
Prevalence and Antimicrobial Resistance of Bacterial Foodborne Pathogens Isolated from Raw Bivalve Molluscs Subjected to Consumption in Poland during a Ten-Year Period

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Isolation and Identification of Bacterial Pathogens
2.2.1. Salmonella spp.
2.2.2. L. monocytogenes
2.2.3. V. parahaemolyticus
2.2.4. S. aureus
2.3. Antimicrobial Resistance Testing
2.3.1. Salmonella spp.
2.3.2. L. monocytogenes
2.3.3. V. parahaemolyticus
2.3.4. S. aureus
2.4. Statistical Analysis
3. Results
3.1. Prevalence and Antimicrobial Resistance of Bacterial Pathogens
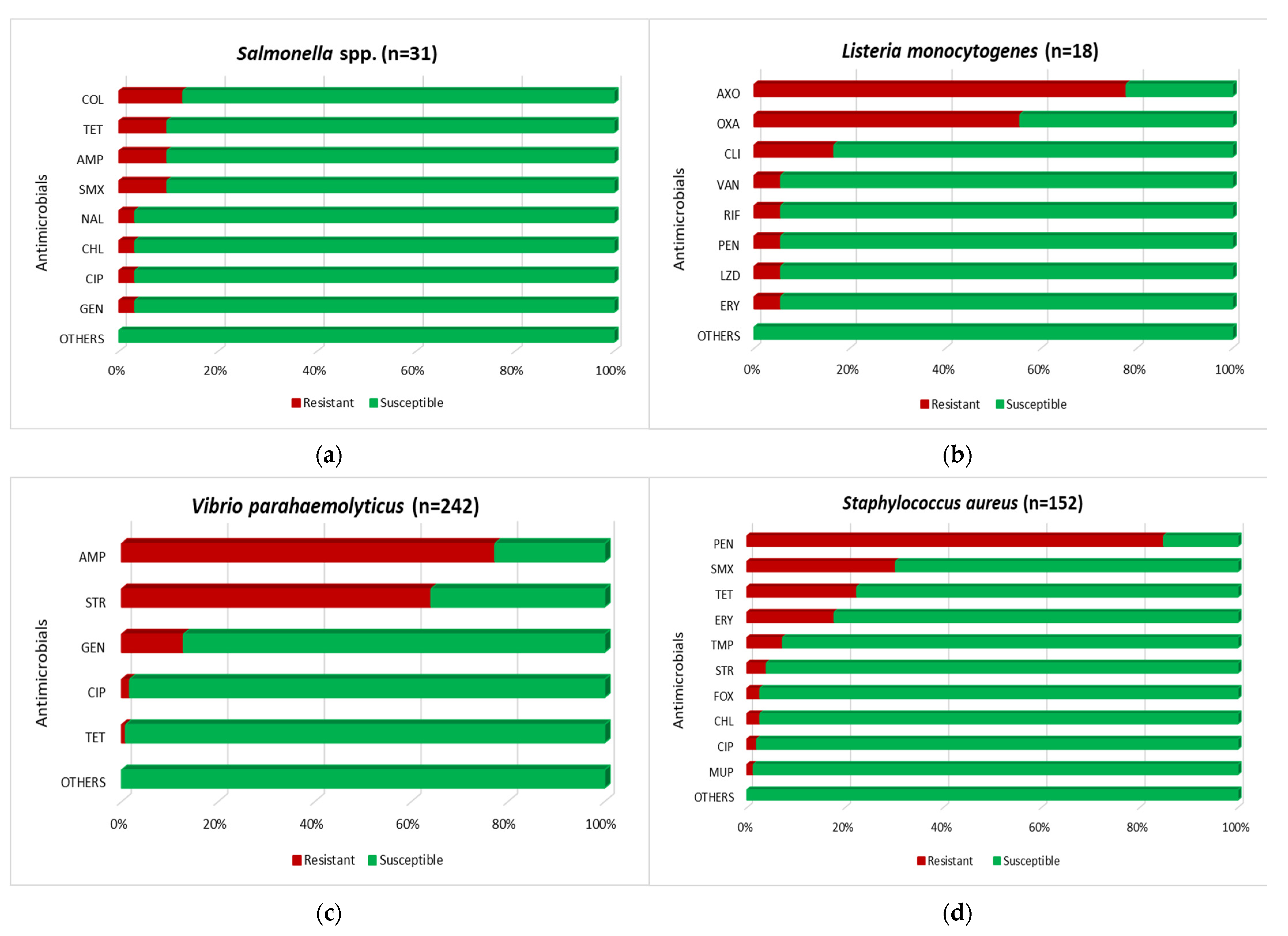
3.1.1. Salmonella spp.
3.1.2. L. monocytogenes
3.1.3. V. parahaemolyticus
3.1.4. S. aureus
3.2. Simultaneous Occurrence of Bacterial Pathogens
3.3. Seasonal Prevalence of Bacterial Pathogens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mahaffey, K.R.; Clickner, R.P.; Jeffries, R.A. Methylmercury and omega-3 fatty acids: Co-occurrence of dietary sources with emphasis on fish and shellfish. Environ. Res. 2008, 107, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.; Cunha, A.; Castilho, F.; Romalde, J.L.; Pereira, M.J. Microbial contamination and purification of bivalve shellfish: Crucial aspects in monitoring and future perspectives—A mini-review. Food Control 2011, 22, 805–816. [Google Scholar] [CrossRef]
- Ali, A.; Parisi, A.; Conversano, M.C.; Iannacci, A.; D’Emilio, F.; Mercurio, V.; Normanno, G. Food-borne bacteria associated with seafoods: A brief review. J. Food Qual. Hazards Control 2020, 7, 4–10. [Google Scholar] [CrossRef]
- Feldhusen, F. The role of seafood in bacterial foodborne diseases. Microbes Infect. 2000, 2, 1651–1660. [Google Scholar] [CrossRef]
- Hernroth, B.E.; Conden-Hanson, A.C.; Rehnstam-Holm, A.S.; Girones, R.; Allard, A.K. Environmental factors influencing human pathogens and their potential indicator organisms in the blue mussel Mytilus edulis: The first Scandinavian report. App. Environ. Microbiol. 2002, 68, 4523–4533. [Google Scholar] [CrossRef] [Green Version]
- Stewart, J.R.; Gast, R.J.; Fujioka, R.S.; Solo-Gabriele, H.M.; Meschke, J.S.; Amaral-Zettler, L.A.; del Castillo, E.; Polz, M.F.; Collier, T.K.; Strom, M.S.; et al. The coastal environment and human health: Microbial indicators, pathogens, sentinels and reservoirs. Environ. Health 2008, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.C.; Danyluk, M.D.; Otwell, W.S. Pathogens in raw foods: What the salad bar learn from the raw bar. Curr. Opin. Biotechnol. 2009, 20, 172–177. [Google Scholar] [CrossRef]
- Marques, A.; Nunes, M.L.; Moore, S.K.; Strom, M.S. Climate change and seafood safety: Human health implications. Food Res. Int. 2010, 43, 1766–1779. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Trinanes, J.A.; Taylor, N.G.H.; Hartnell, R.; Siitonem, A.; Martinez-Urtaza, J. Emerging Vibrio risk at high latitudes in response to ocean warming. Nat. Clim. Chang. 2013, 3, 73–77. [Google Scholar] [CrossRef]
- Tirado, M.C.; Clarke, R.; Jaykus, L.A.; McQuatters-Gollop, A.; Frank, J.M. Climate change and food safety: A review. Food Res. Int. 2010, 43, 1745–1765. [Google Scholar] [CrossRef]
- Huss, H.H.; Reilly, A.; Embarek, P.K.B. Prevention and control of hazards in seafood. Food Control 2000, 11, 149–156. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control (EFSA and ECDC). The European Union One Health 2020 zoonoses report. EFSA J. 2021, 19, 6971. [Google Scholar] [CrossRef]
- Cabello, F.C. Heavy use of prophylactic antibiotics in aquaculture: A growing problem for human and animal health and for the environment. Environ. Microbiol. 2006, 8, 1137–1144. [Google Scholar] [CrossRef]
- Wright, G.D. Antibiotic resistance in the environment: A link to the clinic? Curr. Opin. Microbiol. 2010, 13, 589–594. [Google Scholar] [CrossRef]
- Hariharan, H.; Amadi, V. Shellfish as reservoirs of bacterial pathogens. J. Coast. Life Med. 2016, 4, 253–258. [Google Scholar] [CrossRef]
- ISO 6887-3:2003; Microbiology of Food and Animal Feeding Stuffs—Preparation of Test Samples, Initial Suspension and Decimal Dilutions for Microbiological Examination—Part 3: Specific Rules for the Preparation of Fish and Fishery Products. International Organization for Standardization: Geneva, Switzerland, 2003.
- ISO 6579:2002; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Detection of Salmonella spp. International Organization for Standardization: Geneva, Switzerland, 2002.
- Grimont, P.A.D.; Weill, F.-X. Antigenic Formulae of Salmonella Serovars, 9th ed.; WHO Collaborating Centre for Research on Salmonella, Institute Pasteur: Paris, France, 2007. [Google Scholar]
- ISO 11290-1:1996; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Detection and Enumeration of Listeria Monocytogenes—Part 1: Detection Method. International Organization for Standardization: Geneva, Switzerland, 1996.
- Doumith, M.; Buchrieser, C.; Glaser, P.; Jacquet, C.; Martin, P. Differentiation of the major Listeria monocytogenes serovars by multiplex PCR. J. Clin. Microbiol. 2004, 42, 3819–3822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ISO/TS 21872-1:2007; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Detection of Potentially Enteropathogenic Vibrio spp.—Part 1: Detection of Vibrio Parahaemolyticus and Vibrio Cholera. International Organization for Standardization: Geneva, Switzerland, 2007.
- Kim, Y.B.; Okuda, J.; Matsumoto, C.; Takahashi, N.; Hashimoto, S.; Nishibuchi, M. Identification of Vibrio parahaemolyticus strains at the species level by PCR targeted to the toxR gene. J. Clin. Microbiol. 1999, 37, 1173–1177. [Google Scholar] [CrossRef] [Green Version]
- ISO 6888-3:2003; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Coagulase-Positive Staphylococci (Staphylococcus aureus and Other Species)—3: Detection and MPN Technique for Low Numbers. International Organization for Standardization: Geneva, Switzerland, 2003.
- European Commission (EC). Commission Implementing Decision 2013/652/EU of 12 November 2013 on the Monitoring and Reporting of Antimicrobial Resistance in Zoonotic and Commensal Bacteria. Off. J. Eur. Union 2013, 303, 26–39. [Google Scholar]
- CLSI Supplement M100-S22; Performance Standards for Antimicrobial Susceptibility Testing—Twenty Second Informational Supplement. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012.
- CLSI Guideline M45-A2; Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria, 3rd ed. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2016.
- Lyon, S.A.; Berrang, M.E.; Fedorka-Cray, P.J.; Fletcher, D.L.; Meinersmann, R.J. Antimicrobial resistance of Listeria monocytogenes isolated from a poultry further processing plant. Foodborne Pathog. Dis. 2008, 5, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Escolar, C.; Gómez, D.; del Carmen Rota García, M.; Conchello, P.; Herrera, A. Antimicrobial resistance profiles of Listeria monocytogenes and Listeria innocua isolated from ready-to-eat products of animal origin in Spain. Foodborne Pathog. Dis. 2017, 14, 357–363. [Google Scholar] [CrossRef]
- CLSI Guideline M45-A2; Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria—Approved Guideline, 2nd ed. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2010.
- Baker-Austin, C.; McArthur, J.V.; Tuckfield, R.C.; Najarro, M.; Lindell, A.H.; Gooch, J.; Stepanauskas, R. Antibiotic resistance in the shellfish pathogen Vibrio parahaemolyticus isolated from the coastal water and sediment of Georgia and South Carolina, USA. J. Food Prot. 2008, 71, 2552–2558. [Google Scholar] [CrossRef] [PubMed]
- Shaw, K.S.; Rosenberg Goldstein, R.E.; He, X.; Jacobs, J.M.; Crump, B.C.; Sapkota, A.R. Antimicrobial susceptibility of Vibrio vulnificus and Vibrio parahaemolyticus recovered from recreational and commercial areas of Chesapeake Bay and Maryland Coastal Bays. PLoS ONE 2014, 9, e89616. [Google Scholar] [CrossRef] [PubMed]
- International Commission on Microbiological Specifications for Foods (ICMSF). Microorganisms in Foods 8: Use of Data for Assessing Process Control and Product Acceptance, 2nd ed.; Springer: New York, NY, USA, 2011; pp. 107–133. [Google Scholar]
- European Commission. Commission Regulation (EC) No. 2073/2005 of 15 November 2005 on Microbiological Criteria for Foodstuffs. Off. J. Eur. Union. 2005, 338, 1–26. [Google Scholar]
- Amagliani, G.; Brandi, G.; Schiavano, G.F. Incidence and role of Salmonella in seafood safety. Food Res. Int. 2012, 45, 780–788. [Google Scholar] [CrossRef]
- Phan, T.T.; Khai, L.T.L.; Ogasawara, N.; Tam, N.T.; Okatani, A.T.; Akiba, M.; Hayashidani, H. Contamination of Salmonella in retail meats and shrimps in the Mekong Delta, Vietnam. J. Food Prot. 2005, 68, 1077–1080. [Google Scholar] [CrossRef]
- Kumar, R.; Surendran, P.K.; Thampuran, N. Distribution and genotypic characterization of Salmonella serovars isolated from tropical seafood in cochin, India. J. Appl. Microbiol. 2009, 106, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Thompson, C.K.; Wang, Q.; Bag, S.K.; Franklin, N.; Shadbolt, C.T.; Howard, P.; Fearnley, E.J.; Quinn, H.E.; Sintchenko, V.; Hope, K.G. Epidemiology and whole genome sequencing of an ongoing point-source Salmonella Agona outbreak associated with sushi consumption in western Sydney, Australia 2015. Epidemiol. Infect. 2017, 145, 2062–2071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novoslavskij, A.; Terentjeva, M.; Eizenberga, I.; Valciņa, O.; Bartkevičs, V.; Bērziņš, A. Major foodborne pathogens in fish and fish products: A review. Ann. Microbiol. 2016, 66, 1–15. [Google Scholar] [CrossRef]
- Hansen, C.H.; Vogel, B.F.; Gram, L. Prevalence and survival of Listeria monocytogenes in Danish aquatic and fish processing environments. J. Food Prot. 2006, 69, 2113–2122. [Google Scholar] [CrossRef]
- Papadopoulou, C.; Economou, E.; Zakas, G.; Salamoura, C.; Dontorou, C.; Apostolou, J. Microbiological and pathogenic contaminants of seafood in Greece. J. Food Qual. 2007, 30, 28–42. [Google Scholar] [CrossRef]
- Gambarin, P.; Magnabosco, C.; Losio, M.N.; Pavoni, E.; Gattuso, A.; Arcangeli, G.; Favretti, M. Listeria monocytogenes in ready-to-eat seafood and potential hazards for the consumers. Int. J. Microbiol. 2012, 2012, 497635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jami, M.; Ghanbari, M.; Zunabovic, M.; Domig, K.J.; Kneifel, W. Listeria monocytogenes in aquatic food products—A review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 798–813. [Google Scholar] [CrossRef]
- Barrett, K.A.; Nakao, J.H.; Taylor, E.V.; Eggers, C.; Gould, L.H. Fish-associated foodborne disease outbreaks: United States, 1998–2015. Foodborne Pathog. Dis. 2017, 14, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Johansson, T.; Rantala, L.; Palmu, L.; Honkanen-Buzalski, T. Occurrence and typing of Listeria monocytogenes strains in retail vacuum-packed fish products and in a production plant. Int. J. Food Microbiol. 1999, 47, 111–119. [Google Scholar] [CrossRef]
- Corcoran, D.; Clancy, D.; O’Mahony, M.; Grant, K.; Hyland, E.; Shanaghy, N.; Whyte, P.; McLauchlin, J.; Moloney, A.; Fanning, S. Comparison of Listeria monocytogenes strain types in Irish smoked salmon and other foods. Int. J. Hyg. Environ. Health 2006, 209, 527–534. [Google Scholar] [CrossRef]
- Momtaz, H.; Yadollahi, S. Molecular characterization of Listeria monocytogenes isolated from fresh seafood samples in Iran. Diagn. Pathol. 2013, 8, 149. [Google Scholar] [CrossRef] [Green Version]
- Davies, A.R.; Capell, C.; Jehanno, D.; Nychas, G.J.E.; Kirby, R.M. Incidence of foodborne pathogens on European fish. Food Control 2001, 12, 67–71. [Google Scholar] [CrossRef]
- Roque, A.; Lopez-Joven, C.; Lacuesta, B.; Elandaloussi, L.; Wagley, S.; Furones, M.D.; Ruiz-Zarzuela, I.; de Blas, I.; Rangdale, R.; Gomez-Gil, B. Detection and identification of tdh- and trh-positive Vibrio parahaemolyticus strains from four species of cultured bivalve molluscs on the Spanish Mediterranean coast. Appl. Environ. Microbiol. 2009, 75, 7574–7577. [Google Scholar] [CrossRef] [Green Version]
- Biasizzo, M.; Kirbiś, A.; Marinśek, J. Bacterial contamination of shellfish in Slovenia. Slov. Vet. Res. 2005, 42, 83–87. [Google Scholar]
- Nakaguchi, Y. Contamination by Vibrio parahaemolyticus and its virulent strains in seafood marketed in Thailand, Vietnam, Malaysia, and Indonesia. Trop. Med. Health 2013, 41, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Roldán, E.M.; Rodriguez, E.E.; Vicente, C.N.; Fernández-Crehuet Navajas, M.; Abril Moreno, O. Microbial contamination of bivalve molluscs used for human consumption. J. Food Saf. 2011, 31, 257–261. [Google Scholar] [CrossRef]
- Atwill, E.R.; Jeamsripong, S. Bacterial diversity and potential risk factors associated with Salmonella contamination of seafood products sold in retail markets in Bangkok, Thailand. Peer J. 2021, 9, e12694. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, D.; Leoni, F.; Rocchegiani, E.; Santarelli, S.; Canonico, C.; Masini, L.; DiTrani, V. First clinical report of pandemic Vibrio parahaemolyticus O3:K6 infection in Italy. J. Clin. Microbiol. 2008, 46, 2144–2145. [Google Scholar] [CrossRef] [PubMed]
- Ndraha, N.; Hsiao, H.-I. Influence of climatic factors on the temporal occurrence and distribution of total and pathogenic Vibrio parahaemolyticus in oyster culture environments in Taiwan. Food Microbiol. 2021, 98, 103765. [Google Scholar] [CrossRef] [PubMed]
- Hatha, A.A.M.; Lakshmanaperumalsamy, P. Prevalence of Salmonella in fish and crustaceans from markets in Coimbatore, South India. Food Microbiol. 1997, 14, 111–116. [Google Scholar] [CrossRef] [Green Version]
- Eng, S.-K.; Pusparajah, P.; Mutalib, N.-S.A.; Ser, H.-L.; Chan, K.-G.; Lee, L.-H. Salmonella: A review on pathogenesis, epidemiology and antibiotic resistance. Front. Life Sci. 2015, 8, 284–293. [Google Scholar] [CrossRef] [Green Version]
- Portes, A.B.; Rodrigues, G.; Leitão, M.P.; Ferrari, R.; Conte Junior, C.A.; Panzenhagen, P. Global distribution of plasmid-mediated colistin resistance mcr gene in Salmonella: A systematic review. J. Appl. Microbiol. 2022, 132, 872. [Google Scholar] [CrossRef]
- Peruzy, M.F.; Capuano, F.; Proroga, Y.T.R.; Cristiano, D.; Carullo, M.R.; Murru, N. Antimicrobial Susceptibility Testing for Salmonella Serovars Isolated from Food Samples: Five-Year Monitoring (2015–2019). Antibiotics 2020, 9, 365. [Google Scholar] [CrossRef]
- Noll, M.; Kleta, S.; Al Dahouk, S. Antibiotic susceptibility of 259 Listeria monocytogenes strains isolated from food, food-processing plants and human samples in Germany. J. Infect. Public Health. 2018, 11, 572–577. [Google Scholar] [CrossRef]
- Conter, M.; Paludi, D.; Zanardi, E.; Ghidini, S.; Vergara, A.; Ianieri, A. Characterization of antimicrobial resistance of foodborne Listeria monocytogenes. Int. J. Food Microbiol. 2009, 128, 497–500. [Google Scholar] [CrossRef]
- Sosnowski, M.; Lachtara, B.; Wieczorek, K.; Osek, J. Antimicrobial resistance and genotypic characteristics of Listeria monocytogenes isolated from food in Poland. Int. J. Food Microbiol. 2019, 289, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Elmahdi, S.; DaSilva, L.V.; Parveen, S. Antibiotic resistance of Vibrio parahaemolyticus and Vibrio vulnificus in various countries: A review. Food Microbiol. 2016, 57, 128–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottaviani, D.; Bacchiocchi, I.; Masini, L.; Leoni, F.; Carraturo, A.; Giammarioli, M.; Sbaraglia, G. Antimicrobial susceptibility of potentially pathogenic halophilic vibrios isolated from seafood. Int. J. Antimicrob. Agents 2001, 18, 135–140. [Google Scholar] [CrossRef]
- Wong, M.H.Y.; Liu, M.; Wan, H.Y.; Chen, S. Characterization of extended-spectrum-β-lactamase-producing Vibrio parahaemolyticus. Antimicrob. Agents Chemother. 2012, 56, 4026–4028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vestergaard, M.; Frees, D.; Ingmer, H. Antibiotic resistance and the MRSA problem. Microbiol. Spectr. 2019, 7, GPP3-0057-2018. [Google Scholar] [CrossRef] [PubMed]
- Marijani, E. Prevalence and Antimicrobial Resistance of Bacteria Isolated from Marine and Freshwater Fish in Tanzania. Int. J. Microbiol. 2022, 2022, 4652326. [Google Scholar] [CrossRef] [PubMed]
- Wellington, E.M.H.; Boxall, A.B.A.; Cross, P.; Feil, E.J.; Gaze, W.H.; Hawkey, P.M.; Johnson-Rollings, A.S.; Jones, D.L.; Lee, N.M.; Otten, W.; et al. The role of the natural environment in the emergence of antibiotic resistance in Gram-negative bacteria. Lancet Infect. Dis. 2013, 13, 155–165. [Google Scholar] [CrossRef]


{kind=link}
| Sample Type (Species of Bivalve Molluscs) | Country of Origin | Number of Samples | Total Number (%) of Samples | ||
|---|---|---|---|---|---|
| Clams | Manila clams | The Netherlands | 97 | 220 (22.0) | 437 (43.7) |
| Italy | 92 | ||||
| France | 31 | ||||
| Razor clams | The Netherlands | 78 | 78 (7.8) | ||
| Amandes | The Netherlands | 41 | 49 (4.9) | ||
| France | 8 | ||||
| Cockle | The Netherlands | 40 | 47 (4.7) | ||
| France | 7 | ||||
| Hard clams | Canada | 16 | 43 (4.3) | ||
| The Netherlands | 12 | ||||
| France | 12 | ||||
| Italy | 3 | ||||
| Mussels | The Netherlands | 114 | 269 (26.9) | ||
| Norway | 81 | ||||
| Denmark | 47 | ||||
| Spain | 15 | ||||
| France | 6 | ||||
| Italy | 4 | ||||
| Ireland | 2 | ||||
| Oysters | The Netherlands | 166 | 225 (22.5) | ||
| France | 52 | ||||
| Ireland | 7 | ||||
| Scallops | Norway | 41 | 69 (6.9) | ||
| The Netherlands | 18 | ||||
| France | 4 | ||||
| USA | 4 | ||||
| Italy | 2 | ||||
| All samples | 1000 | ||||
| Antimicrobials | Abbreviation | Application for Resistance Testing | |||
|---|---|---|---|---|---|
| Salmonella spp. | L. monocytogenes | V. parahaemolyticus | S. aureus | ||
| Ampicillin | AMP | + | + | + | − |
| Penicillin | PEN | − | + | − | + |
| Oxacillin | OXA | − | + | − | − |
| Ceftriaxone | AXO | − | + | − | − |
| Cefotaxime | FOT | + | − | − | − |
| Cefoxitin | FOX | − | − | − | + |
| Ceftazidime | TAZ | + | − | − | − |
| Meropenem | MERO | + | − | − | − |
| Chloramphenicol | CHL | + | − | + | + |
| Ciprofloxacin | CIP | + | + | + | + |
| Nalidixic acid | NAL | + | − | − | − |
| Gentamicin | GEN | + | + | + | + |
| Streptomycin | STR | − | + | + | + |
| Erythromycin | ERY | − | + | − | + |
| Gatifloxacin | GAT | − | + | − | − |
| Levofloxacin | LEVO | − | + | − | − |
| Mupirocin | MUP | − | − | − | + |
| Linezolid | LZD | − | + | − | − |
| Clindamycin | CLI | − | + | − | + |
| Colistin | COL | + | − | − | − |
| Vancomycin | VAN | − | + | − | + |
| Rifampicin | RIF | − | + | − | + |
| Tetracycline | TET | + | + | + | + |
| Sulfamethoxazole | SMX | + | − | − | + |
| Trimethoprim | TMP | + | − | − | + |
| Quinupristin/Dalfopristin | SYN | − | + | − | − |
| Trimethoprim/Sulfamethoxazole | SXT | − | + | − | − |
| Source | No. (%) of Samples Tested | No. (%) of Positive Samples for | ||||
|---|---|---|---|---|---|---|
| Sample Type | Country of Origin | Salmonella spp. | L. monocytogenes | V. parahaemolyticus | S. aureus | |
| Clams | The Netherlands | 268 | 7 | 4 | 92 | 58 |
| Italy | 95 | 6 | 2 | 42 | 19 | |
| France | 58 | 0 | 6 | 11 | 11 | |
| Canada | 16 | 0 | 0 | 6 | 0 | |
| Total (%) | 437 (43.7) | 13 (3.0) | 12 (2.7) | 151 (34.6) | 88 (20.1) | |
| Mussels | The Netherlands | 114 | 2 | 2 | 29 | 13 |
| Norway | 81 | 5 | 0 | 9 | 13 | |
| Denmark | 47 | 1 | 1 | 16 | 11 | |
| Spain | 15 | 0 | 0 | 1 | 0 | |
| France | 6 | 3 | 0 | 0 | 1 | |
| Italy | 4 | 0 | 0 | 1 | 0 | |
| Ireland | 2 | 0 | 0 | 0 | 1 | |
| Total (%) | 269 (26.9) | 11 (4.1) | 3 (1.1) | 56 (20.8) | 39 (14.5) | |
| Oysters | The Netherlands | 166 | 5 | 0 | 26 | 18 |
| France | 52 | 1 | 0 | 2 | 4 | |
| Ireland | 7 | 0 | 3 | 1 | 0 | |
| Total (%) | 225 (22.5) | 6 (2.7) | 3 (1.3) | 29 (12.9) | 22 (9.8) | |
| Scallops | Norway | 41 | 0 | 0 | 3 | 1 |
| The Netherlands | 18 | 1 | 0 | 3 | 1 | |
| France | 4 | 0 | 0 | 0 | 0 | |
| USA | 4 | 0 | 0 | 0 | 0 | |
| Italy | 2 | 0 | 0 | 0 | 1 | |
| Total (%) | 69 (6.9) | 1 (1.4) | 0 | 6 (8.7) | 3 (4.3) | |
| All samples (%) | 1000 (100) | 31 (3.1) | 18 (1.8) | 242 (24.2) | 152 (15.2) | |
| Bacterial Pathogens | No. of Positive Samples | ||
|---|---|---|---|
| Sample Type (No. of Samples) | Country Origin (No. of Samples) | Total (%) | |
| V. parahaemolyticus + S. aureus | clams (33), mussels (7), oysters (3) | the Netherlands (28), Italy (8), Denmark (4), France (3) | 43 (71.6) |
| Salmonella spp. + V. parahaemolyticus | clams (4), mussels (1) | the Netherlands (3), Denmark (1), Italy (1) | 5 (8.3) |
| L. monocytogenes + V. parahaemolyticus | clams (2), mussels (1), oysters (1) | the Netherlands (2), France (1), Ireland (1) | 4 (6.7) |
| Salmonella spp. + S. aureus | mussels (3), clams (2), oysters (2) | the Netherlands (2), Norway (2), Italy (2), France (1) | 7 (11.7) |
| Salmonella spp. + V. parahaemolyticus + S. aureus | clams (1) | the Netherlands (1) | 1 (1.7) |
| Sampling | No. of Samples | No. (%) of Positive Samples for Pathogens | ||||
|---|---|---|---|---|---|---|
| Period | Tested | Positive (%) | Salmonella spp. | L. monocytogenes | V. parahaemolyticus | S. aureus |
| warmer months (May–September) | 473 | 255 (53.9%) | 10 (2.1) | 12 (2.5) | 152 (32.1) | 81 (17.1) |
| colder months (October–April) | 527 | 188 (35.7%) | 21 (4.0) | 6 (1.1) | 90 (17.1) | 71 (13.5) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopatek, M.; Wieczorek, K.; Osek, J. Prevalence and Antimicrobial Resistance of Bacterial Foodborne Pathogens Isolated from Raw Bivalve Molluscs Subjected to Consumption in Poland during a Ten-Year Period. Foods 2022, 11, 3521. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11213521
Lopatek M, Wieczorek K, Osek J. Prevalence and Antimicrobial Resistance of Bacterial Foodborne Pathogens Isolated from Raw Bivalve Molluscs Subjected to Consumption in Poland during a Ten-Year Period. Foods. 2022; 11(21):3521. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11213521
Chicago/Turabian StyleLopatek, Magdalena, Kinga Wieczorek, and Jacek Osek. 2022. "Prevalence and Antimicrobial Resistance of Bacterial Foodborne Pathogens Isolated from Raw Bivalve Molluscs Subjected to Consumption in Poland during a Ten-Year Period" Foods 11, no. 21: 3521. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11213521