
Phenolic Profile, Antioxidant Activity and Amino Acid Composition of Moringa Leaves Fermented with Edible Fungal Strains

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Microbial Strains
2.3. Autoclaving and Fermentation
2.4. Analyses
2.4.1. Antioxidant Methods
2.4.2. Determination of Phenolic Compounds
2.4.3. Amino Acid Profile
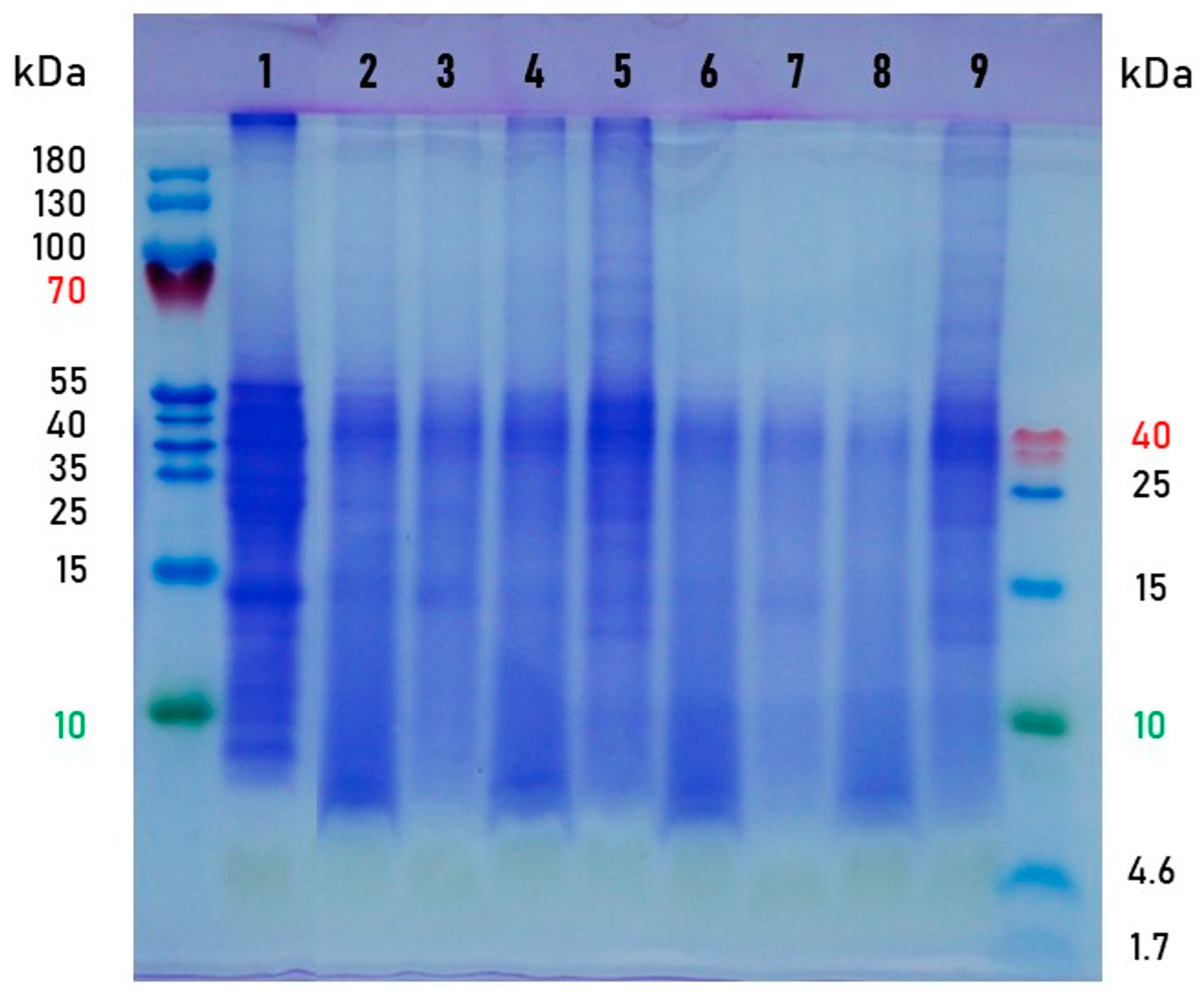
2.4.4. SDS-PAGE Electrophoresis
2.4.5. Glucosamine Determination
2.4.6. Dry Matter Determination
2.5. Statistical Analysis
3. Results and Discussion
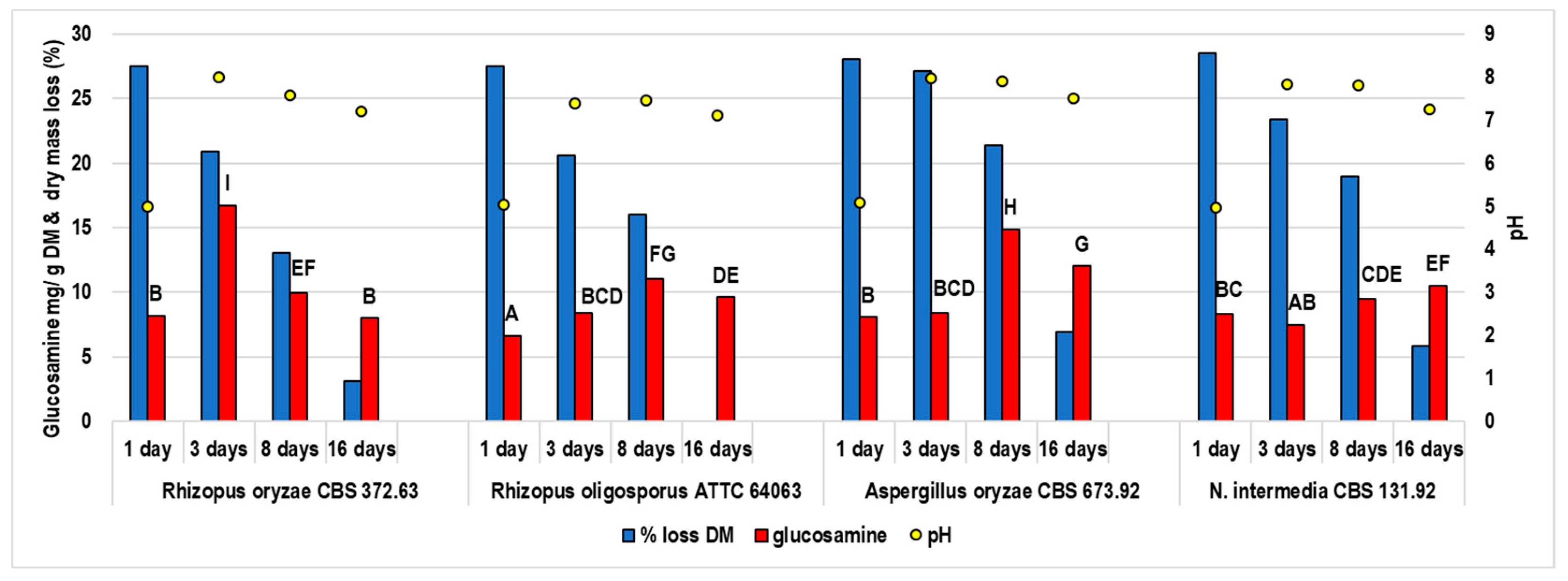
3.1. The Characteristics of the Fungal Growth during the Fermentation of Moringa Leaves
3.2. Antioxidant Activity in the Dried, Autoclaved, and Fermented Moringa Leaves
3.3. Determination of Phenolic Compounds in Dried, Autoclaved, and Fermented Samples by Using HPLC-ESI-TOF-MS
3.4. Amino Acid Profile and Protein in the Dried, Autoclaved, and Fermented Moringa Leaves
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kashyap, P.; Kumar, S.; Riar, C.S.; Jindal, N.; Baniwal, P.; Guiné, R.P.F.; Correia, P.M.R.; Mehra, R.; Kumar, H. Recent advances in drumstick (Moringa oleifera) leaves bioactive compounds: Composition, health benefits, bioaccessibility, and dietary applications. Antioxidants 2022, 11, 402. [Google Scholar] [CrossRef] [PubMed]
- Bourekoua, H.; Różyło, R.; Gawlik-Dziki, U.; Benatallah, L.; Zidoune, M.N.; Dziki, D. Evaluation of physical, sensorial, and antioxidant properties of gluten-free bread enriched with Moringa oleifera leaf powder. Eur. Food Res. Technol. 2018, 244, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Salem, A.S.; Salama, W.M.; Ragab, W.A. Prolonged shelf life of sour cream by adding Moringa oleifera leaves extract (MOLE) or Moringa oleifera oil (Moo). Am. J. Food Technol. 2015, 10, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Devisetti, R.; Sreerama, Y.N.; Bhattacharya, S. Processing effects on bioactive components and functional properties of moringa leaves: Development of a snack and quality evaluation. J. Food Sci. Technol. 2016, 53, 649–657. [Google Scholar] [CrossRef] [Green Version]
- Owens, J.D.; Astuti, M.; Kuswanto, K.R. Indigenous Fermented Foods of Southeast Asia; Owens, J.D., Ed.; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar] [CrossRef]
- Djonu, A.; Andayani, S.; Nursyam, H. Identification of Moringa oleifera leaves content fermented by Rhizopus oligosporus. Int. J. Sci. Technol. Res. 2018, 7, 102–104. [Google Scholar]
- Feitosa, P.R.B.; Santos, T.R.J.; Gualberto, N.C.; Narain, N.; de Aquino Santana, L.C.L. Solid-state fermentation with Aspergillus niger for the bio-enrichment of bioactive compounds in Moringa oleifera (moringa) leaves. Biocatal. Agric. Biotechnol. 2020, 27, 101709. [Google Scholar] [CrossRef]
- Shi, H.; Yang, E.; Li, Y.; Chen, X.; Zhang, J. Effect of solid-state fermentation on nutritional quality of leaf flour of the drumstick tree (Moringa oleifera Lam.). Front. Bioeng. Biotechnol. 2021, 9, 626628. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Yang, E.; Yang, H.; Huang, X.; Zheng, M.; Chen, X.; Zhang, J. Dynamic changes in the chemical composition and metabolite profiles of drumstick (Moringa oleifera Lam.) leaf flour during fermentation. LWT 2022, 155, 112973. [Google Scholar] [CrossRef]
- Dai, J.; Tao, L.; Shi, C.; Yang, S.; Li, D.; Sheng, J.; Tian, Y. Fermentation improves calcium bioavailability in Moringa oleifera leaves and prevents bone loss in calcium-deficient rats. Food Sci. Nutr. 2020, 8, 3692–3703. [Google Scholar] [CrossRef]
- Ali, M.W.; Ilays, M.Z.; Saeed, M.T.; Shin, D.-H. Comparative assessment regarding antioxidative and nutritional potential of Moringa oleifera leaves by bacterial fermentation. J. Food Sci. Technol. 2020, 57, 1110–1118. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, C.; Quirantes-Piné, R.; Fernández-Gutiérrez, A.; Segura-Carretero, A. Optimization of extraction method to obtain a phenolic compounds-rich extract from Moringa oleifera Lam leaves. Ind. Crops Prod. 2015, 66, 246–254. [Google Scholar] [CrossRef]
- Starzyńska-Janiszewska, A.; Stodolak, B.; Duliński, R.; Mickowska, B.; Sabat, R. Fermentation of colored quinoa seeds with Neurospora intermedia to obtain oncom-type products of favorable nutritional and bioactive characteristics. Cereal Chem. 2017, 94, 619–624. [Google Scholar] [CrossRef]
- Stodolak, B.; Starzyńska-Janiszewska, A.; Mika, M.; Wikiera, A. Rhizopus oligosporus and Lactobacillus plantarum co-fermentation as a tool for increasing the antioxidant potential of grass pea and flaxseed oil-cake tempe. Molecules 2020, 25, 4759. [Google Scholar] [CrossRef] [PubMed]
- Stodolak, B.; Starzyńska-Janiszewska, A.; Bączkowicz, M. Aspergillus oryzae (koji mold) and Neurospora intermedia (oncom mold) application for flaxseed oil cake processing. LWT 2020, 131, 109651. [Google Scholar] [CrossRef]
- Jennessen, J.; Schnürer, J.; Olsson, J.; Samson, R.A.; Dijksterhuis, J. Morphological characteristics of sporangiospores of the tempe fungus Rhizopus oligosporus differentiate it from other taxa of the R. microsporus group. Mycol. Res. 2008, 112, 547–563. [Google Scholar] [CrossRef]
- Perkins, D.D.; Davis, R.H. Evidence for safety of Neurospora species for academic and commercial uses. Appl. Environ. Microbiol. 2000, 66, 5107–5109. [Google Scholar] [CrossRef] [Green Version]
- Daba, G.M.; Mostafa, F.A.; Elkhateeb, W.A. The ancient koji mold (Aspergillus oryzae) as a modern biotechnological tool. Bioresour. Bioprocess. 2021, 8, 52. [Google Scholar] [CrossRef]
- Swain, T.; Hillis, W.E. The phenolic constituents of Prunus domestica. I. The quantitative analysis of phenolic constituents. J. Sci. Food Agric. 1959, 10, 63–68. [Google Scholar] [CrossRef]
- Cano, A.; Acosta, M.; Arnao, M.B. Hydrophilic and lipophilic antioxidant activity changes during on-vine ripening of tomatoes (Lycopersicon esculentum Mill.). Postharvest Biol. Technol. 2003, 28, 59–65. [Google Scholar] [CrossRef]
- Marambe, P.W.M.L.H.K.; Shand, P.J.; Wanasundara, J.P.D. An in-vitro investigation of selected biological activities of hydrolysed flaxseed (Linum usitatissimum L.) proteins. J. Am. Oil Chem. Soc. 2008, 85, 1155–1164. [Google Scholar] [CrossRef]
- Moore, S.; Stein, W. Chromatography of amino acids on sulfonated polystyrene resins. J. Biol. Chem. 1951, 92, 663–681. [Google Scholar] [CrossRef]
- Davidson, I. Hydrolysis of samples for amino acid analysis. In Methods in Molecular Biology, Vol. 211: Protein Sequencing Protocols, 2nd ed.; Smith, B.J., Ed.; Humana Press Inc.: Totowa, NJ, USA, 2003. [Google Scholar]
- Smith, A.J. Post column amino acid analysis. In Methods in Molecular Biology, Vol. 211: Protein Sequencing Protocols, 2nd ed.; Smith, B.J., Ed.; Humana Press Inc.: Totowa, NJ, USA, 2003. [Google Scholar]
- Schägger, H.; von Jagow, G. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal. Biochem. 1987, 166, 368–379. [Google Scholar] [CrossRef]
- Tsuji, A.; Kinoshita, T.; Hoshino, M. Analytical chemical studies on amino sugars. II. Determination of hexosamines using 3-methyl-2-benzothiazolone hydrazone hydrochloride. Chem. Pharm. Bull. 1969, 17, 1505–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vongsak, B.; Sithisarn, P.; Mangmool, S.; Thongpraditchote, S.; Wongkrajang, Y.; Gritsanapan, W. Maximizing total phenolics, total flavonoids contents and antioxidant activity of Moringa oleifera leaf extract by the appropriate extraction method. Ind. Crops Prod. 2013, 44, 566–571. [Google Scholar] [CrossRef]
- Huynh, N.T.; Van Camp, J.; Smagghe, G.; Raes, K. Improved release and metabolism in flavonoids by steered fermentation process: A review. Int. J. Mol. Sci. 2014, 15, 19369–19388. [Google Scholar] [CrossRef]
- Schmidt, C.G.; Gonçalves, L.M.; Prietto, L.; Hackbart, H.S.; Furlong, E.B. Antioxidant activity and enzyme inhibition of phenolic acids from fermented rice bran with fungus Rhizopus oryzae. Food Chem. 2014, 146, 371–377. [Google Scholar] [CrossRef] [Green Version]
- El-Katony, T.M.; El-Dein, M.M.N.; El-Fallal, A.A.; Ibrahim, N.G.; Mousa, M.M. Substrate-fungus interaction on the enzymatic and non-enzymatic antioxidant activities of solid state fermentation system. Bioresour. Bioprocess. 2020, 7, 28. [Google Scholar] [CrossRef]
- Balli, D.; Bellumori, M.; Pucci, L.; Gabriele, M.; Longo, V.; Paoli, P.; Melani, F.; Mulinacci, N.; Innocenti, M. Does fermentation really increase the phenolic contents in cereals? A study on millet. Foods. 2020, 9, 303. [Google Scholar] [CrossRef] [Green Version]
- Tang, P.-J.; Zhang, Z.-H.; Niu, L.-L.; Gu, C.-B.; Zheng, W.-Y.; Cui, H.-C.; Yuan, X.-H. Fusarium solani G6, a novel vitexin-producing endophytic fungus: Characterization, yield improvement and osteoblastic proliferation activity. Biotechnol. Lett. 2021, 43, 1371–1383. [Google Scholar] [CrossRef]
- Gu, C.B.; Ma, H.; Ning, W.J.; Niu, W.J.; Han, H.Y.; Yuan, X.H.; Fu, Y.J. Characterisation, culture medium optimization and antioxidant activity of an endophytic vitexin-producing fungus Dichotomopilus funicola Y3 from pigeon pea (Cajanus cajan (L.) Millsp.). J. Appl. Microbiol. 2018, 125, 1054–1065. [Google Scholar] [CrossRef]
- Gebru, Y.A.; Sbhatu, D.B. Effects of fungi-mediated solid-state fermentation on phenolic contents and antioxidant activity of brown and white teff (Eragrostis tef (Zucc.) Trotter) grains. J. Food Qual. 2020, 2020, 8819555. [Google Scholar] [CrossRef]
- Ajila, C.M.; Gassara, F.; Brar, S.K.; Verma, M.; Tyagi, R.D.; Valéro, J.R. Polyphenolic antioxidant mobilization in apple pomace by different methods of solid-state fermentation and evaluation of its antioxidant activity. Food Bioprocess Technol. 2012, 5, 2697–2707. [Google Scholar] [CrossRef]
- FAO. Dietary Protein Quality Evaluation in Human Nutrition. Report of an FAQ Expert Consultation; FAO Food and Nutrition Paper. Volume 92; FAO: Auckland, New Zealand, 2013. [Google Scholar] [PubMed]
- Shojaosadati, S.A.; Faraidouni, R.; Madadi-Nouei, A.; Mohamadpour, I. Protein enrichment of lignocellulosic substrates by solid state fermentation using Neurospora sitophila. Resour. Conserv. Recycl. 1999, 27, 73–87. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| FCRS (mg/g DM) | SA-ABTS˙+ (µmol Trolox/g DM) | SA-˙OH (IC50) | |||
|---|---|---|---|---|---|
| Dried moringa | 38.91 ± 0.82 ij | 505.44 ± 9.53 l | 0.80 ± 0.07 a | ||
| Autoclaved moringa | 36.44 ± 0.59 h | 442.32 ± 6.82 j | 1.60 ± 0.06 i | ||
| Fermented moringa | Strain | Time (days) | |||
| Rhizopus oryzae CBS 372.63 | 1 | 42.91 ± 1.01 l | 462.06 ± 6.93 k | 0.97 ± 0.05 bcd | |
| 3 | 29.70 ± 0.64 g | 316.57 ± 5.23 g | 0.84 ± 0.02 ab | ||
| 8 | 26.69 ± 0.73 f | 306.72 ± 15.70 fg | 0.87 ± 0.06 abc | ||
| 16 | 25.71 ± 0.45 e | 297.71 ± 9.30 f | 0.93 ± 0.12 abdc | ||
| Rhizopus oligosporus ATCC 64063 | 1 | 39.23 ± 0.60 jk | 533.67 ± 14.99 m | 1.21 ± 0.04 fgh | |
| 3 | 30.44 ± 0.63 g | 343.45 ± 2.93 h | 0.98 ± 0.01 bcd | ||
| 8 | 20.33 ± 0.54 b | 243.52 ± 4.07 b | 1.02 ± 0.05 cde | ||
| 16 | 18.46 ± 0.26 a | 216.70 ± 3.27 a | 1.16 ± 0.24 efg | ||
| Aspergillus oryzae CBS 673.92 | 1 | 38.12 ± 1.13 i | 552.29 ± 11.30 n | 1.16 ± 0.10 efg | |
| 3 | 21.68 ± 0.44 c | 257.17 ± 5.95 cd | 1.30 ± 0.11 gh | ||
| 8 | 22.92 ± 0.42 d | 271.72 ± 6.60 e | 1.20 ± 0.05 fgh | ||
| 16 | 21.85 ± 0.54 b | 267.17 ± 7.63 de | 1.16 ± 0.07 efg | ||
| Neurospora intermedia CBS 131.92 | 1 | 40.15 ± 0.46 k | 396.28 ± 8.83 i | 1.34 ± 0.07 h | |
| 3 | 23.24 ± 0.45 d | 269.56 ± 7.01 e | 0.97 ± 0.05 bcd | ||
| 8 | 22.39 ± 0.29 cd | 251.64 ± 9.69 bc | 1.04 ± 0.13 de | ||
| 16 | 18.95 ± 0.93 a | 226.36 ± 6.03 a | 1.08 ± 0.16 def | ||
| Dried Moringa | Autoclaved Moringa | Rhizopus oryzae CBS 372.63 | Rhizopus oligosporus ATCC 64063 | Aspergillus oryzae CBS 673.92 | Neurospora intermedia CBS 131.92 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Pa | Compound | - | 1 Day | 3 Days | 1 Day | 3 Days | 1 Day | 3 Days | 1 Day | 3 Days | |
| Hydroxycinnamic acids and derivatives | |||||||||||
| 1 | Glucopyranosyl-caffeoylquinic acid isomer a | 0.028 ± 0.001 a | 0.118 ± 0.008 c | 0.107 ± 0.000 b | <LOD | 0.106 b ± 0.000 b | <LOD | 0.107 ± 0.003 b | <LOD | 0.107 ± 0.005 b | <LOD |
| 4 | Glucopyranosyl-caffeoylquinic acid isomer b | <LOD | 0.069 ± 0.010 b | 0.049 ± 0.000 a | <LOD | 0.048 ± 0.001 a | <LOD | 0.051 ± 0.001 a | <LOD | 0.051 ± 0.001 a | <LOD |
| 6 | Glucopyranosyl-caffeoylquinic acid isomer c | 0.009 ± 0.001 a | 0.091 ± 0.003 d | 0.082 ± 0.000 c | <LOD | 0.078 ± 0.002 bc | <LOD | 0.075 ± 0.002 b | <LOD | 0.077 ± 0.006 bc | <LOD |
| 9 | 1-Caffeoylquinic acid | <LOD | 0.226 ± 0.011 e | 0.085 ± 0.001 c | 0.082 ± 0.000 c | 0.070 ± 0.003 b | 0.169 ± 0.001 d | 0.072 ± 0.001 b | 0.018 ± 0.001 a | 0.085 ± 0.002 c | <LOQ |
| 2 | 3-Caffeoylquinic acid | 1.180 ± 0.021 e | 0.355 ± 0.033 d | 0.204 ± 0.006 c | 0.064 ± 0.002 a | 0.155 ± 0.000 b | 0.158 ± 0.003 b | 0.149 ± 0.003 b | <LOD | 0.193 ± 0.009 c | <LOD |
| 7 | 4-Caffeoylquinic acid | 0.080 ± 0.007 b | 0.472 ± 0.034 g | 0.250 ± 0.001 f | 0.081 ± 0.003 b | 0.175 ± 0.005 cd | 0.206 ± 0.001 e | 0.164 ± 0.004 c | 0.017 ± 0.001 a | 0.202 ± 0.007 de | <LOQ |
| 3 | 3-p-Coumaroylquinic acid isomer a | 0.074 ± 0.006 b | 0.119 ± 0.004 d | 0.140 ± 0.005 f | 0.076 ± 0.001 b | 0.129 ± 0.003 e | 0.090 ± 0.000 c | 0.114 ± 0.004 d | <LOD | 0.116 ± 0.001 d | 0.058 ± 0.002 a |
| 5 | 3-p-Coumaroylquinic acid isomer b | 0.059 ± 0.003 b | 0.166 ± 0.000 f | 0.186 ± 0.003 g | 0.103 ± 0.001 c | 0.168 ± 0.002 f | 0.123 ± 0.003 d | 0.161 ± 0.007 ef | <LOD | 0.152 ± 0.007 e | 0.035 ± 0.001 a |
| 10 | 4-p-Coumaroylquinic acid isomer a | 0.003 ± 0.000 a | 0.167 ± 0.011 f | 0.132 ± 0.003 e | 0.091 ± 0.003 c | 0.111 ± 0.001 d | 0.102 ± 0.002 cd | 0.101 ± 0.006 cd | <LOD | 0.108 ± 0.004 d | 0.058 ± 0.003 b |
| 12 | 4-p-Coumaroylquinic acid isomer b | 0.004 ± 0.001 a | 0.252 ± 0.012 g | 0.216 ± 0.000 f | 0.133 ± 0.005 c | 0.193 ± 0.008 e | 0.151 ± 0.008 d | 0.190 ± 0.008 e | <LOD | 0.183 ± 0.002 e | 0.040 ± 0.002 b |
| 13 | 5-p-Coumaroylquinic acid isomer a | <LOD | 0.071 ± 0.007 b | 0.050 ± 0.001 a | 0.113 ± 0.004 d | 0.048 ± 0.000 a | 0.104 ± 0.000 c | 0.052 ± 0.001 a | <LOD | 0.051 ± 0.002 a | 0.068 ± 0.004 b |
| 14 | 5-p-Coumaroylquinic acid isomer b | <LOD | 0.074 ± 0.001 b | 0.051 ± 0.000 a | 0.118 ± 0.000 d | 0.049 ± 0.001 a | 0.107 ± 0.004 c | 0.052 ± 0.002 a | <LOD | 0.052 ± 0.001 a | <LOQ |
| 8 | 3-Feruloylquinic acid | 0.016 ± 0.001 a | 0.069 ± 0.001 f | 0.069 ± 0.001 f | 0.042 ± 0.000 b | 0.065 ± 0.002 e | 0.052 ± 0.000 c | 0.061 ± 0.003 d | <LOD | 0.061 ± 0.001 d | <LOD |
| 15 | 4-Feruloylquinic acid | <LOD | 0.068 ± 0.007 d | 0.058 ± 0.002 c | 0.042 ± 0.001 a | 0.056 ± 0.001 c | 0.048 ± 0.000 ab | 0.057 ± 0.002 a | <LOD | 0.054 ± 0.001 bc | <LOD |
| 17 | 5-Feruloylquinic acid | <LOD | 0.047 ± 0.004 b | 0.036 ± 0.000 a | 0.049 ± 0.001 b | 0.035 ± 0.001 a | 0.051 ± 0.001 b | 0.037 ± 0.001 a | <LOD | 0.037 ± 0.001 a | <LOD |
| Flavones and derivatives | |||||||||||
| 11 | Vicenin-2 (apigenin 6,8-C-dihexoside) isomer a | <LOQ | 0.408 ± 0.032 b | 0.748 ± 0.013 f | 0.474 ± 0.023 c | 0.707 ± 0.016 ef | 0.603 ± 0.025 d | 0.674 ± 0.030 e | 0.047 ± 0.001 a | 0.600 ± 0.026 d | 0.715 ± 0.001 ef |
| 16 | Vicenin-2 (apigenin 6,8-C-dihexoside) isomer c | <LOQ | 1.085 ± 0.033 d | 1.217 ± 0.006 f | 0.893 ± 0.014 b | 1.146 ± 0.041 e | 1.011 ± 0.028 b | 1.046 ± 0.029 cd | 0.179 ± 0.002 a | 1.000 ± 0.025 c | 1.083 ± 0.025 d |
| 18 | Vicenin-2 (apigenin 6,8-C-dihexoside) isomer d | <LOQ | 0.056 ± 0.006 e | 0.056 ± 0.002 e | 0.041 ± 0.001 b | 0.052 ± 0.003 de | 0.051 ± 0.001 de | 0.049 ± 0.004 cd | 0.007 ± 0.001 a | 0.043 ± 0.003 bc | 0.050 ± 0.001 de |
| 19 | Vicenin-2 (apigenin 6,8-C-dihexoside) isomer e | 0.025 ± 0.001 f | 0.020 ± 0.001 e | <LOQ | 0.019 ± 0.001 de | 0.009 ± 0.001 b | 0.017 ± 0.002 cd | <LOQ | <LOQ | 0.014 ± 0.001 c | 0.001 ± 0.000 a |
| 26 | Vicenin-2 (apigenin 6,8-C-dihexoside) isomer f | <LOQ | 0.087 ± 0.006 e | 0.075 ± 0.005 d | 0.007 ± 0.001 b | 0.074 ± 0.003 d | 0.045 ± 0.000 c | 0.075 ± 0.002 d | <LOQ | 0.072 ± 0.002 d | 0.006 ± 0.001 a |
| 21 | (Vitexin) Apigenin 8-C-glucoside isomer a | 0.541 ± 0.002 e | 0.472 ± 0.003 d | 0.521 ± 0.031 e | 0.177 ± 0.002 b | 0.485 ± 0.010 d | 0.355 ± 0.013 c | 0.461 ± 0.015 d | <LOQ | 0.456 ± 0.017 d | 0.124 ± 0.003 a |
| 22 | (Vitexin) Apigenin 8-C-glucoside isomer b | 0.270 ± 0.016 f | 0.142 ± 0.012 de | 0.154 ± 0.000 e | 0.044 ± 0.002 a | 0.138 ± 0.004 de | 0.086 ± 0.006 b | 0.126 ± 0.001 cd | <LOQ | 0.121 ± 0.004 c | 0.048 ± 0.001 a |
| Flavonols and derivatives | |||||||||||
| 24 | Quercetin rutinoside (rutin) | 0.673 ± 0.023 g | 0.424 ± 0.007 e | 0.467 ± 0.006 f | 0.041 ± 0.001 a | 0.390 ± 0.014 d | 0.154 ± 0.004 b | 0.350 ± 0.006 c | <LOQ | 0.363 ± 0.005 c | 0.033 ± 0.004 a |
| 25 | Quercetin 3-O-β-D-glucoside/Isoquercetin | 1.347 ± 0.086 d | 2.491 ± 0.003 g | 2.144 ± 0.105 f | 0.844 ± 0.016 c | 2.010 ± 0.047 e | 1.272 ± 0.018 d | 1.975 ± 0.069 e | 0.071 ± 0.017 a | 2.087 ± 0.027 ef | 0.660 ± 0.041 b |
| 27 | Quercetin hydroxy-methylglutaroyl galactoside isomer a | <LOQ | 0.126 ± 0.012 c | 0.123 ± 0.004 c | <LOQ | 0.113 ± 0.003 bc | 0.031 ± 0.001 a | 0.105 ± 0.004 b | <LOD | 0.103 ± 0.004 b | <LOQ |
| 29 | Quercetin 3-O-malonylglucoside | 0.353 ± 0.002 f | 0.109 ± 0.008 b | 0.192 ± 0.005 e | <LOQ | 0.177 ± 0.001 d | 0.044 ± 0.000 a | 0.131 ± 0.005 c | <LOD | 0.139 ± 0.006 c | <LOQ |
| 30 | Quercetin 3-O-acetyl glucoside isomer a | 3.973±0.322 e | 2.189 ± 0.180 c | 3.021 ± 0.110 d | 0.029 ± 0.001 a | 2.821 ± 0.013 d | 1.054 ± 0.023 b | 2.297 ± 0.080 c | <LOQ | 2.420 ± 0.071 c | 0.090 ± 0.011 a |
| 32 | Quercetin 3-O-acetyl glucoside isomer b | 0.619 ± 0.038 c | 0.933 ± 0.105 d | 0.688 ± 0.042 c | <LOQ | 0.630 ± 0.030 c | 0.288 ± 0.003 b | 0.627 ± 0.013 c | <LOQ | 0.632 ± 0.013 c | 0.013 ± 0.001 a |
| 33 | Quercetin 3-O-acetyl glucoside isomer c | 0.047 ± 0.003 b | 0.109 ± 0.015 e | 0.100 ± 0.005 de | <LOQ | 0.088 ± 0.001 d | 0.007 ± 0.001 a | 0.068 ± 0.003 c | <LOD | 0.064 ± 0.003 c | <LOQ |
| 34 | Quercetin 3-O-acetyl glucoside isomer d | <LOD | 0.042 ± 0.002 b | 0.095 ± 0.005 f | <LOQ | 0.084 ± 0.004 e | 0.016 ± 0.001 a | 0.072 ± 0.004 d | <LOD | 0.055 ± 0.003 c | <LOQ |
| 42 | Quercetin | <LOQ | 0.130 ± 0.012 c | 0.136 ± 0.004 c | 0.048 ± 0.000 b | 0.132 ± 0.002 c | 0.170 ± 0.002 d | 0.129 ± 0.004 c | 0.008 ± 0.001 a | 0.132 ± 0.005 c | <LOD |
| 40 | Quercetin derivative | <LOQ | 0.069 ± 0.008 d | 0.048 ± 0.003 b | <LOD | 0.047 ± 0.002 b | 0.020 ± 0.001 a | 0.047 ± 0.002 b | <LOD | 0.058 ± 0.000 c | <LOQ |
| 28 | Kaempferol 3-O-glucoside/Kaempferol 3-O-hexose/Astragalin | <LOQ | 0.569 ± 0.031 h | 0.333 ± 0.021 b | 0.527 ± 0.012 g | 0.380 ± 0.016 c | 0.521 ± 0.002 f | 0.454 ± 0.011 de | 0.010 ± 0.001 e | 0.421 ± 0.001 d | 0.482 ± 0.023 e |
| 35 | Kaempferol 3-O-acetyl glucoside isomer a | 2.051 ± 0.167 e | 1.198 ± 0.024 c | 1.719 ± 0.096 d | 0.020 ± 0.000 a | 1.617 ± 0.076 d | 0.605 ± 0.011 b | 1.305 ± 0.033 c | <LOQ | 1.328 ± 0.024 c | 0.078 ± 0.006 a |
| 36 | Kaempferol 3-O-acetyl glucoside isomer b | <LOQ | 0.002 ± 0.000 a | 0.011 ± 0.001 c | <LOD | 0.008 ± 0.001 b | <LOQ | 0.003 ± 0.001 a | <LOD | 0.004 ± 0.001 a | <LOQ |
| 38 | Kaempferol 3-O-acetyl glucoside isomer c | 0.187 ± 0.002 b | 0.635 ± 0.029 e | 0.545 ± 0.001 d | <LOQ | 0.529 ± 0.031 cd | 0.197 ± 0.002 b | 0.532 ± 0.008 cd | <LOQ | 0.504 ± 0.006 c | 0.012 ± 0.001 a |
| 39 | Kaempferol 3-O-acetyl glucoside isomer d | <LOQ | 0.069 ± 0.004 e | 0.054 ± 0.001 d | <LOQ | 0.046 ± 0.004 c | <LOQ | 0.036 ± 0.001 b | <LOD | 0.022 ± 0.002 a | <LOQ |
| 43 | Kaempferol | <LOQ | 0.037 ± 0.001 d | 0.031 ± 0.001 c | 0.030 ± 0.000 c | 0.030 ± 0.002 c | 0.025 ± 0.000 ab | 0.026 ± 0.000 b | <LOQ | 0.023 ± 0.000 a | <LOQ |
| 31 | Isorhamnetin 3-O-glucoside | 0.009 ± 0.000 a | 0.096 ± 0.008 c | 0.043 ± 0.004 a | <LOQ | 0.042 ± 0.003 a | 0.017 ± 0.001 b | 0.041 ± 0.000 a | <LOQ | 0.042 ± 0.002 a | <LOQ |
| 37 | Isorhamnetin 3-acetyl-glucoside isomer a | 0.237 ± 0.010 c | 0.044 ± 0.005 a | 0.081 ± 0.003 b | <LOQ | 0.072 ± 0.003 b | <LOQ | 0.047 ± 0.003 a | <LOD | 0.049 ± 0.001 a | <LOQ |
| 41 | Isorhamnetin 3-acetyl-glucoside isomer b | <LOQ | 0.018 ± 0.001 c | 0.008 ± 0.000 b | <LOD | 0.007 ± 0.001 b | <LOQ | 0.008 ± 0.000 b | <LOD | 0.003 ± 0.000 a | <LOQ |
| Lignans | |||||||||||
| 20 | Isolariciresinol glycoside (lariciresinol glycoside) isomer a | 0.010 ± 0.001 a | 0.050 ± 0.001 f | 0.046 ± 0.001 e | 0.033 ± 0.000 c | 0.046 ± 0.001 e | <LOD | 0.044 ± 0.001 e | 0.028 ± 0.000 b | 0.044 ± 0.001 e | 0.039 ± 0.002 d |
| 23 | Isolariciresinol glycoside (lariciresinol glycoside) isomer b | 0.016 ± 0.001 a | 0.037 ± 0.001 f | 0.037 ± 0.001 ef | 0.032 ± 0.000 c | 0.036 ± 0.000 e | 0.031 ± 0.000 c | 0.030 ± 0.000 b | <LOD | 0.034 ± 0.000 d | <LOD |
| Compound | Dried Leaves | Autoclaved Leaves | Rhizopus oryzae CBS 372.63 | Rhizopus oligosporus ATCC 64063 | Aspergillus oryzae CBS 673.92 | Neurospora intermedia CBS 131.92 | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| - | 3 Days | 8 Days | 3 Days | 8 Days | 3 Days | 8 Days | 3 Days | 8 Days | ||
| Essential amino acids | ||||||||||
| Isoleucine | 1.22 ± 0.02 b | 1.27 ± 0.01 bc | 1.04 ± 0.04 a | 1.06 ± 0.02 a | 1.30 ± 0.11 c | 1.22 ± 0.01 bc | 1.21 ± 0.02 b | 1.23 ± 0.01 bc | 1.52 ± 0.08 d | 1.54 ± 0.07 d |
| Leucine | 2.31 ± 0.03 ab | 2.42 ± 0.03 bc | 1.83 ± 0.08 a | 1.82 ± 0.03 a | 2.49 ± 0.22 bc | 2.35 ± 0.01 c | 2.19 ± 0.05 b | 2.19 ± 0.04 b | 2.89 ± 0.15 d | 2.83 ± 0.14 d |
| Lysine | 1.56 ± 0.02 f | 1.32 ± 0.02 cd | 1.10 ± 0.04 a | 1.05 ± 0.02 a | 1.37 ± 0.12 de | 1.23 ± 0.01 b | 1.27 ± 0.02 bc | 1.25 ± 0.04 bc | 1.63 ± 0.08 f | 1.43 ± 0.06 e |
| Methionine | 0.41 ± 0.06 a | 0.62 ± 0.02 ef | 0.53 ± 0.01 bcd | 0.48 ± 0.02 ab | 0.62 ± 0.01 ef | 0.60 ± 0.01 de | 0.57 ± 0.07 cde | 0.48 ± 0.13 abc | 0.75 ± 0.01 g | 0.68 ± 0.08 fg |
| Phenylalanine | 1.60 ± 0.02 c | 1.64 ± 0.02 c | 1.29 ± 0.05 a | 1.26 ± 0.02 a | 1.65 ± 0.15 c | 1.56 ± 0.02 bc | 1.49 ± 0.04 b | 1.47 ± 0.03 b | 1.95 ± 0.11 d | 1.91 ± 0.09 d |
| Threonine | 1.26 ± 0.01 cd | 1.30 ± 0.02 d | 1.00 ± 0.04 a | 0.99 ± 0.02 a | 1.27 ± 0.11 cd | 1.20 ± 0.01 bc | 1.20 ± 0.02 bc | 1.17 ± 0.01 b | 1.58 ± 0.08 e | 1.58 ± 0.08 e |
| Valine | 1.51 ± 0.02 bc | 1.59 ± 0.02 c | 1.23 ± 0.05 a | 1.26 ± 0.02 a | 1.54 ± 0.14 bc | 1.46 ± 0.01 b | 1.48 ± 0.03 b | 1.50 ± 0.01 bc | 1.85 ± 0.10 d | 1.87 ± 0.08 d |
| Histidine | 0.64 ± 0.01 d | 0.65 ± 0.01 d | 0.61 ± 0.03 abc | 0.57 ± 0.01 a | 0.70 ± 0.06 e | 0.65 ± 0.01 d | 0.62 ± 0.01 bcd | 0.58 ± 0.01 ab | 0.85 ± 0.04 g | 0.78 ± 0.04 f |
| Nonessential amino acids | ||||||||||
| Alanine | 1.79 ± 0.01 d | 1.86 ± 0.02 d | 1.53 ± 0.07 b | 1.64 ± 0.03 c | 1.64 ± 0.14 bc | 1.54 ± 0.01 ab | 1.59 ± 0.03 abc | 1.60 ± 0.01 abc | 1.97 ± 0.10 e | 2.01 ± 0.10 e |
| Arginine | 1.66 ± 0.01 d | 1.65 ± 0.02 cd | 1.34 ± 0.05 ab | 1.24 ± 0.02 a | 1.66 ± 0.15 d | 1.56 ± 0.01 c | 1.42 ± 0.02 b | 1.40 ± 0.01 b | 2.25 ± 0.11 f | 1.89 ± 0.09 e |
| Aspartic acid | 2.57 ± 0.03 bc | 2.67 ± 0.03 bc | 2.36 ± 0.10 a | 2.22 ± 0.04 a | 2.71 ± 0.23 c | 2.53 ± 0.02 b | 2.53 ± 0.04 b | 2.53 ± 0.02 b | 3.17 ± 0.16 d | 3.08 ± 0.16 d |
| Cysteine | 0.25 ± 0.04 a | 0.31 ± 0.01 b | 0.39 ± 0.00 d | 0.37 ± 0.01 cd | 0.34 ± 0.00 bc | 0.34 ± 0.00 bc | 0.34 ± 0.04 bc | 0.30 ± 0.07 ab | 0.46 ± 0.00 e | 0.45 ± 0.05 e |
| Glutamic acid | 3.64 ± 0.03 a | 3.72 ± 0.04 a | 3.64 ± 0.15 a | 3.57 ± 0.06 a | 3.96 ± 0.34 b | 3.73 ± 0.03 ab | 3.77 ± 0.06 ab | 3.76 ± 0.04 ab | 4.75 ± 0.25 c | 4.83 ± 0.24 c |
| Glycine | 1.38 ± 0.02 bc | 1.46 ± 0.01 c | 1.26 ± 0.06 a | 1.22 ± 0.02 a | 1.55 ± 0.13 d | 1.46 ± 0.01 c | 1.35 ± 0.02 b | 1.31 ± 0.01 ab | 1.81 ± 0.09 e | 1.79 ± 0.09 e |
| Serine | 1.33 ± 0.01 cd | 1.37 ± 0.01 d | 1.06 ± 0.04 a | 1.04 ± 0.02 a | 1.25 ± 0.11 bc | 1.19 ± 0.01 b | 1.22 ± 0.02 b | 1.19 ± 0.01 b | 1.55 ± 0.08 e | 1.56 ± 0.08 e |
| Tyrosine | 1.02 ± 0.01 b | 1.01 ± 0.01 b | 0.90 ± 0.03 a | 0.87 ± 0.01 a | 1.12 ± 0.10 c | 1.04 ± 0.01 bc | 1.10 ± 0.06 c | 1.05 ± 0.06 bc | 1.45 ± 0.07 d | 1.40 ± 0.07 d |
| Proline | 1.32 ± 0.01 cd | 1.37 ± 0.02 d | 1.07 ± 0.05 a | 1.00 ± 0.02 a | 1.29 ± 0.12 cd | 1.20 ± 0.01 b | 1.28 ± 0.02 bc | 1.29 ± 0.02 cd | 1.57 ± 0.09 e | 1.54 ± 0.08 e |
| Total g/100 g | 25.47 ± 0.23 bcd | 26.23 ± 0.26 cd | 22.17 ± 0.88 a | 21.67 ± 0.38 a | 26.45 ± 2.22 d | 24.85 ± 0.18 bc | 24.63 ± 0.49 b | 24.29 ± 0.17 b | 32.02 ± 1.58 e | 31.20 ± 1.59 e |
| Compound | Dried Leaves | Autoclaved Leaves | Rhizopus oryzae CBS 372.63 | Rhizopus oligosporus ATCC 64063 | Aspergillus oryzae CBS 673.92 | Neurospora intermedia CBS 131.92 | RSP1 | RSP2 | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| - | 3 Days | 8 Days | 3 Days | 8 Days | 3 Days | 8 Days | 3 Days | 8 Days | ||||
| Histidine | 2.52 ± 0.03 abc | 2.47 ± 0.04 a | 2.73 ± 0.12 d | 2.65 ± 0.06 bcd | 2.64 ± 0.21 bcd | 2.62 ± 0.02 bcd | 2.50 ± 0.05 ab | 2.38 ± 0.04 a | 2.67 ± 0.13 cd | 2.51 ± 0.12 ab | 2.0 | 1.6 |
| Isoleucine | 4.78 ± 0.08 a | 4.83 ± 0.05 ab | 4.67 ± 0.19 a | 4.87 ± 0.08 ab | 4.90 ± 0.43 ab | 4.91 ± 0.03 ab | 4.93 ± 40.10 ab | 5.07 ± 0.05 b | 4.75 ± 0.24 a | 4.91 ± 0.22 ab | 3.2 | 3.0 |
| Leucine | 9.08 ± 0.10 cd | 9.22 ± 0.10 cd | 8.24 ± 0.34 a | 8.41 ± 0.14 ab | 9.42 ± 0.84 cd | 9.44 ± 0.06 d | 8.91 ± 0.21 bc | 8.99 ± 0.15 cd | 9.02 ± 0.47 cd | 9.08 ± 0.43 cd | 6.6 | 6.1 |
| Lysine | 6.14 ± 0.07 d | 5.05 ± 0.06 bc | 4.97 ± 0.19 bc | 4.85 ± 0.09 ab | 5.16 ± 0.44 c | 4.96 ± 0.03 bc | 5.16 ± 0.09 c | 5.13 ± 0.17 bc | 5.09 ± 0.25 bc | 4,56 ± 0.21 a | 5.7 | 4.8 |
| Methionine + Cysteine | 2.59 ± 0.38 a | 3.56 ± 0.12 bc | 4.15 ± 0.05 d | 3.91 ± 0.13 cd | 3.61 ± 0.05 bc | 3.78 ± 0.04 cd | 3.69 ± 0.44 bcd | 3.22 ± 0.84 b | 3.78 ± 0.03 cd | 3.65 ± 0.41 bcd | 2.7 | 2.3 |
| Phenylalanine + Tyrosine | 10.27 ± 0.07 abc | 10.11 ± 0.12 abc | 9.88 ± 0.39 ab | 9.85 ± 0.13 a | 10.46 ± 0.94 abc | 10.47 ± 0.10 abc | 10.51 ± 0.39 bc | 10.38 ± 0.37 abc | 10.65 ± 0.56 c | 10.63 ± 0.54 c | 5.2 | 4.1 |
| Threonine | 4.94 ± 0.04 c | 4.96 ± 0.07 c | 4.50 ± 0.19 a | 4.59 ± 0.07 ab | 4.80 ± 0.41 bc | 4.83 ± 0.03 bc | 4.86 ± 0.08 c | 4.83 ± 0.06 bc | 4.93 ± 0.25 c | 5.06 ± 0.25 c | 3.1 | 2.5 |
| Valine | 5.93 ± 0.09 bc | 6.04 ± 0.07 bc | 5.56 ± 0.23 a | 5.82 ± 0.10 ab | 5.82 ± 0.53 ab | 5.86 ± 0.05 abc | 5.99 ± 0.10 bc | 6.18 ± 0.05 c | 5.79 ± 0.31 ab | 6.00 ± 0.27 bc | 4.3 | 4.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Starzyńska-Janiszewska, A.; Stodolak, B.; Fernández-Fernández, C.; Mickowska, B.; Verardo, V.; Gómez-Caravaca, A.M. Phenolic Profile, Antioxidant Activity and Amino Acid Composition of Moringa Leaves Fermented with Edible Fungal Strains. Foods 2022, 11, 3762. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11233762
Starzyńska-Janiszewska A, Stodolak B, Fernández-Fernández C, Mickowska B, Verardo V, Gómez-Caravaca AM. Phenolic Profile, Antioxidant Activity and Amino Acid Composition of Moringa Leaves Fermented with Edible Fungal Strains. Foods. 2022; 11(23):3762. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11233762
Chicago/Turabian StyleStarzyńska-Janiszewska, Anna, Bożena Stodolak, Carmen Fernández-Fernández, Barbara Mickowska, Vito Verardo, and Ana María Gómez-Caravaca. 2022. "Phenolic Profile, Antioxidant Activity and Amino Acid Composition of Moringa Leaves Fermented with Edible Fungal Strains" Foods 11, no. 23: 3762. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11233762