
Comparative Peptidomics Analysis of Milk Fermented by Lactobacillus helveticus
by
Shuman Gao
1,2, 
Yang Jiang
1,2,
Xinyi Zhang
1,2,
Shumao Cui
1,2,
Xiaoming Liu
2,3,*,
Jianxin Zhao
1,2,
Hao Zhang
1,2,4 and
Wei Chen
1,2,5 1
State Key Laboratory of Food Science and Technology, Jiangnan University, Wuxi 214122, China
2
School of Food Science and Technology, Jiangnan University, Wuxi 214122, China
3
International Joint Research Laboratory for Pharmabiotics & Antibiotic Resistance, Jiangnan University, Wuxi 214122, China
4
Wuxi Translational Medicine Research Center and Jiangsu Translational Medicine Research Institute Wuxi Branch, Wuxi 214122, China
5
National Engineering Research Centre for Functional Food, Wuxi 214122, China
*
Author to whom correspondence should be addressed.
Foods 2022, 11(23), 3885; https://0-doi-org.brum.beds.ac.uk/10.3390/foods11233885
Submission received: 14 October 2022
/
Revised: 21 November 2022
/
Accepted: 25 November 2022
/
Published: 1 December 2022
(This article belongs to the Special Issue Milk and Dairy Products: Linking the Chemistry, Structure, Processing, and Food Properties)
Abstract
:Lactobacillus helveticus is one of the commonly used starter cultures for manufacturing various fermented dairy products. However, only a few studies have explored the cleavage region preference of L. helveticus with different cell envelope proteinase (CEP) genes. In the present study, we profiled the peptide composition of milk samples fermented by three different L. helveticus strains by means of peptidomics to illustrate their different proteolysis patterns. The result revealed that the differences in peptide profiles of milk samples fermented by different L. helveticus strains were mainly a result of variations in the peptide patterns of the casein fractions, which were correlated with CEP genotypes. This was mainly reflected in the extensiveness of the hydrolysis region of αS1-casein and the degree of β-casein hydrolysis. Bioactive peptides were mostly derived from the hydrolysis region common to the three L. helveticus strains, and DQHXN-Q32M42 fermentation resulted in the highest diversity and abundance of bioactive peptides and a significant antihypertensive effect in spontaneous hypertension rats.
1. Introduction
Lactobacillus helveticus is one of the commonly used starter cultures in the manufacture of fermented dairy products, such as yogurt, Swiss-type cheeses, and Italian cheeses [1] and has considerable proteolytic capacity [2]. During proteolysis, proteins are usually degraded into oligopeptides (2–20 amino acids) by the cell envelope proteinase (CEP) of lactic acid bacteria (LAB), and are then transported via peptide transport systems into cells for further degradation into shorter peptides or amino acids under the synergistic effect of various intracellular peptidases [3]. Among LAB, L. helveticus possesses the most diverse distribution of CEP genes, with five paralogs (PrtH1, PrtH2, PrtH3, PrtH4, PrtH1 Variant) identified at present [4].
Previous studies have shown variations in the ability of different L. helveticus strains to hydrolyze milk proteins, especially casein, and the milk fermented by L. helveticus strains with different CEP genotype led to peptide heterogeneity [5]. Sadat-Mekmene et al. [6] found that the hydrolysis kinetics of αs1-casein was enhanced in the presence of L. helveticus strains with both PrtH1 and PrtH2. Skrzypczak et al. [7] showed that L. helveticus strains expressing three CEPs exhibit a greater degree of casein hydrolysis than those expressing one or two CEPs. On the other hand, previous research has also indicated that milk fermentation by L. helveticus notably increased the quantity and variety of bioactive peptides, such as angiotensin-I converting enzyme inhibitory (ACE-I) peptides [8], antioxidant peptides [9], anti-inflammatory peptides, and cognitive-improving peptides [10]. Among them, the lactotripeptides, Ile-Pro-Pro (IPP) and Val-Pro-Pro (VPP), and the lactodipeptide Tyr-Pro (YP), which are released from β- and κ-casein and cannot be obtained directly via gastrointestinal digestive enzymes [11,12], have been identified as ACE inhibitors and displayed the ability to reduce blood pressure in vivo [13,14,15,16]. However, few studies have profiled the peptide composition of fermented milk samples by different L. helveticus strains by means of peptidomics.
Recent advances in peptidomics provide us with a tool of characterization of the peptide profiles of fermented milks by various L. helveticus strains with different CEP genotypes, as well as the cleavage region preference of the caseins by L. helveticus strains. Thus, in the present study, the peptide profile of milk fermented by three L. helveticus strains with different CEPs, ATCC15009, DQHXN-Q32M42, and DYNDL36-6, was analyzed. Quantification of the IPP, VPP, and YP was also carried out, and the antihypertensive effect of milk fermented by L. helveticus was evaluated through spontaneously hypertensive rats (SHRs).
2. Materials and Methods
2.1. Lactobacillus helveticus Strains
The three L. helveticus strains used in this study were ATCC15009, DQHXN-Q32M42, and DYNDL36-6. DQHXN-Q32M42 and DYNDL36-6 were isolated from naturally fermented dairy products in China, and had been deposited in the Culture Collection of Food Microorganisms (CCFM) of Jiangnan University (Wuxi, China); ATCC15009 was a type strain isolated from Swiss Emmenthal cheese.
2.2. Analysis of CEP Genes
The blastn tool was used to analyze the CEP genes, with a BLAST identity cutoff of 98%. The sequences of CEP genes were obtained from NCBI, and the accession numbers were as follows: AAD50643 (PrtH1), ABI13574 (PrtH2), AER42337 (PrtH3), AER42338 (PrtH4), and ADX70200.1 (PrtH1-Variant).
2.3. Preparation of Fermented Milk Samples
The preparation method described by Wang et al. [17] was used with moderate modifications. All three strains were sub-cultured three times in MRS medium and then twice in sterile reconstituted skim milk (11% w/w) at 37 °C prior to experimental use. After two washes in Tris-HCl (pH 6.5), 2% cultures with an initial culture concentration of 1–5 × 108 CFU/mL were inoculated with sterile reconstituted skim milk (11% w/w) and incubated for 48 h at 37 °C.
2.4. Preparation of Whey Fractions
The whey fraction was prepared as described previously [18] with adaptations. Briefly, trichloroacetic acid solution (TCA, 10% w/v in water) was added to fermented milk samples until the pH reached 4.6, and the supernatants were harvested after centrifugation at 10,000× g for 10 min at 4 °C. Then filtration with a 0.45 µm nylon syringe filter and ultrafiltration using a membrane with a cut-off value of 10 kDa were performed. The whey fractions were desalted using a C18 SPE column, collected, and dried by centrifugal evaporation at 40 °C for 2 h.
2.5. Identification of Peptide Sequences by UPLC-ESI-MS/MS
A method previously described [19] was followed with minor modification. Samples were analyzed using an EASY nLC 1200 system equipped with an Acclaim PepMapTM RSLC (50 μm × 15 cm, 2 μm, 100 Å). Mobile phase A was an aqueous solution containing 0.1% (v/v) formic acid and 2% (v/v) acetonitrile, while mobile phase B was an aqueous solution containing 0.1% (v/v) formic acid and 90% (v/v) acetonitrile.
The gradient elution procedure was as follows: 6–20% B, 0–40 min; 20–32% B, 40–52 min; 32–80% B, 52–56 min. The flow rate was set to 0.2 μL/min. The eluted peptides were directly ionized in a mass spectrometer (Thermo Scientific, Waltham, MA, USA) at an ion source voltage of 2.3 kV and subjected to a fully continuous MS scan followed by MS/MS scans three times. MS analysis was performed using the positive ion and data dependence analysis mode. The MS spectra were collected from 150–2000 M/Z with a resolution of 60,000. The MS/MS data acquisition range was 180–2000 M/Z, and the scanning time was 0.1 s.
Using the protein of Bos taurus (signal peptide excluded) from the Uniprot database (available online: https://www.uniprot.org/ (accessed on 5 January 2021)) as the comparison data, the original data were compared and retrieved using MaxQuant (1.5.2.8; Max-Planck Institute for Biochemistry, Martinsried, Germany). A non-specific enzyme cleavage mode was used, and the parameter settings for the database search were used as follows: mass tolerance, 0.02 Da; de novo synthesis tolerance,10 ppm; MS/MS deisotope tolerance, 7 ppm; false discovery rate threshold for protein and peptide matching, 1%. Each sample had three replicates; when the same peptide appeared twice, it was considered to be present in the sample. Subsequently, the peptides detected in each sample were uploaded to the Milk Bioactive Peptide Database (MBPDB) (http://mbpdb.nws.oregonstate.edu/, accessed on 4 June 2022) for functional searches [20].
2.6. Peptide Quantification
The IPP, VPP, and YP contents were determined by triple quadrupole linear ion trap liquid chromatography-mass spectrometry (AB Sciex, Framingham, MA, USA), and a suitable targeted mass spectrometry multi-reaction monitoring method was established. The standard samples of these three peptides were diluted into concentration gradients using 50% acetonitrile aqueous solution, and the contents of each peptide in the samples were calculated by external standard calibration.
2.7. Animals and Experimental Design
Sixteen-week-old male SHRs were purchased from the Beijing Vital River Laboratory Animal Technology Co., Ltd. (Beijing, China). All animal procedures were approved by the Ethics Committee of the Jiangnan University, China (JN. No 20210630W0661216 [245]). The rats were housed in an animal facility under specific pathogen-free conditions at the Laboratory Animal Center of the Department of Food Science and Technology, Jiangnan University, Wuxi, China. The rats were kept under the following conditions: 22 ± 1 °C, 55 ± 10% humidity, and a daily 12 h light/dark cycle.
A single intragastric administration experiment was started after one-week acclimatization. Rats were assigned to the vehicle, positive control, and fermented milk sample-treated groups (n = 6 for each group). Fermented milk sample-treated groups were administered a gavage dose of 10 mL/kg bodyweight, the given vehicle group was gavaged with 0.9% saline once daily with an equal volume, and the positive control group was gavaged with 100 mg/kg bodyweight captopril. All animals were provided ad libitum access to food and water. Blood pressure was measured before and 3 h after the gavage administration. After blood pressure readings stabilized, 10–12 additional consecutive readings were averaged.
2.8. Statistical Analysis
Log10 scale transformation was applied to carry out the statistical analysis of peptidomics, and principal component analysis (PCA) and partial least squares discriminant analysis (PLSDA) were performed to characterize the differences in peptidomic profiles in MetaboAnalyst 5.0 (available online: https://www.metaboanalyst.ca (accessed on 12 July 2022)) [21]. The heat maps of the peptidomics data were constructed using Peptigram, a web-based application for peptidomics data visualization [22], and the heatmap of bioactive peptides was drawn using TBtools [23]. Bar plots were constructed using GraphPad Prism 8.0 (GraphPad Software, San Diego, CA, USA). The mean ± standard deviation (SD) was used for the presentation of data, and one-way ANOVA with Tukey’s multiple comparison was carried out. Statistical significance was set at p < 0.05.
3. Results and Discussion
3.1. CEP Genes Distribution in Strains
The distribution of CEP genes in the selected strains is listed in Table 1. Three L. helveticus strains belong to different cell protease genotypes. More specifically, ATCC15009 possesses the prtH3 gene, DYNDL36-6 possesses prtH2 and prtH4 genes, and DQHXN-Q32M42 possesses three genes (prtH2, prtH3, and prtH4), with the genotypes of prtH3 and prtH4 being rarely reported according to the literature [4,18].
3.2. Peptide Profile Analysis
The PCA across the peptides of skim milk and three fermented milk samples is shown in Figure 1a. Clear separation among the four samples was observed. The result indicated that all three L. helveticus strains significantly altered the peptide composition of the fermented milk samples, with variation observed among them. PLSDA revealed the differential peptides among the three fermented samples, with the top 20 shown in Figure 1b, ranging from 9 to 24 amino acids in length. DQHXN-Q32M42 possessed all 20 peptides, and 19 peptides were recorded in the ATCC15009 sample with αS2-casein f(149,162) missing, whereas DYNDL36-6 only possessed 2 out of 20 differential peptides, αS2-casein f(149,162) and αS2-casein f(100,115).
Figure 2a shows great variation in the ability of different L. helveticus strains to generate peptides, with the highest number of peptides (450) present in DQHXN-Q32M42 and only 181 in DYNDL36-6. These peptides mostly originated from β-casein (32%–51%), followed by αS1-casein (14%–30%), κ-casein (13%–15%), and αS2-casein (6%–10%) (Figure 2b), which is in line with the results reported by Fan et al. [24]. However, considering the fact that αS1-casein, β-casein, αS2-casein, and κ-casein (κ-CN) account for 30%, 28%, 8%, and 10% of caseins, respectively [25], β-casein seems to be preferred by the proteases of L. helveticus strains. In total, 65 common peptides were identified among the three samples, with 80% from casein, which was consistent with the percentage of casein among all the bovine proteins. Regarding the unique peptides, DQHXN-Q32M42 had the largest number with 262 peptides, followed by ATCC15009 (152) and DYNDL36-6 (76). The lengths of the peptides varied among the samples, with DQHXN-Q32M42 normally distributed around peptides with chain lengths of 10, and ATCC15009 and DYNDL36-6 around 14–16 (Figure 2c). The results also indicated the presence of a large number of tripeptides in the three fermented samples.
3.3. Correlation between CEP Genotypes and αS1- and β-Casein Cleavage Patterns
Figure 3 shows the proteolysis patterns of the casein fractions with the detected peptides mapped on the αS1- and β-casein sequences. αS1-casein was divided into eight regions, with regions I, IV, and VIII covering the main common sequences (Figure 3a; see Table S1 for specific details of the regions). There were significant differences in the sequence coverage for αS1-casein among the three fermented samples (55% in ATCC15009, 73% in DQHXN-Q32M42, and 37% in DYNDL36-6). In general, αS1-casein was mainly cleaved in regions I, IV, VI, and VIII by three strains, including the N- and C-termini. Sadat-Mekmene et al. [6] showed similar results and proposed that the lack of secondary structure at the N- and C-terminal ends of αS1-casein led to the susceptibility to hydrolysis in these regions. On the other hand, the micellar forms of casein would also influence the accessibility of different regions of caseins to the CEP of the strains.
Overall, DQHXN-Q32M42, the L. helveticus strain with prtH2–3–4, can cleave more regions of αS1-casein than the strain with only prtH3 and that with prtH2–4. The common hydrolytic sections of II and V in αS1-casein were observed for ATCC15009 and DQHXN-Q32M42, which might be related to the possession of prtH3 in these two strains. On the other hand, the regions of VI (131–142) were shared by DQHXN-Q32M42 and DYNDL36-6, which both possessed prtH2 and prtH4. Further research with more strains of various CEP genotypes is necessary to interpret the pattern of cleavage regions.
Regarding β-casein, the peptides from three L. helveticus strains cover most of the sequence of β-casein, except for the N-terminal region (signal peptide excluded) (Figure 3b). All three L. helveticus strains were able to cleave over 70% of the β-casein (84% in ATCC15009, 75% in DQHXN-Q32M42, and 73% in DYNDL36-6). These findings are in agreement with the results of Begunova et al. [26]. Notably, β-casein seemed to be more accessible to cleavage and subsequently more hydrolyzed than the αS1-casein, probably because this molecule is considered to be an intrinsically unstructured protein [27]. The peptides from regions III and V of β-casein were shared by the three samples, with I (3–5, 22–27) being the unique section of ATCC15009. Section II is the common section for ATCC15009 and DYNDL36-6, and section IV is the common section for ATCC15009 and DQHXN-Q32M42.
However, as shown in Figure 2b, the number of peptides (ATCC15009: 176, DQHXN-Q32M42: 185, and DYNDL36-6: 58) derived from β-casein differed significantly. Further analysis indicated that DQHXN-Q32M42, the strain with prtH2–3–4, generated more widely distributed peptides than ATCC15009 (prtH3) and DYNDL36-6 (prtH2–4). For example, Figure 4 shows the distribution of various peptides from β-casein (142–166) among the three fermented milk samples. It can be observed that DQHXN-Q32M42 had more diverse cleavage sites than the other two strains, which was in agreement with the fact that the peptide length of DQHXN-Q32M42 was normally distributed around 10, while those of ATCC15009 and S12 were 14 and 16, respectively. Thus, differences in CEP genes affected the degree of β-casein hydrolysis, which was reflected by the length and number of peptide segments, but not the preference of regions of hydrolysis.
3.4. Bioactive Peptide and Function Confirmation
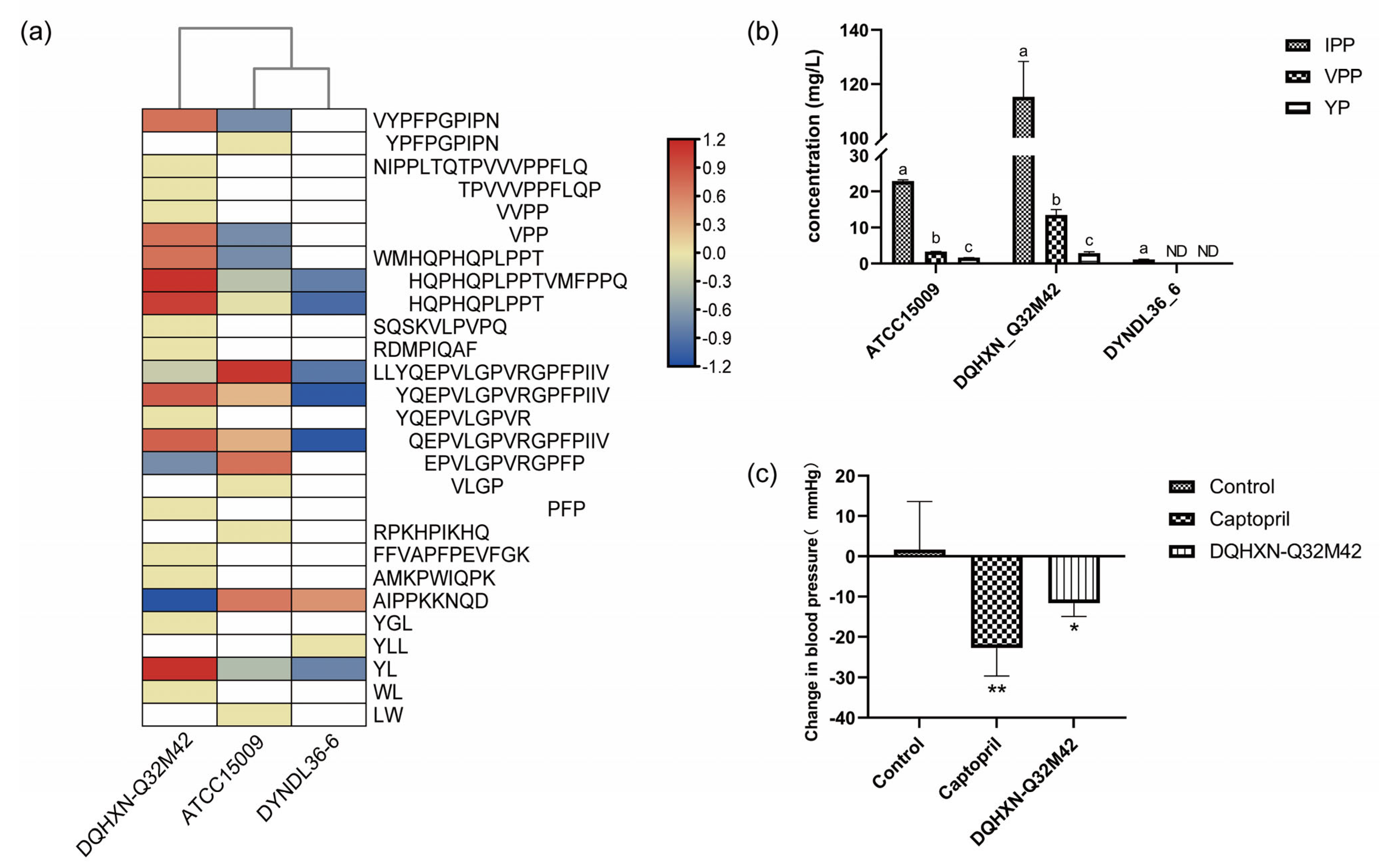
A MBPDB search was performed for the identification of known bioactive sequences, and the results are presented in Table S2. Forty-five bioactive peptides with fourteen functions were identified, mainly including ACE inhibition activity, antioxidant activity, and antibacterial activity. DQHXN-Q32M42 fermentation resulted in the highest diversity and abundance of bioactive peptides, particularly of small bioactive peptides (fewer than 16 amino acid residues), followed by ATCC15009 and DYNDL36-6. These bioactive peptides were mainly from the common sections of αS1-casein (I, II, IV, and VIII) and β-casein (III and V), which might be because these regions can be hydrolyzed extensively (not only by L. helveticus) and have been widely studied. These findings were at odds with the findings of Skrzypczak et al. [28], who revealed that κ-casein proved to be the main source of short peptides released by bacterial enzymes. Because the presence of ACE-I peptides exceeded half of the number of bioactive peptides in the samples, these peptides were selected for further analysis. The distribution of the ACE-I peptides among three samples fermented by L. helveticus strains is shown in Figure 5a. Compared with the other two fermented milk samples, the sample fermented by DQHXN-Q32M42 had a higher diversity and abundance of ACE-I peptides. Even though all the strains could hydrolyze proteins to generate the peptide “LLYQEPVLGPVRGPFPIIV”, “EPVLGPVRGPFP” was only present in the samples of DQHXN-Q32M42 and ATCC15009, with “YQEPVLGPVR” and “PFP” only in the fermented milk of DQHXN-Q32M42. In short, these results suggested that the strain with prtH2–3–4 had a stronger casein hydrolysis capability and was subsequently more prone to generate more and smaller bioactive peptides, which was further supported by the quantitative analysis of typical ACE-I peptides such as IPP and VPP. Quantitative results showed that the DQHXN-Q32M42 sample had the highest IPP and VPP contents among the samples, at 115.269 ± 13.058 and 13.472 ± 1.554 mg/L, respectively (Figure 5b). The milk fermented by DQHXN-Q32M42 had a significant antihypertensive effect (−11.62 ± 3.33 mmHg) in SHR (Figure 5c). Therefore, our study revealed that the existence of different CEPs in L. helveticus results in different peptide profiles of casein and ultimately impacts bioactivity, and that the strain with prtH2–3–4 exhibited greater biological potential among the three strains. Further study with more L. helveticus strains is required to validate the mechanism of cleavage patterns of various CEP genotypes and the biological potential of milk fermented by L. helveticus.
4. Conclusions
Comparative peptidomics analysis revealed different peptide profiles in milk fermented by different L. helveticus strains. These differences were mainly attributed to variations in the peptide profiles of the casein fractions. The αS1- and β-casein hydrolysis patterns were correlated with the CEP genotypes of L. helveticus strains. Among the three L. helveticus strains, DQHXN-Q32M42 with prtH2–3–4, which possessed two unique hydrolysis regions III (50–63) and VI (143–157) of αS1-casein and generated smaller peptides from β-casein, demonstrated the highest casein hydrolysis activities. Bioactive peptides were mostly derived from the hydrolysis region common to the three L. helveticus strains, and DQHXN-Q32M42 fermentation resulted in the highest diversity and abundance of bioactive peptides and a significant antihypertensive effect in SHRs.
Supplementary Materials
The following supporting information can be downloaded at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/foods11233885/s1: Table S1: Partitioning of αS1 and β-casein according to heat map of αS1-casein and β-casein; Table S2: Bioactive peptides identified in the fermented milk samples of Lactobacillus helveticus.
Author Contributions
S.G. conducted the experiment and wrote the manuscript; S.G., X.Z., Y.J. and S.C. analyzed the data; X.L. designed the study and secured the funding; J.Z., H.Z. and W.C. reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The present research work was supported by the National Natural Science Foundation of China (No. 31871829, 31820103010, and 32021005) and the Collaborative Innovation Centre of Food Safety and Quality Control in Jiangsu Province (Jiangsu, China).
Data Availability Statement
The datasets generated for this study are available on request to the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Slattery, L.; O’Callaghan, J.; Fitzgerald, G.F.; Beresford, T.; Ross, R.P. Invited review: Lactobacillus helveticus--a thermophilic dairy starter related to gut bacteria. J. Dairy Sci. 2010, 93, 4435–4454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, M.W.; Tellez, A.M. Lactobacillus helveticus: The proteolytic system. Front. Microbiol. 2013, 4, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y.X.; Li, X.; Liu, H.Y.; Li, Q.M.; Xiao, R.; Dudu, O.E.; Yang, L.; Ma, Y. The impact of multiple-species starters on the peptide profiles of yoghurts. Int. Dairy J. 2020, 106, 104684. [Google Scholar] [CrossRef]
- Broadbent, J.R.; Cai, H.; Larsen, R.L.; Hughes, J.E.; Welker, D.L.; De Carvalho, V.G.; Tompkins, T.A.; Ardo, Y.; Vogensen, F.; De Lorentiis, A.; et al. Genetic diversity in proteolytic enzymes and amino acid metabolism among Lactobacillus helveticus strains. J. Dairy Sci. 2011, 94, 4313–4328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, M.P.; Vogensen, F.K.; Ardö, Y. Variation in caseinolytic properties of six cheese related Lactobacillus helveticus strains. Int. Dairy J. 2009, 19, 661–668. [Google Scholar] [CrossRef]
- Sadat-Mekmene, L.; Jardin, J.; Corre, C.; Molle, D.; Richoux, R.; Delage, M.M.; Lortal, S.; Gagnaire, V. Simultaneous presence of PrtH and PrtH2 proteinases in Lactobacillus helveticus Strains improves breakdown of the pure alphas1-casein. Appl. Environ. Microbiol. 2011, 77, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Skrzypczak, K.W.; Gustaw, W.Z.; Wasko, A.D. Distribution of Cell Envelope Proteinases Genes among Polish Strains of Lactobacillus helveticus. Pol. J. Microbiol. 2018, 67, 203–211. [Google Scholar] [CrossRef] [Green Version]
- Pan, D.; Guo, Y. Optimization of sour milk fermentation for the production of ACE-inhibitory peptides and purification of a novel peptide from whey protein hydrolysate. Int. Dairy J. 2010, 20, 472–479. [Google Scholar] [CrossRef]
- Yang, W.; Hao, X.; Zhang, X.; Zhang, G.; Li, X.; Liu, L.; Sun, Y.; Pan, Y. Identification of antioxidant peptides from cheddar cheese made with Lactobacillus helveticus. LWT 2021, 141, 110866. [Google Scholar] [CrossRef]
- Ohsawa, K.; Nakamura, F.; Uchida, N.; Mizuno, S.; Yokogoshi, H. Lactobacillus helveticus-fermented milk containing lactononadecapeptide (NIPPLTQTPVVVPPFLQPE) improves cognitive function in healthy middle-aged adults: A randomised, double-blind, placebo-controlled trial. Int. J. Food Sci. Nutr. 2018, 69, 369–376. [Google Scholar] [CrossRef]
- Ohsawa, K.; Satsu, H.; Ohki, K.; Enjoh, M.; Takano, T.; Shimizu, M. Producibility and Digestibility of Antihypertensive β-Casein Tripeptides, Val-Pro-Pro and Ile-Pro-Pro, in the Gastrointestinal Tract: Analyses Using an in Vitro Model of Mammalian Gastrointestinal Digestion. J. Agric. Food Chem. 2008, 56, 854–858. [Google Scholar] [CrossRef]
- Foltz, M.; Meynen, E.E.; Bianco, V.; Platerink, C.v.; Koning, T.M.M.G.; Kloek, J. Angiotensin converting enzyme inhibitory peptides from a lactotripeptide-enriched milk beverage are absorbed intact into the circulation. J. Nutr. 2007, 137, 953–958. [Google Scholar] [CrossRef] [Green Version]
- Fekete, A.A.; Givens, D.I.; Lovegrove, J.A. Casein-derived lactotripeptides reduce systolic and diastolic blood pressure in a meta-analysis of randomised clinical trials. Nutrients 2015, 7, 659–681. [Google Scholar] [CrossRef] [Green Version]
- Turpeinen, A.M.; Jarvenpaa, S.; Kautiainen, H.; Korpela, R.; Vapaatalo, H. Antihypertensive effects of bioactive tripeptides-a random effects meta-analysis. Ann. Med. 2013, 45, 51–56. [Google Scholar] [CrossRef]
- Yamamoto, N.; Maeno, M.; Takano, T. Purification and characterization of an antihypertensive peptide from a yogurt-like product fermented by Lactobacillus helveticus CPN4. J. Dairy Sci. 1999, 82, 1388–1393. [Google Scholar] [CrossRef]
- Okamoto, K.; Kawamura, S.; Tagawa, M.; Mizuta, T.; Zahid, H.M.; Nabika, T. Production of an antihypertensive peptide from milk by the brown rot fungus Neolentinus lepideus. Eur. Food Res. Technol. 2020, 246, 1773–1782. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, Y.; Yu, P.; Lee, Y.K.; Liu, X.; Zhao, J.; Zhang, H.; Chen, W. Effect of carbon catabolite repression on lactose and galactose catabolism in Lacticaseibacillus paracasei. Food Biosci. 2021, 40, 100912. [Google Scholar] [CrossRef]
- Raveschot, C.; Cudennec, B.; Deracinois, B.; Fremont, M.; Vaeremans, M.; Dugersuren, J.; Demberel, S.; Drider, D.; Dhulster, P.; Coutte, F.; et al. Proteolytic activity of Lactobacillus strains isolated from Mongolian traditional dairy products: A multiparametric analysis. Food Chem. 2020, 304, 125415. [Google Scholar] [CrossRef]
- Ye, H.J.; Zhang, X.Y.; Jiang, Y.; Guo, M.; Liu, X.M.; Zhao, J.X.; Zhang, H.; Chen, W. Comparative Peptidomics Analysis of Fermented Milk by Lactobacillus delbrueckii ssp. bulgaricus and Lactobacillus delbrueckii ssp. lactis. Foods 2021, 10, 3028. [Google Scholar]
- Nielsen, S.D.; Beverly, R.L.; Qu, Y.; Dallas, D.C. Milk bioactive peptide database: A comprehensive database of milk protein-derived bioactive peptides and novel visualization. Food Chem. 2017, 232, 673–682. [Google Scholar] [CrossRef]
- Pang, Z.; Zhou, G.; Ewald, J.; Chang, L.; Hacariz, O.; Basu, N.; Xia, J. Using MetaboAnalyst 5.0 for LC-HRMS spectra processing, multi-omics integration and covariate adjustment of global metabolomics data. Nat. Protoc. 2022, 17, 1735–1761. [Google Scholar] [CrossRef] [PubMed]
- Manguy, J.; Jehl, P.; Dillon, E.T.; Davey, N.E.; Shields, D.C.; Holton, T.A. Peptigram: A Web-Based Application for Peptidomics Data Visualization. J. Proteome Res. 2017, 16, 712–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Guo, T.; Li, W.; Chen, J.; Li, F.; Wang, C.; Shi, Y.; Li, D.X.-a.; Zhang, S. Isolation and identification of novel casein-derived bioactive peptides and potential functions in fermented casein with Lactobacillus helveticus. Food Sci. Hum. Wellness 2019, 8, 156–176. [Google Scholar] [CrossRef]
- Bhat, M.Y.; Dar, T.A.; Singh, L.R. Chapter 1—Casein Proteins: Structural and Functional Aspects. In Milk Proteins—From Structure to Biological Properties and Health Aspects; Gigli, I., Ed.; IntechOpen: London, UK, 2016; pp. 3–18. [Google Scholar]
- Begunova, A.V.; Savinova, O.S.; Glazunova, O.A.; Moiseenko, K.V.; Rozhkova, I.V.; Fedorova, T.V. Development of Antioxidant and Antihypertensive Properties during Growth of Lactobacillus helveticus, Lactobacillus rhamnosus and Lactobacillus reuteri on Cow’s Milk: Fermentation and Peptidomics Study. Foods 2021, 10, 17. [Google Scholar] [CrossRef]
- Kumosinski, T.F.; Brown, E.M.; Farrell, H.M., Jr. Three-Dimensional Molecular Modeling of Bovine Caseins: An Energy-Minimized β-Casein Structure. J. Dairy Sci. 1993, 76, 931–945. [Google Scholar] [CrossRef]
- Skrzypczak, K.; Gustaw, W.; Szwajgier, D.; Fornal, E.; Wasko, A. kappa-Casein as a source of short-chain bioactive peptides generated by Lactobacillus helveticus. J. Food Sci. Technol. 2017, 54, 3679–3688. [Google Scholar] [CrossRef]
Figure 1.
(a) Principal component analysis for peptidomics of fermented milks by Lactobacillus helveticus. (b) Peptides with the top 20 variable importance in projection (VIP) scores in the fermented milk of L. helveticus according to partial least-squares discriminant analysis.
Figure 1.
(a) Principal component analysis for peptidomics of fermented milks by Lactobacillus helveticus. (b) Peptides with the top 20 variable importance in projection (VIP) scores in the fermented milk of L. helveticus according to partial least-squares discriminant analysis.

Figure 2.
Set analysis of peptidomics data. (a) Venn diagram of the peptidomics of fermented milks by Lactobacillus helveticus. The top figure is a classical Venn diagram, the middle figure shows the number of peptides in every sample, and the bottom figure lists the numbers of common peptides and unique peptide. (b) Peptide source protein distribution map of the identified peptides. (c) Peptide length distribution map.
Figure 2.
Set analysis of peptidomics data. (a) Venn diagram of the peptidomics of fermented milks by Lactobacillus helveticus. The top figure is a classical Venn diagram, the middle figure shows the number of peptides in every sample, and the bottom figure lists the numbers of common peptides and unique peptide. (b) Peptide source protein distribution map of the identified peptides. (c) Peptide length distribution map.

Figure 3.
Heat map of (a) αS1-casein and (b) β-casein. The height of the green bars and the intensity of the green color are proportional to the count of peptides and the sum of the peptide peak intensities overlapping this position, respectively. The count of peptides overlapping a residue and the sum of their intensities is displayed at the top of the figure.
Figure 3.
Heat map of (a) αS1-casein and (b) β-casein. The height of the green bars and the intensity of the green color are proportional to the count of peptides and the sum of the peptide peak intensities overlapping this position, respectively. The count of peptides overlapping a residue and the sum of their intensities is displayed at the top of the figure.

Figure 4.
Heat map of peptides derived from β-casein (142–166).

Figure 5.
Distribution of ACE inhibitory peptides in Lactobacillus helveticus-fermented milk and their blood pressure regulation ability. (a) ACE-I peptides identified in fermented milk samples of L. helveticus. (b) Concentrations of IPP, VPP, and YP in the milks fermented by L. helveticus. Significant differences (One-way ANOVA with Tukey’s multiple comparison, p < 0.05) between groups are indicated with different letters (a, b and c) above the bars. ND means not detected. (c) Effects of single gavage of milks fermented by L. helveticus for blood pressure in early SHR. One-way ANOVA with Tukey’s multiple comparison; * p <0.05, ** p <0.01.
Figure 5.
Distribution of ACE inhibitory peptides in Lactobacillus helveticus-fermented milk and their blood pressure regulation ability. (a) ACE-I peptides identified in fermented milk samples of L. helveticus. (b) Concentrations of IPP, VPP, and YP in the milks fermented by L. helveticus. Significant differences (One-way ANOVA with Tukey’s multiple comparison, p < 0.05) between groups are indicated with different letters (a, b and c) above the bars. ND means not detected. (c) Effects of single gavage of milks fermented by L. helveticus for blood pressure in early SHR. One-way ANOVA with Tukey’s multiple comparison; * p <0.05, ** p <0.01.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Distribution of CEP genes in experimental strains.
| Strains | ATCC15009 | DYNDL36_6 | DQHXN-Q32M42 |
|---|---|---|---|
| PrtH1 | − | − | − |
| PrtH2 | − | + | + |
| PrtH3 | + | − | + |
| PrtH4 | − | + | + |
| PrtH1 Variant | − | − | − |
“+” means that the gene has been identified; “−” means that the gene has not been identified. Genes listed with bold font indicate that the genes were differentially expressed in strains.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gao, S.; Jiang, Y.; Zhang, X.; Cui, S.; Liu, X.; Zhao, J.; Zhang, H.; Chen, W. Comparative Peptidomics Analysis of Milk Fermented by Lactobacillus helveticus. Foods 2022, 11, 3885. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11233885
AMA Style
Gao S, Jiang Y, Zhang X, Cui S, Liu X, Zhao J, Zhang H, Chen W. Comparative Peptidomics Analysis of Milk Fermented by Lactobacillus helveticus. Foods. 2022; 11(23):3885. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11233885
Chicago/Turabian StyleGao, Shuman, Yang Jiang, Xinyi Zhang, Shumao Cui, Xiaoming Liu, Jianxin Zhao, Hao Zhang, and Wei Chen. 2022. "Comparative Peptidomics Analysis of Milk Fermented by Lactobacillus helveticus" Foods 11, no. 23: 3885. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11233885
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.