
Improvement in Emulsifying Capacity of Goose Liver Protein Treated by pH Shifting with Addition of Sodium Tripolyphosphate and Its Proteomics Analysis

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Goose Liver
2.2. Extraction and Treatment
2.3. Measurement of Protein Solubility
2.4. Determination of Surface Hydrophobicity
2.5. Intrinsic Fluorescence Analysis
2.6. Sulfhydryl Content
2.7. Emulsifying Properties
2.8. Particle Size
2.9. Proteomic Analysis
2.9.1. Extraction of Emulsion-Layer Protein
2.9.2. Protein Digestion
2.9.3. Protein Identification by MS
2.10. Statistical Analysis
3. Results and Discussion
3.1. Protein Solubility
3.2. Surface Hydrophobicity
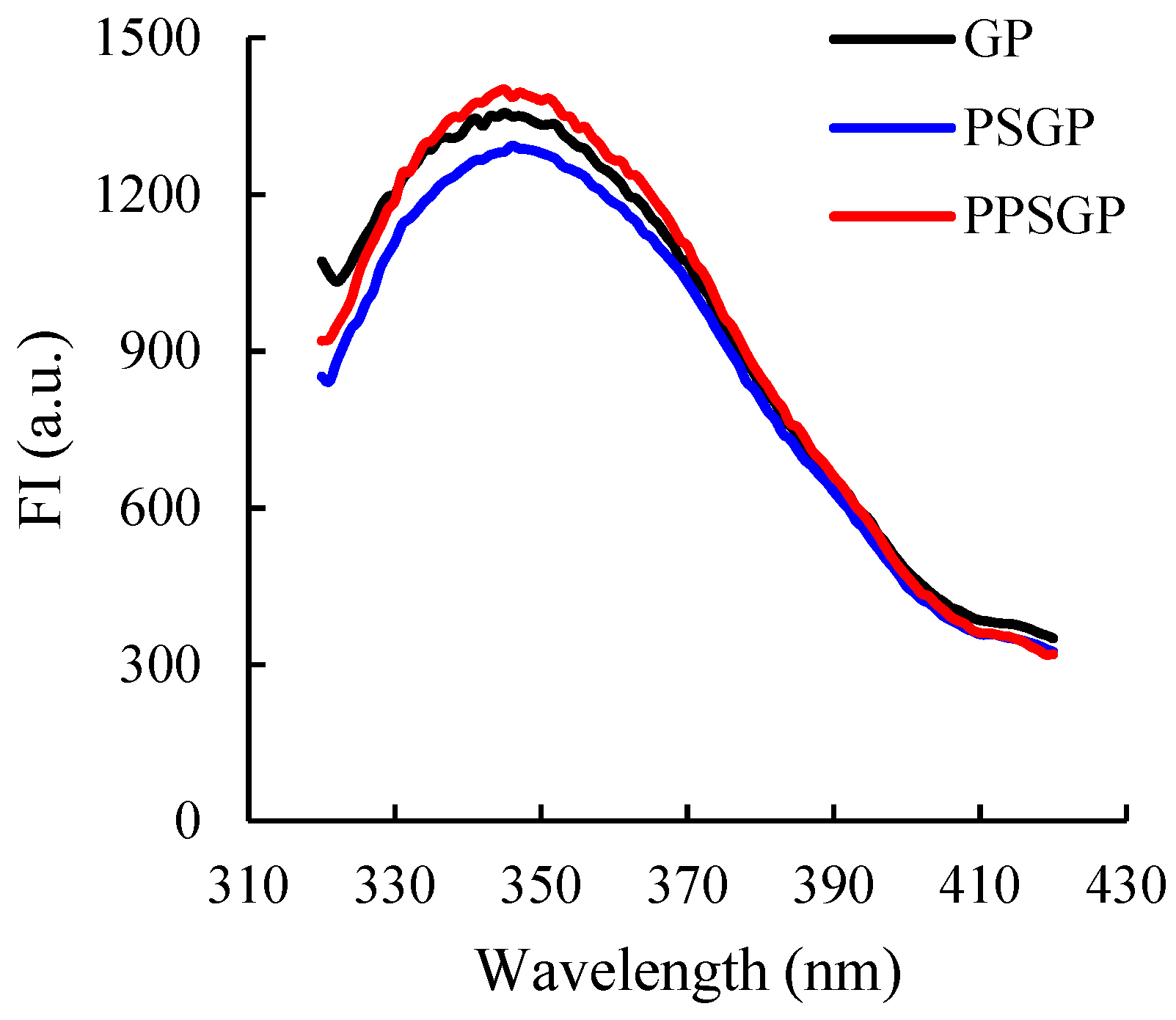
3.3. Intrinsic Fluorescence
3.4. Sulfhydryl Content
3.5. Emulsifying Property
3.6. Particle Size
3.7. Proteomic Analysis for Proteins Involved in Interfacial Layer
3.8. Correlation Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, L.; Zou, Y.; Zhang, K.; Yu, H. Ultrasonic-assisted alkaline extraction of duck liver protein and its antioxidant. Food Sci. 2017, 38, 222–228. [Google Scholar]
- Chen, R.; Zong, Z.Q. Analysis of nutritional composition in muscle and liver of Lueyang black-bone chicken. Hubei Agr. Sci. 2013, 52, 3402–3405. [Google Scholar]
- Xia, L.L.; Wang, Q.Q.; Yang, B.; Sun, X.X.; Zhang, Y.H.; Liu, L.; Geng, T.Y.; Gong, D.Q. Study on changes of blood chemistry indexes, hepatic routine nutritional composition and expression of lipid metabolism-associated genes during the recovery of geese with fatty livers. China Anim. Husb. Vet. Med. 2016, 43, 967–972. [Google Scholar]
- Huang, F.; Wu, W. Antidiabetic effect of a new peptide from Squalus mitsukurii liver (S-8300) in streptozocin-induced diabetic mice. J. Pharm. Pharmacol. 2005, 57, 1575–1580. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Li, T. Study on anti-coagulation activity in Tibetan Medicine Yak liver protein in vitro. Lishizhen Med. Mater. Med. Res. 2012, 23, 1927–1928. [Google Scholar]
- Kristinsson, H.G.; Hultin, H.O. Changes in conformation and subunit assembly of cod myosin at low and high pH and after subsequent refolding. J. Agric. Food Chem. 2003, 51, 7187–7196. [Google Scholar] [CrossRef] [PubMed]
- Ingadottir, B.; Kristinsson, H.G. Gelation of protein isolates extracted from tilapia light muscle by pH shift processing. Food Chem. 2010, 118, 789–798. [Google Scholar] [CrossRef]
- Goto, Y. Conformational states of β-lactamase: Molten-globule states at acidic and alkaline pH with high salt. Biochemistry 1989, 28, 945–952. [Google Scholar] [CrossRef]
- Wang, Q.; Jin, Y.; Xiong, Y.L. Heating-aided pH shifting modifies hemp seed protein structure, cross-linking, and emulsifying properties. J. Agric. Food Chem. 2018, 66, 10827–10834. [Google Scholar] [CrossRef] [PubMed]
- Goto, Y.; Calciano, L.J.; Fink, A.L. Acid-induced folding of proteins. Proc. Natl. Acad. Sci. USA 1990, 87, 573–577. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Ding, J.; Andrade, J.; Rababah, T.M.; Almajwal, A.; Abulmeaty, M.M.; Feng, H. Modifying the physicochemical properties of pea protein by pH-shifting and ultrasound combined treatments. Ultrason. Sonochem. 2017, 38, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wu, M.; Wang, Y.; Li, K.; Du, J.; Bai, Y. Effect of pH-shifting treatment on structural and heat induced gel properties of peanut protein isolate. Food Chem. 2020, 325, 126921. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, C.; Zhao, J.; Ma, T.; He, Z.; Huang, M.; Wang, Y. Modification of structure and functionalities of ginkgo seed proteins by pH-shifting treatment. Food Chem. 2021, 358, 129862. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.J.; Lin, Q.L.; Li, Z.H.; Xu, S.Y.; Kim, J.M. Effect of pH-shifting treatment on the Biochemical and thermal properties of myofibril protein. Adv. Mater. Res. 2011, 236, 2231–2235. [Google Scholar] [CrossRef]
- Li, X. Study on Extraction and Processing Properties of Goose Liver Proteins. Master’s Thesis, Nanjing Agriculture University, Nanjing, China, March 2018. [Google Scholar]
- Sung, H.Y.; Chen, H.J.; Liu, T.Y.; Su, J.C. Improvement of the functionalities of soy protein isolate through chemical phosphorylation. J. Food Sci. 1983, 48, 716–721. [Google Scholar] [CrossRef]
- Moure, A.; Sineiro, J.; Dominguez, H.; Parajo, J.C. Functionality of oilseed protein products: A review. Food Res. Int. 2006, 39, 945–963. [Google Scholar] [CrossRef]
- Miedzianka, J.; Peksa, A. Effect of pH on phosphorylation of potato protein isolate. Food Chem. 2013, 138, 2321–2326. [Google Scholar] [CrossRef]
- Yin, C.Y.; Yang, L.; Zhao, H.; Li, C.P. Improvement of antioxidant activity of egg white protein by phosphorylation and conjugation of epigallocatechin gallate. Food Res. Int. 2014, 64, 855–863. [Google Scholar] [CrossRef]
- Xiong, Z.Y.; Ma, M.H. Enhanced ovalbumin stability at oil-water interface by phosphorylation and identification of phosphorylation site using MALDI-TOF mass spectrometry. Colloids Surf. B Biointerfaces 2017, 153, 253–262. [Google Scholar] [CrossRef]
- Thaiphanit, S.; Anprung, P. Physicochemical and emulsion properties of edible protein concentrate from coconut (Cocos nucifera L.) processing by-products and the influence of heat treatment. Food Hydrocoll. 2016, 52, 756–765. [Google Scholar] [CrossRef]
- Kim, N.; Kwon, D.; Nam, Y. Effects of phosphorylation and acetylation on functional properties and structure of soy protein. Korean J. Food Sci. Technol. 1988, 20, 625–630. [Google Scholar]
- Hu, Z.; Qiu, L.; Sun, Y.; Xiong, H.; Ogra, Y. Improvement of the solubility and emulsifying properties of rice bran protein by phosphorylation with sodium trimetaphosphate. Food Hydrocoll. 2019, 96, 288–299. [Google Scholar] [CrossRef]
- Chen, G.; Wang, S.; Feng, B.; Jiang, B.; Miao, M. Interaction between soybean protein and tea polyphenols under high pressure. Food Chem. 2019, 277, 632–638. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Zhang, M.; Liu, H.; Li, Q.; Xue, D.; Nian, Y.; Zhao, D.; Shan, K.; Dai, C.; Li, C. Ultrasound treatment can increase digestibility of myofibrillar protein of pork with modified atmosphere packaging. Food Chem. 2022, 377, 131811. [Google Scholar] [CrossRef]
- Yan, S.; Xu, J.; Zhang, S.; Li, Y. Effects of flexibility and surface hydrophobicity on emulsifying properties: Ultrasound-treated soybean protein isolate. LWT 2021, 142, 110881. [Google Scholar] [CrossRef]
- Chen, X.; Zou, Y.; Han, M.; Pan, L.; Xing, T.; Xu, X.; Zhou, G. Solubilisation of myosin in a solution of low ionic strength L-histidine: Significance of the imidazole ring. Food Chem. 2016, 196, 42–49. [Google Scholar] [CrossRef]
- Maghamian, N.; Goli, M.; Najarian, A. Ultrasound-assisted preparation of double nano-emulsions loaded with glycyrrhizic acid in the internal aqueous phase and skim milk as the external aqueous phase. LWT 2021, 141, 110850. [Google Scholar] [CrossRef]
- Jorge, S.; Capelo, J.L.; LaFramboise, W.; Satturwar, S.; Korentzelos, D.; Bastacky, S.; Quiroga-Garza, G.; Dhir, R.; Wiśniewski, J.R.; Lodeiro, C.; et al. Absolute quantitative proteomics using the total protein approach to identify novel clinical immunohistochemical markers in renal neoplasms. BMC Med. 2021, 19, 196. [Google Scholar] [CrossRef]
- Hu, H.; Wu, J.; Li-Chan, E.C.Y.; Zhu, L.; Zhang, F.; Xu, X.; Fan, G.; Wang, L.; Huang, X.; Pan, S. Effects of ultrasound on structural and physical properties of soy protein isolate (SPI) dispersions. Food Hydrocoll. 2013, 30, 647–655. [Google Scholar] [CrossRef]
- Damodaran, S. Functional properties. In Food Proteins-Properties and Characterization; Nakai, S., Modler, H.W., Eds.; Wiley-VCH: Weinheim, Germany, 1996; pp. 167–233. [Google Scholar]
- Abdollahi, M.; Rezaei, M.; Jafarpour, A.; Undeland, I. Sequential extraction of gel-forming proteins, collagen and collagen hydrolysate from gutted silver carp (Hypophthalmichthys molitrix), a biorefinery approach. Food Chem. 2018, 242, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wang, W.; Ma, X.; Lv, R.; Watharkar, R.B.; Ding, T.; Ye, X.; Liu, D. Effect of pH-shifting treatment on structural and functional properties of whey protein isolate and its interaction with (−)-epigallocatechin-3-gallate. Food Chem. 2019, 274, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Xiong, Y.L.; Chen, J. pH shifting alters solubility characteristics and thermal stability of soy protein isolate and its globulin fractions in different pH, salt concentration, and temperature conditions. J. Agric. Food Chem. 2010, 58, 8035–8042. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, D.; Wang, J.; Yang, Y.; Zhang, L.; Li, J.; Wang, S. Functional properties and structural characteristics of phosphorylated pea protein isolate. Int. J. Food Sci. Technol. 2020, 55, 2002–2010. [Google Scholar] [CrossRef]
- Sheng, L.; Ye, S.; Han, K.; Zhu, G.; Ma, M.; Cai, Z. Consequences of phosphorylation on the structural and foaming properties of ovalbumin under wet-heating conditions. Food Hydrocoll. 2019, 91, 166–173. [Google Scholar] [CrossRef]
- Jiang, L.Z.; Wang, Z.J.; Li, Y.; Meng, X.H.; Sui, X.N.; Qi, B.K.; Zhou, L.Y. Relationship between surface hydrophobicity and structure of soy protein isolate subjected to different ionic strength. Int. J. Food Prop. 2015, 18, 1059–1074. [Google Scholar] [CrossRef]
- Jambrak, A.R.; Mason, T.J.; Lelas, V.; Herceg, Z.; Herceg, I.L. Effect of ultrasound treatment on solubility and foaming properties of whey protein suspensions. J. Food Eng. 2008, 86, 281–287. [Google Scholar] [CrossRef]
- Broersen, K.; Van Teeffelen, A.M.M.; Vries, A.; Voragen, A.G.J.; Hamer, R.J.; De Jongh, H.H.J. Do sulfhydryl groups affect aggregation and gelation properties of ovalbumin. J. Agric. Food Chem. 2006, 54, 5166–5174. [Google Scholar] [CrossRef]
- Xiong, Z.; Zhang, M.; Ma, M. Emulsifying properties of ovalbumin: Improvement and mechanism by phosphorylation in the presence of sodium tripolyphosphate. Food Hydrocoll. 2016, 60, 29–37. [Google Scholar] [CrossRef]
- Yan, C.; Zhou, Z. Solubility and emulsifying properties of phosphorylated walnut protein isolate extracted by sodium trimetaphosphate. LWT 2021, 143, 111117. [Google Scholar] [CrossRef]
- Stanciuc, N.; Aprodu, I.; Rapeanu, G.; Bahrim, G. Fluorescence spectroscopy and molecular modeling investigations on the thermally induced structural changes of bovine beta-lactoglobulin. Innov. Food Sci. Emerg. 2012, 15, 50–56. [Google Scholar] [CrossRef]
- Ghobadi, S.; Ashrafi-Kooshk, M.R.; Mahdiuni, H.; Khodarahmi, R. Enhancement of intrinsic fluorescence of human carbonic anhydrase II upon topiramate binding: Some evidence for drug-induced molecular contraction of the protein. Int. J. Biol. Macromol. 2018, 108, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Yang, Y.L.; Zhou, P.; Zhang, X.; Wang, J.Y. Effects of high pressure modification on conformation and gelation properties of myofibrillar protein. Food Chem. 2017, 217, 678–686. [Google Scholar] [CrossRef] [PubMed]
- Gong, K.J.; Shi, A.M.; Liu, H.Z.; Liu, L.; Hu, H.; Adhikari, B.; Wang, Q. Emulsifying properties and structure changes of spray and freeze-dried peanut protein isolate. J. Food Eng. 2016, 170, 33–40. [Google Scholar] [CrossRef]
- Xi, C.; Kang, N.; Zhao, C.; Liu, Y.; Sun, Z.; Zhang, T. Effects of pH and different sugars on the structures and emulsification properties of whey protein isolate-sugar conjugates. Food Biosci. 2020, 33, 100507. [Google Scholar] [CrossRef]
- Liu, J.; Wan, Y.; Ren, L.; Li, M.; Lv, Y.; Guo, S.; Waqar, K. Physical-chemical properties and in vitro digestibility of phosphorylated and glycosylated soy protein isolate. LWT 2021, 152, 112380. [Google Scholar] [CrossRef]
- Zhao, Q.; Hong, X.; Fan, L.; Liu, Y.; Li, J. Solubility and emulsifying properties of perilla protein isolate: Improvement by phosphorylation in the presence of sodium tripolyphosphate and sodium trimetaphosphate. Food Chem. 2022, 382, 132252. [Google Scholar] [CrossRef] [PubMed]
- Benelhadj, S.; Gharsallaoui, A.; Degraeve, P.; Attia, H.; Ghorbel, D. Effect of pH on the functional properties of Arthrospira (Spirulina) platensis protein isolate. Food Chem. 2016, 194, 1056–1063. [Google Scholar] [CrossRef] [PubMed]
- Lam, R.S.; Nickerson, M.T. The effect of pH and temperature pre-treatments on the physicochemical and emulsifying properties of whey protein isolate. LWT 2015, 60, 427–434. [Google Scholar] [CrossRef]
- Diao, X.; Guan, H.; Zhao, X.; Chen, Q.; Kong, B. Properties and oxidative stability of emulsions prepared with myofibrillar protein and lard diacylglycerols. Meat Sci. 2016, 115, 16–23. [Google Scholar] [CrossRef]
- Xue, S.; Yu, X.; Li, X.; Zhao, X.; Han, M.; Xu, X.; Zhou, G. Structural changes and emulsion properties of goose liver proteins obtained by isoelectric solubilisation/precipitation processes. LWT 2019, 102, 190–196. [Google Scholar] [CrossRef]
- Yu, Y.; Guan, Y.; Liu, J.; Hedi, W.; Yu, Y.; Zhang, T. Molecular structural modification of egg white protein by pH-shifting for improving emulsifying capacity and stability. Food Hydrocoll. 2021, 121, 107071. [Google Scholar] [CrossRef]
- O’sullivan, J.; Murray, B.; Flynn, C.; Norton, I. The effect of ultrasound treatment on the structural, physical and emulsifying properties of animal and vegetable proteins. Food Hydrocoll. 2016, 53, 141–154. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Beamer, S.K.; Yang, H.; Jaczynski, J. Micro-emulsification/encapsulation of krill oil by complex coacervation with krill protein isolated using isoelectric solubilization/precipitation. Food Chem. 2018, 244, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.H.; Zou, Y.F.; Xu, X.L.; Wu, J.Q.; Zhou, G.H. Evaluation of structural changes in raw and heated meat batters prepared with different lipids using Raman spectroscopy. Food Res. Int. 2011, 44, 2955–2961. [Google Scholar] [CrossRef]
- Li-Chan, E.; Nakai, S.; Wood, D.F. Hydrophobicity and solubility of meat proteins and their relationship to emulsifying properties. J. Food Sci. 1984, 49, 345–350. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Particle Size (nm) | PDI | |
|---|---|---|---|
| GPI | GP | 2614.33 ± 48.69 b | 0.22 ± 0.07 a |
| PSGP | 2838.67 ± 56.37 a | 0.24 ± 0.06 a | |
| PPSGP | 2575.33 ± 69.15 b | 0.02 ± 0.02 b | |
| GPI emulsion | GP | 2668.33 ± 229.84 c | 0.18 ± 0.02 b |
| PSGP | 3497.00 ± 132.35 a | 0.33 ± 0.15 a | |
| PPSGP | 3172.33 ± 232.00 b | 0.45 ± 0.09 a | |
| Protein ID | Description | MW (kDa) | pI | GRAVY | iBAQ Value |
|---|---|---|---|---|---|
| P01989 | Hemoglobin subunit alpha-A | 15.44 | 8.54 | 0.068 | 5.17 × 109 |
| R0KA48 | Histone H4 | 11.37 | 11.36 | −0.521 | 1.79 × 109 |
| U3J6E8 | Apovitellenin-1 | 9.49 | 9.34 | 0.24 | 1.75 × 109 |
| R0KK84 | ATP synthase subunit beta, mitochondrial | 13.80 | 5.32 | 0.117 | 1.68 × 109 |
| U3J1L1 | Glyceraldehyde-3-phosphate dehydrogenase | 35.84 | 8.87 | −0.065 | 1.50 × 109 |
| R0KFP7 | Glutathione S-transferase | 25.34 | 8.87 | −0.319 | 6.70 × 108 |
| U3IHG8 | Fructose-bisphosphate aldolase | 39.27 | 8.74 | −0.266 | 6.59 × 108 |
| R0L1Y3 | 60 kDa heat shock protein, mitochondrial | 61.03 | 5.61 | −0.069 | 5.83 × 108 |
| R0JKI4 | Glutathione-requiring prostaglandin D synthase | 22.46 | 6.83 | −0.215 | 4.56 × 108 |
| A0A0K1R5T3 | ADP/ATP translocase | 32.79 | 9.73 | 0.092 | 4.04 × 108 |
| Protein ID | Description | MW (kDa) | pI | GRAVY | iBAQ Value |
|---|---|---|---|---|---|
| P01989 | Hemoglobin subunit alpha-A | 15.44 | 8.54 | 0.068 | 2.52 × 109 |
| U3J6E8 | Apovitellenin-1 | 9.49 | 9.34 | 0.24 | 1.67 × 109 |
| U3J1L1 | Glyceraldehyde-3-phosphate dehydrogenase | 35.84 | 8.87 | −0.065 | 1.39 × 109 |
| R0KA48 | Histone H4 | 11.37 | 11.36 | −0.521 | 1.35 × 109 |
| R0KK84 | ATP synthase subunit beta, mitochondrial | 13.80 | 5.32 | 0.117 | 1.03 × 109 |
| U3IHG8 | Fructose-bisphosphate aldolase | 39.27 | 8.74 | −0.266 | 7.45 × 108 |
| R0KPC7 | L-lactate dehydrogenase | 36.75 | 7.73 | −0.034 | 5.19 × 108 |
| R0L1Y3 | 60 kDa heat shock protein, mitochondrial | 61.03 | 5.61 | −0.069 | 4.98 × 108 |
| R0M210 | Histone H2B | 13.96 | 10.31 | −0.738 | 4.65 × 108 |
| R0M714 | Betaine--homocysteine S-methyltransferase | 44.06 | 6.42 | −0.348 | 4.35 × 108 |
| Protein ID | Description | MW (kDa) | pI | GRAVY | iBAQ Value |
|---|---|---|---|---|---|
| P01989 | Hemoglobin subunit alpha-A | 15.44 | 8.54 | 0.068 | 4.12 × 109 |
| R0KA48 | Histone H4 | 11.37 | 11.36 | −0.521 | 1.83 × 109 |
| U3J1L1 | Glyceraldehyde-3-phosphate dehydrogenase | 35.84 | 8.87 | −0.065 | 1.81 × 109 |
| U3J6E8 | Apovitellenin-1 | 9.49 | 9.34 | 0.24 | 1.22 × 109 |
| R0KK84 | ATP synthase subunit beta, mitochondrial | 13.80 | 5.32 | 0.117 | 1.10 × 109 |
| U3IHG8 | Fructose-bisphosphate aldolase | 39.27 | 8.74 | −0.266 | 1.03 × 109 |
| R0KPC7 | L-lactate dehydrogenase | 36.75 | 7.73 | −0.034 | 5.63 × 108 |
| R0KFP7 | Glutathione S-transferase | 25.34 | 8.87 | −0.319 | 5.37 × 108 |
| R0L1Y3 | 60 kDa heat shock protein, mitochondrial | 61.03 | 5.61 | −0.069 | 4.95 × 108 |
| A0A0K1R5T3 | ADP/ATP translocase | 32.79 | 9.73 | 0.092 | 4.64 × 108 |
| Protein ID | Phosphorylation Site | Amino Acid | pI | GRAVY | iBAQ Value | ||
|---|---|---|---|---|---|---|---|
| GP | PSGP | PPSGP | |||||
| U3J1L1 | 174 | threonine | 8.87 | −0.065 | 1.50 × 109 | 1.39 × 109 | 1.81 × 109 |
| R0KFP7 | 37 | threonine | 8.87 | −0.319 | 6.70 × 108 | 3.82 × 108 | 5.37 × 108 |
| A8RRQ6 | 237 | serine | 7.62 | −0.326 | 1.83 × 108 | 2.02 × 108 | 2.91 × 108 |
| R0JSM9 | 263 | serine | 5.00 | −0.710 | 9.51 × 107 | 8.55 × 107 | 9.34 × 107 |
| R0LAD1 | 4 | threonine | 9.78 | −0.266 | 4.13 × 107 | 3.31 × 107 | 4.05 × 107 |
| R0LCM7 | 18 | tyrosine | 7.20 | −0.399 | 1.08 × 107 | 8.40 × 105 | 5.74 × 106 |
| U3IQ39 | 20 | serine | 9.24 | −0.975 | 8.70 × 106 | 5.28 × 106 | 4.27 × 106 |
| A0A068C605 | 7 | threonine | 9.39 | 0.027 | 7.02 × 106 | 3.98 × 106 | 5.51 × 106 |
| R0LFA3 | 61 | threonine | 5.61 | −0.084 | 3.52 × 106 | 3.83 × 106 | 3.26 × 106 |
| U3IW62 | 110 | lysine | 8.91 | −0.540 | 1.39 × 105 | / | / |
| U3IFD3 | 668 | threonine | 5.44 | −0.877 | / | 5.22 × 104 | 4.59 × 105 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Bai, Y.; Yu, X.; Li, Z.; Wang, P.; Xu, X. Improvement in Emulsifying Capacity of Goose Liver Protein Treated by pH Shifting with Addition of Sodium Tripolyphosphate and Its Proteomics Analysis. Foods 2022, 11, 3329. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11213329
Zhang Y, Bai Y, Yu X, Li Z, Wang P, Xu X. Improvement in Emulsifying Capacity of Goose Liver Protein Treated by pH Shifting with Addition of Sodium Tripolyphosphate and Its Proteomics Analysis. Foods. 2022; 11(21):3329. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11213329
Chicago/Turabian StyleZhang, Yulong, Yun Bai, Xiaobo Yu, Zhen Li, Peng Wang, and Xinglian Xu. 2022. "Improvement in Emulsifying Capacity of Goose Liver Protein Treated by pH Shifting with Addition of Sodium Tripolyphosphate and Its Proteomics Analysis" Foods 11, no. 21: 3329. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11213329