick, D.; Elllies-Oury, M.-P.; Hocquette, J.F. Are marbling and the prediction of beef eating qualty affected by different grading sites? Front. Vet. Med. 2021, 8, 611153. [Google Scholar] [CrossRef]
- Moloney, A.P.; O’ Riordan, E.G.; McGee, M.; O’Sullivan, M.G.; Kerry, J.P.; Wilson, S.S.; Monahan, F.J.; Kelly, A.K.; McMenamin, K.; Moran, L. Carcass characteristics, colour and eating quality of beef from late maturing suckler bulls finished at pasture or indoors on a high concentrate ration. Anim. Prod. Sci. 2022, 62, 590–600. [Google Scholar] [CrossRef]
- Acciaro, M.; Decandia, M.; Giovanetti, V.; Manca, C.; Dimauro, C.; Giacomo Rassu, S.P.; Addis, M.; Molle, G. Effects of grass- and concentrate-based finishing systems on the quality of meat from the M. longissimus thoracis of young Sarda bulls. Anim. Prod. Sci. 2021, 81, 807–820. [Google Scholar] [CrossRef]
- Dunne, P.G.; O’Mara, F.P.; Monahan, F.J.; Moloney, A.P. Colour of subcutaneous adipose tissue and muscle of Irish beef carcasses destined for the Italian market. Ir. J. Agric. Food Res. 2004, 43, 217–226. [Google Scholar]
- Vestergaard, M.; Oksbjerg, N.; Henckel, P. Influence of feeding intensity, grazing and finishing feeding on muscle fibre characteristics and meat colour of semitendinosus, longissimus dorsi and supraspinatus muscles of young bulls. Meat Sci. 2000, 54, 177–185. [Google Scholar] [CrossRef]
- Apaoblaza, A.; Gerrard, S.D.; Matarneh, S.K.; Wicks, J.C.; Kirkpatrick, L.; England, E.M.; Scheffler, T.L.; Duckett, S.K.; Shi, H.; Silva, S.L.; et al. Muscle from grass- and grain-fed cattle differs energetically. Meat Sci. 2020, 161, 107996. [Google Scholar] [CrossRef]
- Moran, L.; Wilson, S.S.; O’Sullivan, M.G.; McGee, M.; O’Riordan, E.G.; Monahan, F.J.; Kerry, J.P.; Moloney, A.P. Quality of three muscles from suckler bulls finished on concentrates and slaughtered at 16 months of age or slaughtered at 19 months of age from two production systems. Animal 2020, 14, 2203–2211. [Google Scholar] [CrossRef]
- Gagaoua, M.; Picard, B.; Monteils, V. Associations among animal, carcass, muscle characteristics, and fresh meat color traits in Charolais cattle. Meat Sci. 2018, 140, 145–156. [Google Scholar] [CrossRef]
- Djimsa, B.A.; Nair, M.N.; Hess, A.M.; Belk, K.E.; Woerner, D.R. The impact of carcass size, chilling conditions, and electrical stimulation on beef post-mortem temperature and pH decline. Meat Muscle Biol. 2021, 6, 13893. [Google Scholar]
- Hughes, J.M.; Clarke, F.M.; Purslow, P.P.; Warner, R.D. Meat colour is determined not only by chromatic heme pigments but also by the physical structure and achromatic light scattering properties of the muscle. Compr. Rev. Food Sci. Food Saf. 2020, 19, 44–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandstetter, A.M.; Picard, B.; Geay, Y. Muscle fibre characteristics in four muscles of growing male cattle: Effect of castration and feeding level. Livest. Prod. Sci. 1998, 53, 25–36. [Google Scholar] [CrossRef]
- Gagaoua, M.; Monteils, V.; Couvreur, S.; Picard, B. Identification of biomarkers associated with the rearing practices, carcass characteristics, and beef quality: An integrative approach. J. Agric. Food Chem. 2017, 65, 8264–8278. [Google Scholar] [CrossRef]
- Picard, B.; Cassar-Malek, I. Evidence for expression of IIb myosin heavy chain isoform in some skeletal muscles of Blonde d’Aquitaine bulls. Meat Sci. 2009, 82, 30–36. [Google Scholar] [CrossRef]
- Meat and Livestock Australia. Meat Standards Australia Beef Information Kit; Meat and Livestock Australia Ltd.: Sydney, Australia, 2017. [Google Scholar]
- Kim, Y.H.B.; Warner, R.D.; Rosenvold, K. Influence of high pre-rigour temperature and fast pH fall on muscle proteins and meat quality: A review. Anim. Prod. Sci. 2014, 54, 375–395. [Google Scholar] [CrossRef] [Green Version]
- Hughes, J.M.; Kearney, G.; Warner, R.D. Improving beef meat colour scores at carcass grading. Anim. Prod. Sci. 2014, 54, 422–429. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | G0 | G0AL | AL | SED | Significance 1 |
|---|---|---|---|---|---|
| Weight (kg) | |||||
| Initial | 435 | 424 | 424 | 13.7 | NS |
| Housing 2 | 545 a | 534 a | 616 b | 16.6 | *** |
| Final | 639 a | 686 b | 741 c | 20.2 | *** |
| Age (days) | 591 | 584 | 570 | 17.4 | NS |
| Growth (kg/day) | |||||
| Turnout to housing | 1.54 a | 1.43 a | 1.96 b | 0.092 | *** |
| Housing to slaughter | 0.95 a | 1.50 b | 1.25 c | 0.114 | *** |
| Overall | 1.18 a | 1.42 b | 1.60 c | 0.081 | *** |
| Carcass weight (kg) | 359 a | 399 b | 436 c | 11.1 | *** |
| Fat classification 3 | 4.8 a | 7.5 b | 7.5 b | 0.44 | *** |
| Temperature (oC) | |||||
| Hours post-mortem | |||||
| 1 | 36.6 a | 38.1 b | 38.5 b | 0.36 | *** |
| 3 | 26.3 a | 31.4 b | 34.1 c | 0.86 | *** |
| 5 | 19.5 a | 25.1 b | 27.4 c | 0.69 | *** |
| 7 | 16.0 a | 21.2 b | 23.2 c | 0.71 | *** |
| pH | |||||
| Hours post-mortem | |||||
| 1 | 6.61 | 6.50 | 6.62 | 0.053 | NS |
| 3 | 6.26 a | 6.04 b | 6.03 b | 0.072 | ** |
| 5 | 5.98 a | 5.70 b | 5.62 b | 0.076 | *** |
| 7 | 5.87 a | 5.54 b | 5.50 b | 0.058 | *** |
| Temperature (°C) @ pH 6.0 | 19.7 a | 29.7 b | 31.6 b | 1.28 | *** |
| Variable | G0 | G0AL | AL | SED | Significance 1 |
|---|---|---|---|---|---|
| 5/6th rib interface 2 | |||||
| pH | 5.61 | 5.59 | 5.57 | 0.025 | NS |
| L 3 | 31.3 a | 34.5 b | 34.3 b | 1.10 | * |
| a 3 | 20.3 a | 21.0 a | 22.5 b | 0.59 | ** |
| b 3 | 12.6 a | 13.4 b | 13.7 b | 0.36 | * |
| Saturation | 23.8 a | 24.9 a | 26.3 b | 0.64 | ** |
| Hue | 31.8 | 32.7 | 31.4 | 0.62 | NS |
| Variable | G0 | G0AL | AL | SED | Significance 2 |
|---|---|---|---|---|---|
| pH (72 h post-mortem) | 5.61 a | 5.53 b | 5.51 b | 0.028 | ** |
| L* | 45.3 | 45.5 | 45.8 | 0.79 | NS |
| a* | 12.5 a | 14.2 b | 15.2 b | 0.55 | *** |
| b* | 10.6 a | 11.8 b | 12.2 b | 0.49 | ** |
| Saturation | 16.4 a | 18.5 b | 19.5 b | 0.67 | *** |
| Hue | 40.4 | 39.6 | 38.9 | 0.95 | NS |
| % Metmyoglobin | 20.8 a | 20.6 a | 22.1 b | 0.60 | * |
| % Deoxymyoglobin | 26.5 a | 18.6 b | 14.9 b | 2.53 | *** |
| % Oxymyoglobin | 52.7 a | 60.8 b | 63.0 b | 2.33 | *** |
| L* 3 | 46.0 | 45.2 | 45.3 | 0.83 | NS |
| a* 3 | 12.6 a | 14.2 b | 15.1 b | 0.63 | ** |
| b* 3 | 11.0 | 11.6 | 12.0 | 0.55 | NS |
| Saturation 3 | 16.7 a | 18.4 b | 19.3 b | 0.76 | * |
| Hue 3 | 41.1 | 39.4 | 38.5 | 1.05 | NS |
| % Metmyoglobin 3 | 20.5 a | 20.7 a | 22.3 b | 0.68 | * |
| % Deoxymyoglobin 3 | 26.0 a | 18.8 b | 15.2 b | 2.93 | ** |
| % Oxymyoglobin 3 | 53.5 a | 60.5 b | 62.5 b | 2.67 | * |
| L* 4 | 45.5 | 45.4 | 45.6 | 1.37 | NS |
| a* 4 | 13.2 | 13.9 | 14.7 | 0.92 | NS |
| b* 4 | 11.2 | 11.5 | 11.9 | 0.84 | NS |
| Saturation 4 | 17.4 | 18.1 | 18.9 | 1.12 | NS |
| Hue 4 | 40.1 | 39.7 | 39.1 | 1.64 | NS |
| % Metmyoglobin 4 | 20.4 | 20.8 | 22.3 | 1.04 | 0.069 |
| % Deomyoglobin 4 | 24.6 | 19.3 | 16.1 | 4.36 | NS |
| % Oxymyoglobin 4 | 55.0 | 59.9 | 61.6 | 3.99 | NS |
| Variable | G0 | G0AL | AL | SED | Significance 2 |
|---|---|---|---|---|---|
| L* | 45.6 | 46.5 | 46.6 | 0.77 | NS |
| a* | 18.0 a | 19.7 b | 20.9 b | 0.62 | *** |
| b* | 14.7 | 15.0 | 15.1 | 0.45 | NS |
| Saturation | 23.2 a | 24.8 b | 25.8 b | 0.71 | ** |
| Hue | 39.3 a | 37.4 b | 35.9 c | 0.66 | *** |
| % Metmyoglobin | 23.6 a | 23.4 a | 24.9 b | 0.48 | ** |
| % Deoxymyoglobin | 6.1 | 5.8 | 5.1 | 0.57 | NS |
| % Oxymyoglobin | 70.3 | 70.8 | 70.0 | 0.85 | NS |
| L* 3 | 46.4 | 46.2 | 46.1 | 0.79 | NS |
| a* 3 | 18.0 a | 19.7 b | 20.8 b | 0.71 | ** |
| b* 3 | 15.0 | 14.9 | 14.9 | 0.50 | NS |
| Saturation 3 | 23.4 | 24.7 | 25.6 | 0.81 | 0.061 |
| Hue 3 | 39.7 a | 37.2 b | 35.6 c | 0.73 | *** |
| % Metmyoglobin 3 | 23.4 a | 23.4 a | 25.0 b | 0.55 | ** |
| % Deoxymyoglobin 3 | 5.6 | 6.0 | 5.5 | 0.60 | NS |
| % Oxymyoglobin 3 | 71.0 | 70.6 | 69.5 | 0.92 | NS |
| L* 4 | 46.0 | 46.3 | 46.3 | 1.33 | NS |
| a* 4 | 19.5 | 19.2 | 19.9 | 0.96 | NS |
| b* 4 | 15.6 | 14.7 | 14.5 | 0.73 | NS |
| Saturation 4 | 24.9 | 24.2 | 24.7 | 1.11 | NS |
| Hue 4 | 38.7 | 37.6 | 36.3 | 1.14 | NS |
| % Metmyoglobin 4 | 23.0 a | 23.6 a,b | 25.2 b | 0.82 | ** |
| % Deomyoglobin 4 | 5.6 | 6.0 | 5.5 | 0.97 | NS |
| % Oxymyoglobin 4 | 71.4 | 70.4 | 69.3 | 1.45 | NS |
| Variable | G0 | G0AL | AL | SED | Significance 1 |
|---|---|---|---|---|---|
| Composition (g/kg) | |||||
| Lipid | 7 a | 19 b | 28 c | 3.2 | *** |
| Moisture | 756 a | 741 b | 734 c | 3.5 | *** |
| Protein | 232 | 234 | 231 | 3.4 | NS |
| Myoglobin | 7.4 | 7.2 | 7.8 | 0.44 | NS |
| Glycolytic enzyme activity 2 | |||||
| LDH | 839 a | 958 b | 974 b | 26.0 | ** |
| PFK | 89 a | 148 b | 143 b | 9.0 | *** |
| Oxidative enzyme activity | |||||
| ICDH | 0.86 | 0.80 | 0.81 | 0.056 | NS |
| COX | 9.2 a | 16.7 b | 17.0 b | 1.40 | *** |
| CS | 4.8 | 4.6 | 4.6 | 0.48 | NS |
| Fibre type profile (%) | |||||
| MyHC I | 16.3 | 18.3 | 19.8 | 1.54 | 0.089 |
| MyHC IIA | 46.4 | 40.2 | 43.9 | 5.63 | NS |
| MyHC IIX | 33.4 | 36.6 | 35.9 | 6.47 | NS |
| 1-Hour Aerobic Exposure | 24-Hour Aerobic Exposure | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| L* | a* | b* | C* | H* | L* | a* | b* | C* | H* | ||
| Carcass weight | r | −0.05 | −0.03 | −0.16 | −0.10 | −0.18 | −0.03 | 0.13 | 0.03 | 0.10 | −0.11 |
| P | |||||||||||
| Fat score | r | −0.08 | 0.25 | −0.05 | 0.13 | −0.34 | −0.12 | 0.38 | 0.26 | 0.36 | −0.14 |
| P | 0.031 | 0.014 | 0.019 | ||||||||
| Conformation score | r | 0.18 | −0.09 | 0.06 | −0.03 | 0.16 | 0.11 | −0.03 | −0.01 | −0.03 | −0.01 |
| P | |||||||||||
| Intramuscular fat | r | −0.07 | 0.23 | 0.02 | 0.16 | −0.23 | −0.09 | 0.12 | 0.12 | 0.14 | 0.00 |
| P | |||||||||||
| pH | r | −0.43 | −0.17 | −0.32 | −0.26 | −0.22 | −0.35 | −0.09 | −0.28 | −0.17 | −0.20 |
| P | 0.005 | 0.039 | 0.024 | 0.074 | |||||||
| Myoglobin | r | −0.39 | 0.15 | −0.33 | −0.06 | −0.53 | −0.37 | 0.56 | 0.18 | 0.47 | −0.44 |
| P | 0.012 | 0.037 | 0.000 | 0.017 | 0.000 | 0.002 | 0.004 | ||||
| %Met | r | −0.55 | 0.2 | −0.42 | −0.08 | −0.75 | −0.59 | 0.12 | −0.53 | −0.13 | −0.73 |
| P | 0.000 | 0.007 | <0.0001 | <0.0001 | 0.0003 | <0.0001 | |||||
| %Deoxy | r | −0.09 | −0.73 | −0.48 | −0.69 | 0.26 | −0.61 | −0.05 | −0.49 | −0.23 | −0.49 |
| P | <0.0001 | 0.002 | <0.0001 | <0.0001 | 0.001 | 0.001 | |||||
| %Oxy | r | 0.25 | 0.71 | 0.62 | 0.75 | −0.06 | 0.74 | −0.03 | 0.63 | 0.23 | 0.74 |
| P | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |||||
| Temperature at pH | r | 0.21 | 0.39 | 0.30 | 0.39 | −0.08 | 0.10 | 0.46 | 0.44 | 0.50 | −0.07 |
| P | 0.013 | 0.053 | 0.011 | 0.002 | 0.004 | 0.001 | |||||
| ICDH | r | −0.22 | 0.08 | −0.10 | 0.00 | −0.18 | −0.33 | 0.23 | −0.15 | 0.09 | −0.43 |
| P | 0.034 | 0.005 | |||||||||
| LDH | r | 0.40 | −0.22 | 0.21 | −0.03 | 0.50 | 0.37 | −0.20 | 0.12 | −0.09 | 0.37 |
| P | 0.010 | 0.001 | 0.017 | 0.018 | |||||||
| PFK | r | 0.36 | −0.17 | 0.16 | −0.03 | 0.38 | 0.44 | 0.02 | 0.17 | 0.07 | 0.15 |
| P | 0.021 | 0.013 | 0.004 | ||||||||
| COX | r | −0.21 | −0.23 | −0.31 | −0.30 | −0.10 | −0.11 | −0.07 | −0.22 | −0.14 | −0.17 |
| P | 0.050 | 0.055 | |||||||||
| CS | r | −0.02 | −0.14 | −0.05 | −0.11 | 0.13 | 0.06 | 0.14 | 0.22 | 0.19 | 0.11 |
| P | |||||||||||
| IIX | r | 0.30 | −0.04 | 0.27 | 0.10 | 0.40 | 0.45 | 0.11 | 0.40 | 0.23 | 0.32 |
| P | 0.055 | 0.009 | 0.003 | 0.010 | 0.043 | ||||||
| IIA | r | −0.24 | −0.04 | −0.25 | −0.14 | −0.29 | −0.32 | −0.31 | −0.44 | −0.38 | −0.14 |
| P | 0.065 | 0.041 | 0.053 | 0.004 | 0.013 | ||||||
| I | r | −0.08 | −0.14 | −0.20 | −0.18 | −0.10 | −0.30 | 0.16 | −0.02 | 0.11 | −0.20 |
| P | 0.056 | ||||||||||
| Air Exposure | Classes | n | CDA Functions | Total Accuracy | Class Error Rates | |||
|---|---|---|---|---|---|---|---|---|
| G0 | G0AL | AL | C | |||||
| 1 h | G0, AL | 28 | 1 | 77% | 24% | - | 23% | - |
| 24 h | G0, AL | 28 | 1 | 90% | 14% | - | 6% | - |
| 1 h | G0, G0AL, AL | 43 | 1 | 48% | 42% | 70% | 42% | - |
| 24 h | G0, G0AL, AL | 43 | 1 | 63% | 26% | 54% | 31% | - |
| 24 h | G0, C | 43 | 1 | 83% | 27% | - | - | 12% |
| Air Exposure | Classes | n | Spectral Pre-Processing | PLS Factors | Total Accuracy | Class Error Rates | |||
|---|---|---|---|---|---|---|---|---|---|
| G0 | G0AL | AL | C | ||||||
| 1 h | G0, AL | 28 | Abs, SNV | 2 | 78% | 22% | - | 21% | - |
| 1 h | G0, AL | 28 | Abs, SG | 1 | 84% | 13% | - | 14% | - |
| 24 h | G0, AL | 28 | Abs, SNV | 3 | 86% | 19% | - | 9% | - |
| 24 h | G0, AL | 28 | Abs, SG | 2 | 89% | 18% | - | 5% | - |
| 1 h | G0, G0AL, AL | 43 | Abs, SNV | 2 | 54% | 34% | 66% | 37% | - |
| 1 h | G0, G0AL, AL | 43 | Abs, SG | 2 | 60% | 24% | 74% | 20% | - |
| 24 h | G0, G0AL, AL | 43 | Abs, SNV | 3 | 64% | 35% | 52% | 20% | - |
| 24 h | G0, G0AL, AL | 43 | Abs, SG | 5 | 60% | 27% | 70% | 21% | - |
| 24 h | G0, C | 43 | Abs, SG | 2 | 81% | 28% | - | - | 15% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moloney, A.P.; O’Riordan, E.G.; McGee, M.; Picard, B.; Monahan, F.J.; Moran, L.; Cama-Moncunill, R. Does Finishing at Pasture Influence the Colour of Muscle from Suckler Bulls and Can Colour Be Used to Authenticate Their Pre-Slaughter Diet? Foods 2022, 11, 2281. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11152281
Moloney AP, O’Riordan EG, McGee M, Picard B, Monahan FJ, Moran L, Cama-Moncunill R. Does Finishing at Pasture Influence the Colour of Muscle from Suckler Bulls and Can Colour Be Used to Authenticate Their Pre-Slaughter Diet? Foods. 2022; 11(15):2281. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11152281
Chicago/Turabian StyleMoloney, Aidan P., Edward G. O’Riordan, Mark McGee, Brigitte Picard, Frank J. Monahan, Lara Moran, and Raquel Cama-Moncunill. 2022. "Does Finishing at Pasture Influence the Colour of Muscle from Suckler Bulls and Can Colour Be Used to Authenticate Their Pre-Slaughter Diet?" Foods 11, no. 15: 2281. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11152281