
Influence of Lactic Acid Bacterium Strains on Changes in Quality, Functional Compounds and Volatile Compounds of Mango Juice from Different Cultivars during Fermentation


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Microbial Analysis and Survival of Viable LAB Cells
2.3. Evaluation of Sensory Attributes
2.4. Determination of Physicochemical Parameters
2.5. Ascorbic Acid Content
2.6. Total Phenol Content (TPC)
2.7. β-Carotene Content
2.8. Antioxidant Activities
2.8.1. 2,2-Diphenyl-1-picrylhydrazyl Radial Scavenging Activity
2.8.2. ABTS-Based Scavenging Activity
2.8.3. Antioxidant Power
2.9. Volatile Compounds
2.10. Statistical Analysis
3. Results and Discussion
3.1. Total Soluble Solids (TSS)
3.2. pH and Titratable Acidity
3.3. Colour Properties
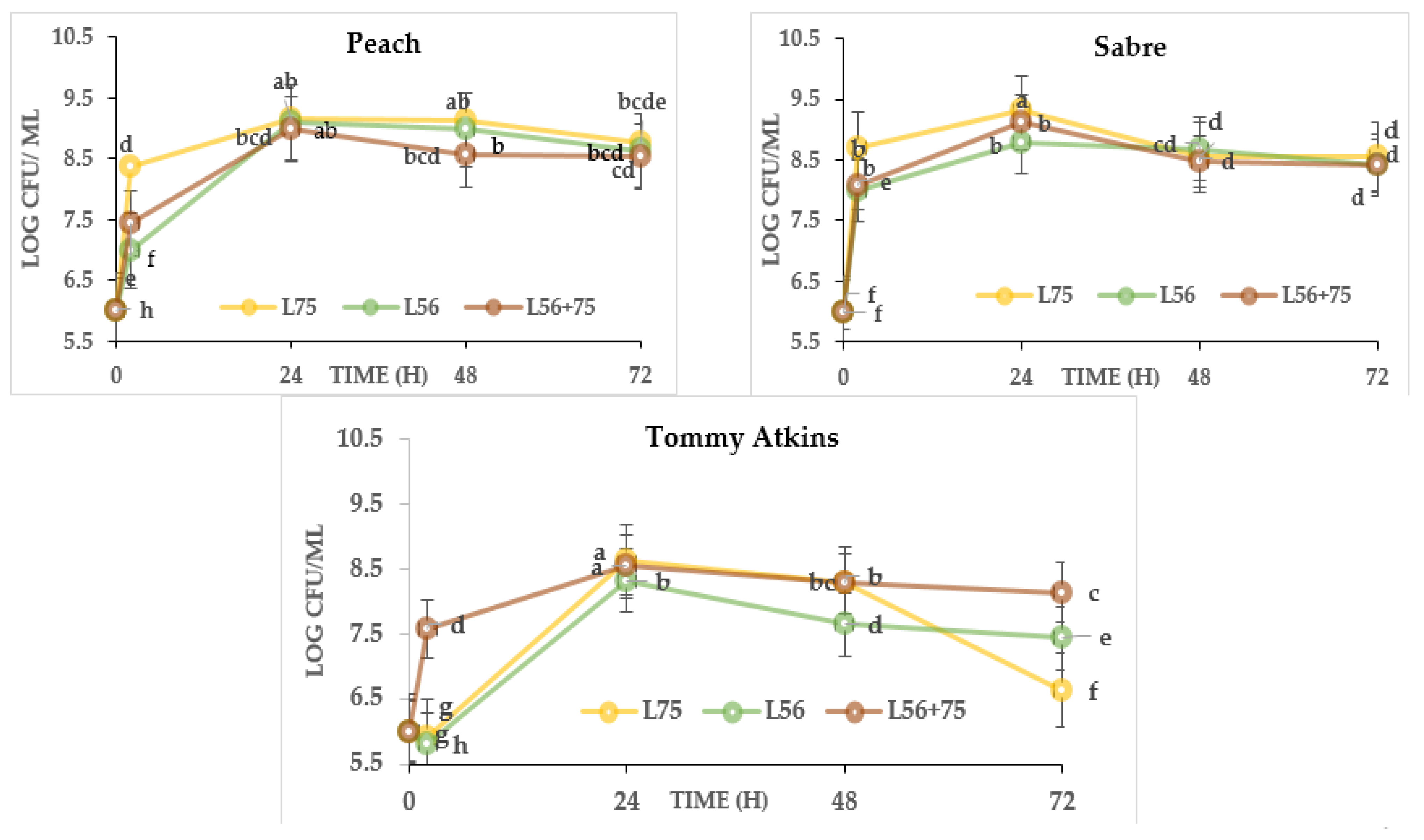
3.4. Microbial Quality and LAB Growth
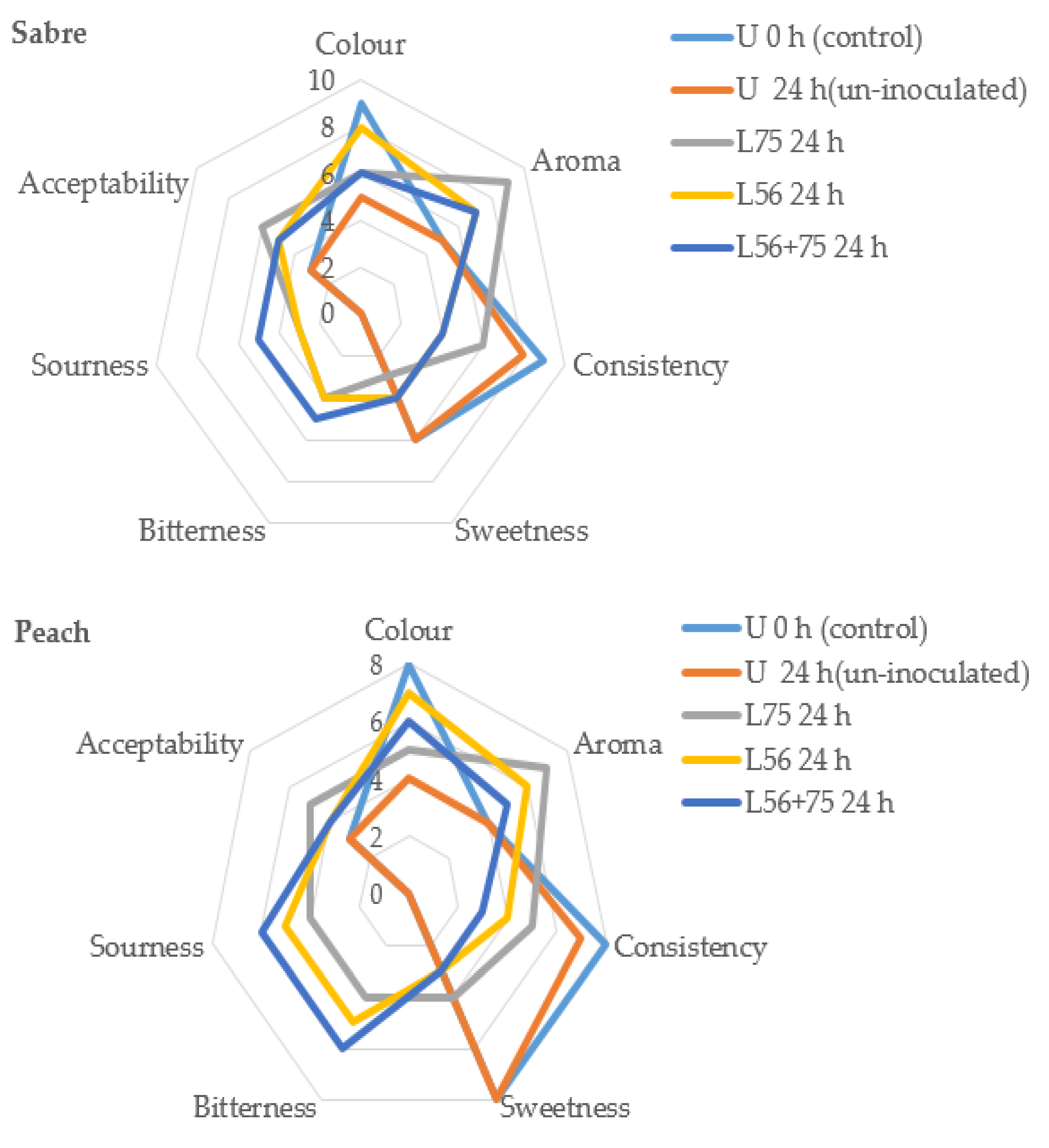
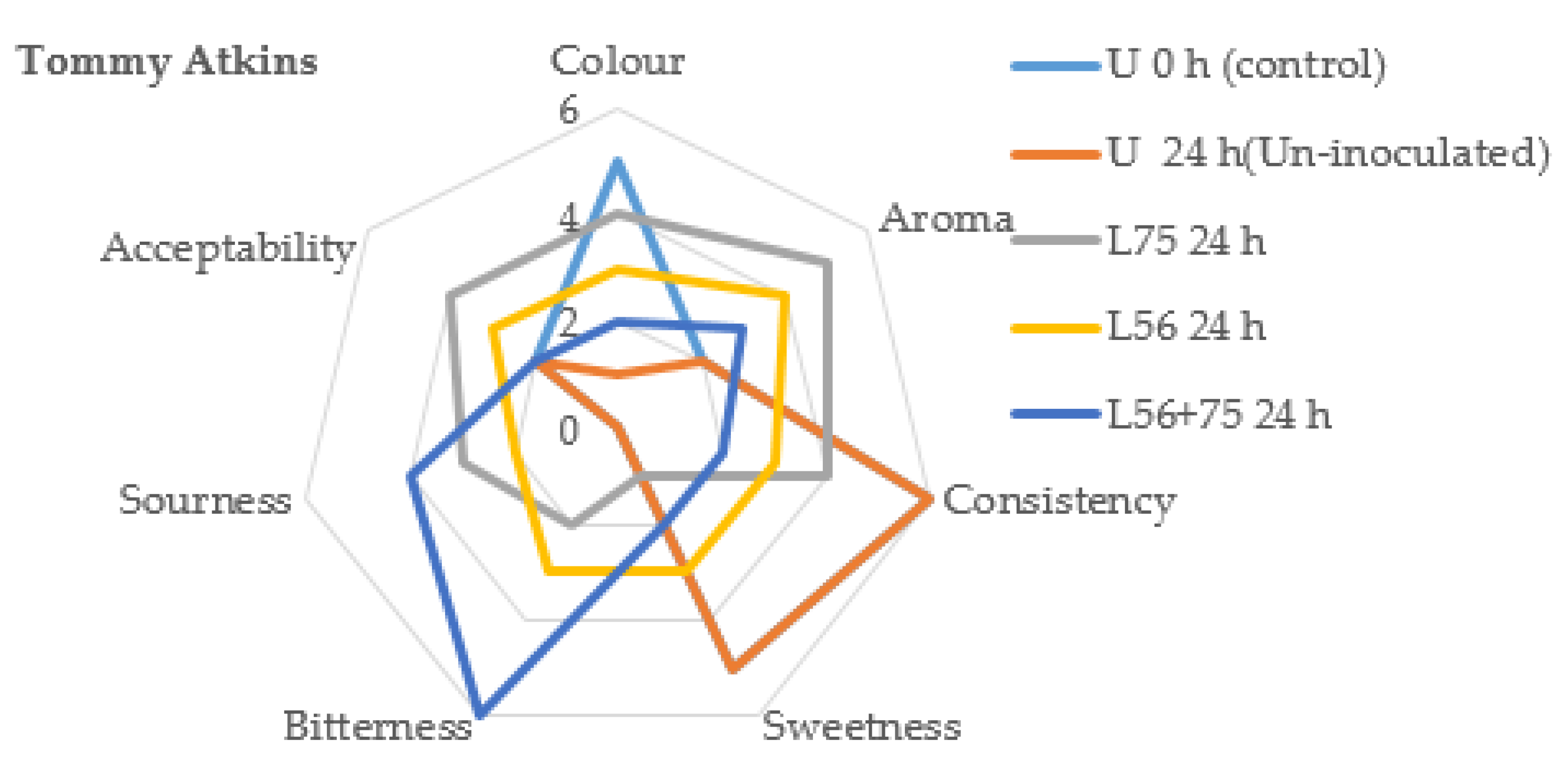
3.5. Sensory Analysis
3.6. Chemical Constituents
3.6.1. Ascorbic Acid
3.6.2. Total Phenol Content
3.6.3. β-Carotene
3.7. Antioxidant Activity
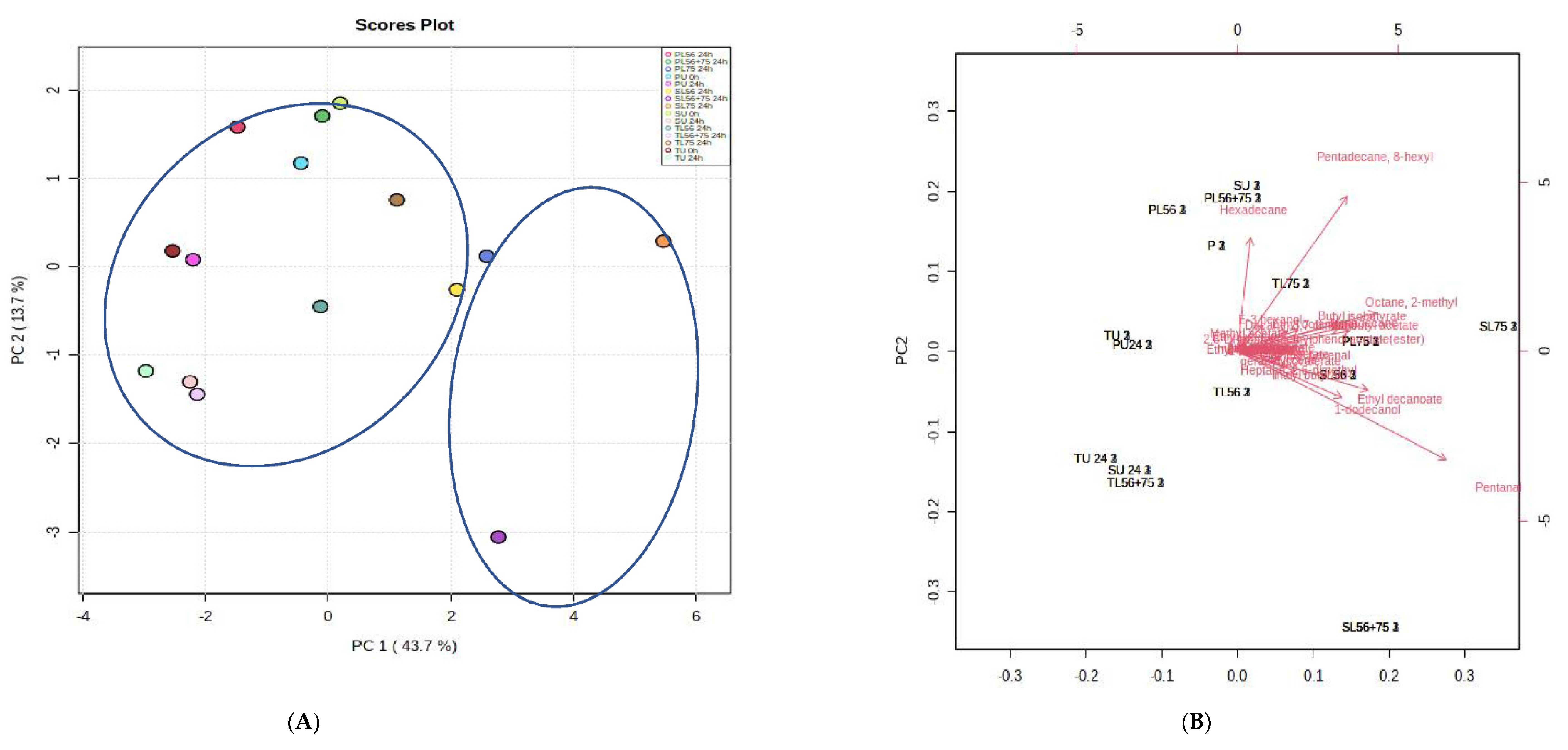
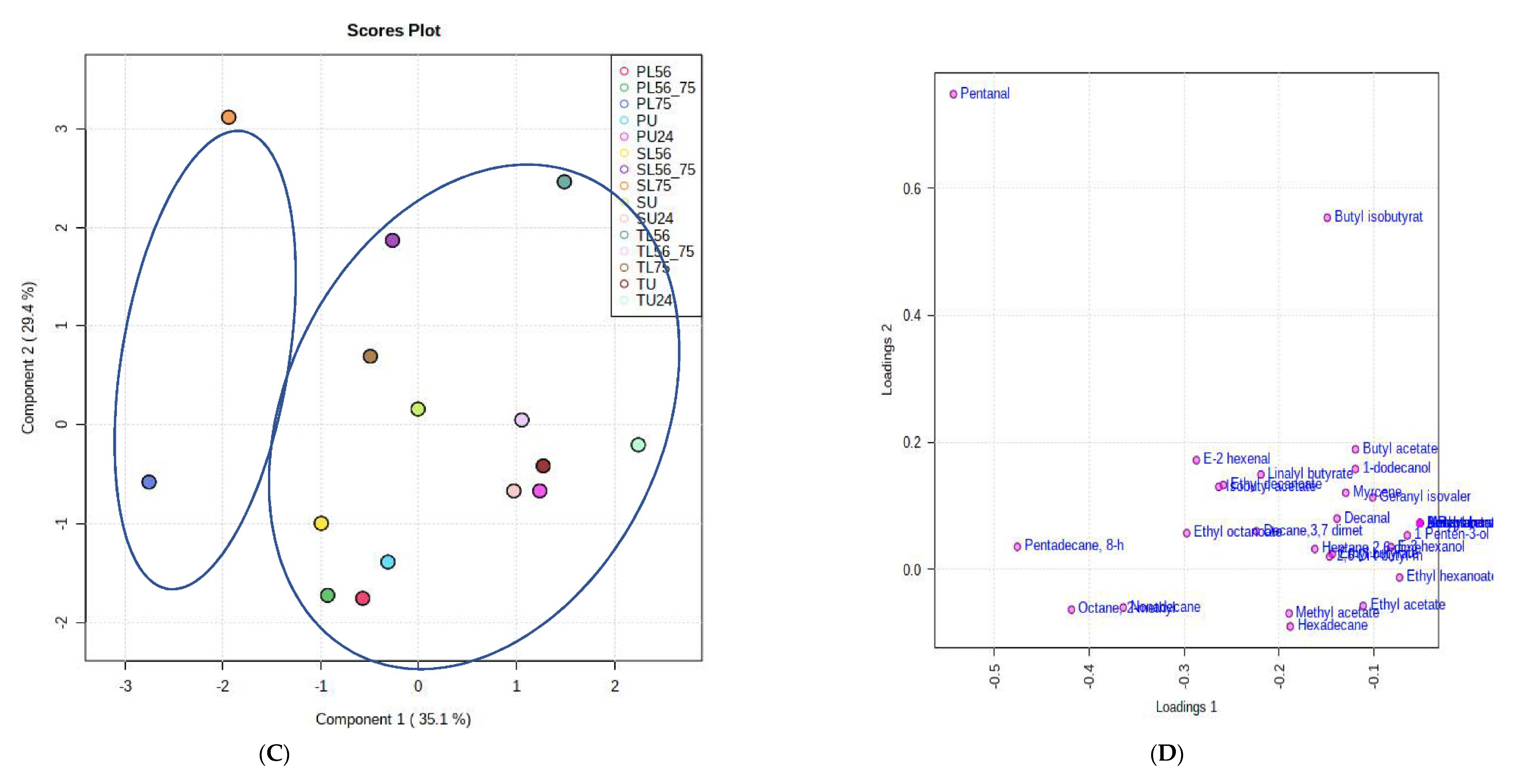
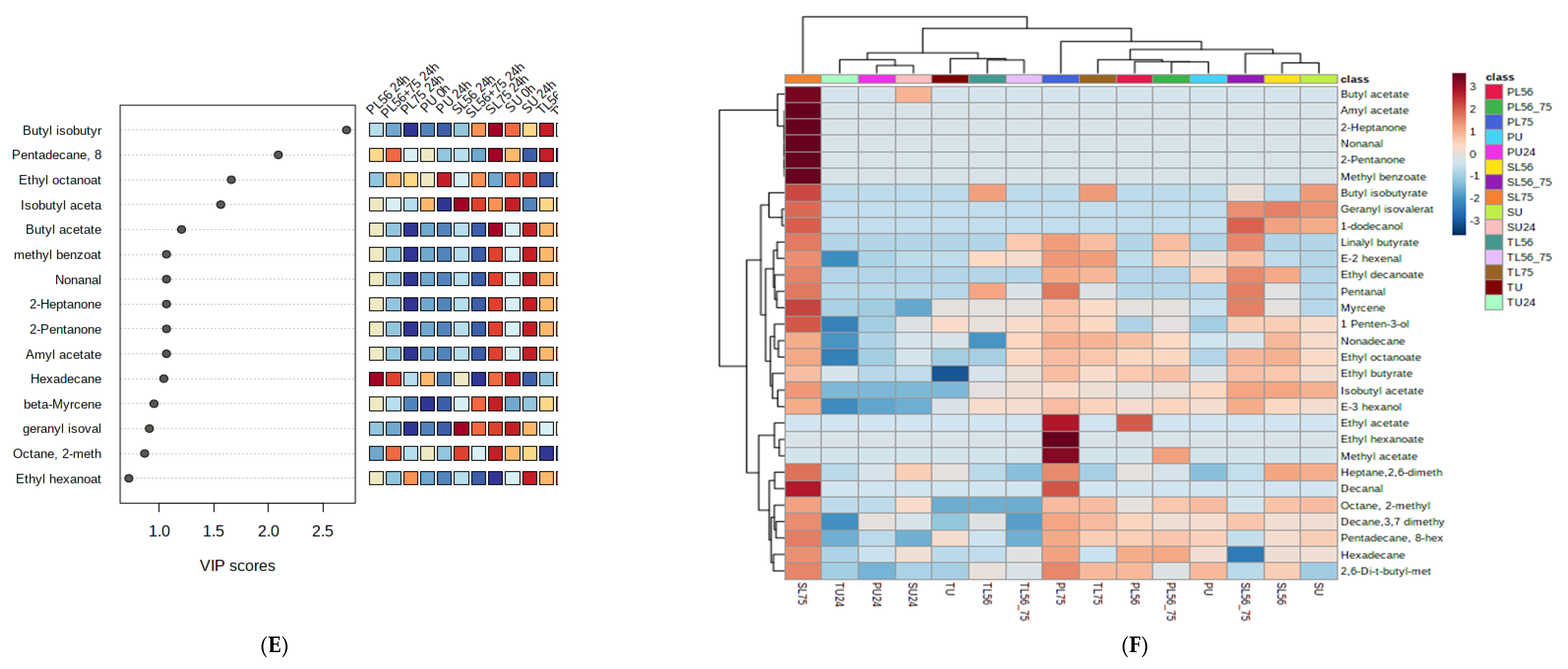
3.8. Effects of Fermentation Process on Volatile Compounds
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Degrain, A.; Manhivi, V.; Remize, F.; Garcia, C.; Sivakumar, D. Effect of lactic acid fermentation on color, phenolic compounds and antioxidant activity in African nightshade. Microorganisms 2020, 8, 1324. [Google Scholar] [CrossRef] [PubMed]
- Fessard, A.; Kapoor, A.; Patche, J.; Assemat, S.; Hoarau, M.; Bourdon, E.; Bahorun, T.; Remize, F. Lactic fermentation as an efficient tool to enhance the antioxidant activity of tropical fruit juices and teas. Microorganisms 2017, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Li, T.; Qi, J.; Jiang, T.; Xu, H.; Lei, H. Effects of lactic acid fermentation-based biotransformation on phenolic profiles, antioxidant capacity and flavor volatiles of apple juice. LWT Food Sci. Technol. 2020, 122, 109064. [Google Scholar] [CrossRef]
- Garcia, C.; Guerin, M.; Souidi, K.; Remize, F. Lactic fermented fruit or vegetable juices: Past, present and future. Beverages 2020, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Arampath, P.C.; Dekker, M. Bulk storage of mango (Mangifera indica L.) and pineapple (Ananas comosus L.) pulp: Effect of pulping and storage temperature on phytochemicals and antioxidant activity. J. Sci. Food Agric. 2019, 99, 5157–5167. [Google Scholar] [CrossRef]
- Abbasi, N.A.; Iqbal, Z.; Maqbool, M.; Hafiz, I.A. Postharvest quality of mango (Mangifera indica L.) fruit as affected by chitosan coating. Pak. J. Bot. 2009, 41, 343–357. [Google Scholar]
- Hashemi, S.M.B.; Jafarpour, D. Fermentation of bergamot juice with Lactobacillus plantarum strains in pure and mixed fermentations: Chemical composition, antioxidant activity and sensorial properties. LWT Food Sci. Technol. 2020, 131, 109803. [Google Scholar] [CrossRef]
- Rakin, M.; Vukasinovic, M.; Siler-Marinkovic, S.; Maksimovic, M. Contribution of lactic acid fermentation to improved nutritive quality vegetable juices enriched with brewer’s yeast autolysate. Food Chem. 2007, 100, 599–602. [Google Scholar] [CrossRef]
- Kumar, B.V.; Sreedharamurthy, M.; Reddy, O.V.S. Probiotication of mango and sapota juices using Lactobacillus plantarum NCDC LP 20. Nutrafoods 2015, 14, 97–106. [Google Scholar] [CrossRef]
- Managa, M.G.; Akinola, S.A.; Remize, F.; Garcia, C.; Sivakumar, D. Physicochemical parameters and bioaccessibility of lactic acid bacteria fermented chayote Leaf (Sechium edule) and pineapple (Ananas comosus) smoothies. Front. Nutr. 2021, 8, 120. [Google Scholar] [CrossRef]
- Mousavi, Z.; Mousavi, S.; Razavi, S.; Emam-Djomeh, Z.; Kiani, H. Fermentation of pomegranate juice by probiotic lactic acid bacteria. World J. Microbiol. Biotechnol. 2011, 27, 123–128. [Google Scholar] [CrossRef]
- European-Commission-Regulation. European Commission Regulation (EC 1441). Microbiological Criteria for Foodstuffs; European Union: Maastrick, The Netherlands, 2007. [Google Scholar]
- Oliveira, A.d.N.; Ramos, A.M.; Minim, V.P.R.; Chaves, J.B.P. Sensory stability of whole mango juice: Influence of temperature and storage time. Food Sci. Technol. 2012, 32, 819–825. [Google Scholar] [CrossRef] [Green Version]
- Reddy, L.V.; Min, J.-H.; Wee, Y.-J. Production of probiotic mango juice by fermentation of lactic acid bacteria. Microbiol. Biotechnol. Lett. 2015, 43, 120–125. [Google Scholar] [CrossRef] [Green Version]
- Al-Sahlany, S.T.; Niamah, A.K. Bacterial viability, antioxidant stability, antimutagenicity and sensory properties of onion types fermentation by using probiotic starter during storage. Nutr. Food Sci. 2022; in press. [Google Scholar] [CrossRef]
- Official-Methods-of-Analysis. Vitamin C (reduced ascorbic acid) in ready-to-feed milk-based infant formula. In AOAC Official Method 985.33; AOAC Official Method: Rockville, MD, USA, 2000. [Google Scholar]
- Panfili, G.; Fratianni, A.; Irano, M. Improved normal-phase high-performance liquid chromatography procedure for the determination of carotenoids in cereals. J. Agric. Food Chem. 2004, 52, 6373–6377. [Google Scholar] [CrossRef]
- Seke, F.; Manhivi, V.E.; Shoko, T.; Slabbert, R.M.; Sultanbawa, Y.; Sivakumar, D. Extraction optimisation, hydrolysis, antioxidant properties and bioaccessibility of phenolic compounds in Natal plum fruit (Carissa macrocarpa). Food Biosci. 2021, 44, 101425. [Google Scholar] [CrossRef]
- Hijaz, F.; Nehela, Y.; Killiny, N. Possible role of plant volatiles in tolerance against huanglongbing in citrus. Plant Signal. Behav. 2016, 11, e1138193. [Google Scholar] [CrossRef] [Green Version]
- Kaprasob, R.; Kerdchoechuen, O.; Laohakunjit, N.; Sarkar, D.; Shetty, K. Fermentation-based biotransformation of bioactive phenolics and volatile compounds from cashew apple juice by select lactic acid bacteria. Process Biochem. 2017, 59, 141–149. [Google Scholar] [CrossRef]
- Mashitoa, F.M.; Akinola, S.A.; Manhevi, V.E.; Garcia, C.; Remize, F.; Slabbert, R.; Sivakumar, D. Influence of Fermentation of Pasteurised Papaya Puree with Different Lactic Acid Bacterial Strains on Quality and bioaccessibility of phenolic compounds during in vitro digestion. Foods 2021, 10, 962. [Google Scholar] [CrossRef]
- Liao, X.-Y.; Guo, L.-Q.; Ye, Z.-W.; Qiu, L.-Y.; Gu, F.-W.; Lin, J.-F. Use of autochthonous lactic acid bacteria starters to ferment mango juice for promoting its probiotic roles. Prep. Biochem. Biotechnol. 2016, 46, 399–405. [Google Scholar] [CrossRef]
- Gong, Y.; Wang, Q.; Ma, S.; Ma, Y.; Meng, Q.; Zhang, Z.; Shi, J. Short-time water immersion inhibits browning of fresh-cut potato by enhancing antioxidant capability and tyrosine scavenging. J. Food Process. Preserv. 2019, 43, e14168. [Google Scholar] [CrossRef]
- Shabala, L.; McMeekin, T.; Budde, B.B.; Siegumfeldt, H. Listeria innocua and Lactobacillus delbrueckii subsp. bulgaricus employ different strategies to cope with acid stress. Int. J. Food Microbiol. 2006, 110, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Hamauzu, Y. Phenolic compounds, ascorbic acid, carotenoids and antioxidant properties of green, red and yellow bell peppers. J. Food Agric. Environ. 2004, 1, 22–27. [Google Scholar] [CrossRef]
- Abourayya, M.; Kassim, N.; El-Sheikh, M.; Rakha, A. Fruit physical and chemical characteristics at maturity stage of Tommy Atkins, Keitt and Kent mango cultivars grown under Nubariya conditions. J. Am. Sci. 2011, 7, 228–233. [Google Scholar]
- Multari, S.; Carafa, I.; Barp, L.; Caruso, M.; Licciardello, C.; Larcher, R.; Tuohy, K.; Martens, S. Effects of Lactobacillus spp. on the phytochemical composition of juices from two varieties of Citrus sinensis L. Osbeck: ‘Tarocco’ and ‘Washington navel’. LWT Food Sci. Technol. 2020, 125, 109205. [Google Scholar] [CrossRef]
- Filannino, P.; Di Cagno, R.; Gobbetti, M. Metabolic and functional paths of lactic acid bacteria in plant foods: Get out of the labyrinth. Curr. Opin. Biotechnol. 2018, 49, 64–72. [Google Scholar] [CrossRef]
- Monsen, E.R. Dietary reference intakes for the antioxidant nutrients: Vitamin C, vitamin E, selenium, and carotenoids. J. Am. Diet. Assoc. 2000, 100, 637–640. [Google Scholar] [CrossRef]
- Huang, X.-M.; Wang, H.-C.; Yuan, W.-Q.; Lu, J.-M.; Yin, J.-H.; Luo, S.; Huang, H.-B. A study of rapid senescence of detached litchi: Roles of water loss and calcium. Postharvest Biol. Technol. 2005, 36, 177–189. [Google Scholar] [CrossRef]
- Piljac-Žegarac, J.; Šamec, D. Antioxidant stability of small fruits in postharvest storage at room and refrigerator temperatures. Food Res. Int. 2011, 44, 345–350. [Google Scholar] [CrossRef]
- Landete, J.M.; Curiel, J.A.; Rodríguez, H.; de las Rivas, B.; Muñoz, R. Aryl glycosidases from Lactobacillus plantarum increase antioxidant activity of phenolic compounds. J. Funct. Foods 2014, 7, 322–329. [Google Scholar] [CrossRef] [Green Version]
- Venil, C.K.; Dufossé, L.; Renuka Devi, P. Bacterial pigments: Sustainable compounds with market potential for pharma and food industry. Front. Sustain. Food Syst. 2020, 4, 100. [Google Scholar] [CrossRef]
- Mapelli-Brahm, P.; Barba, F.J.; Remize, F.; Garcia, C.; Fessard, A.; Khaneghah, A.M.; Sant’Ana, A.S.; Lorenzo, J.M.; Montesano, D.; Meléndez-Martínez, A.J. The impact of fermentation processes on the production, retention and bioavailability of carotenoids: An overview. Trends Food Sci. Technol. 2020, 99, 389–401. [Google Scholar] [CrossRef]
- Antognoni, F.; Mandrioli, R.; Potente, G.; Saa, D.L.T.; Gianotti, A. Changes in carotenoids, phenolic acids and antioxidant capacity in bread wheat doughs fermented with different lactic acid bacteria strains. Food Chem. 2019, 292, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Schlesier, K.; Harwat, M.; Böhm, V.; Bitsch, R. Assessment of antioxidant activity by using different in vitro methods. Free Radic. Res. 2002, 36, 177–187. [Google Scholar] [CrossRef]
- Scalzo, R.L. Organic acids influence on DPPH scavenging by ascorbic acid. Food Chem. 2008, 107, 40–43. [Google Scholar] [CrossRef]
- Karadag, A.; Ozcelik, B.; Saner, S. Review of methods to determine antioxidant capacities. Food Anal. Methods 2009, 2, 41–60. [Google Scholar] [CrossRef]
- Di Cagno, R.; Surico, R.F.; Paradiso, A.; De Angelis, M.; Salmon, J.-C.; Buchin, S.; De Gara, L.; Gobbetti, M. Effect of autochthonous lactic acid bacteria starters on health-promoting and sensory properties of tomato juices. Int. J. Food Microbiol. 2009, 128, 473–483. [Google Scholar] [CrossRef]
- Smit, G.; Smit, B.A.; Engels, W.J. Flavour formation by lactic acid bacteria and biochemical flavour profiling of cheese products. FEMS Microbiol. Rev. 2005, 29, 591–610. [Google Scholar] [CrossRef]
- Tiwari, S.; Kate, A.; Mohapatra, D.; Tripathi, M.K.; Ray, H.; Akuli, A.; Ghosh, A.; Modhera, B. Volatile organic compounds (VOCs): Biomarkers for quality management of horticultural commodities during storage through e-sensing. Trends Food Sci. Technol. 2020, 106, 417–433. [Google Scholar] [CrossRef]
- Di Cagno, R.; Filannino, P.; Gobbetti, M. Lactic acid fermentation drives the optimal volatile flavor-aroma profile of pomegranate juice. Int. J. Food Microbiol. 2017, 248, 56–62. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mango Cultivar Juice | TSS (°Brix) | pH | TA (Equivalent Lactic Acid g/100 g) |
|---|---|---|---|
| ‘peach’ mango juice | |||
| U 0 h (Raw) | 3.42 ± 0.27 a | 4.84 ± 0.03 a | 20.00 ± 0.02 e |
| U 24 h (un-inoculated) | 3.42 ± 0.29 a | 4.85 ± 0.05 a | 21.00 ± 0.01 e |
| L75 24 h | 1.72 ± 0.08 d | 3.88 ± 0.01 b,c | 30.00 ± 0.01 b,c |
| L56 24 h | 2.14 ± 0.17 b | 3.96 ± 0.01 b | 27.00 ± 0.03 c,d |
| L56 + 75 24 h | 2.03 ± 0.04 c | 3.89 ± 0.01 b,c | 29.00 ± 0.01 c |
| ‘sabre’ mango juice | |||
| U 0 h (Raw) | 2.40 ± 0.07 b | 4.28 ± 0.36 a | 23.00 ± 0.03 d,e |
| U 24 h (un-inoculated) | 2.30 ± 0.07 b | 4.18 ± 0.11 a,b | 23.00 ± 0.02 d,e |
| L75 24 h | 1.52 ± 0.08 d | 3.66 ± 0.03 d | 36.00 ± 0.04 a |
| L56 24 h | 2.24 ± 0.11 b | 3.88 ± 0.01 c | 35.00 ± 0.04 a,b |
| L56 + 75 24 h | 2.26 ± 0.11 b | 3.80 ± 0.05 c | 36.00 ± 0.03 a |
| ‘Tommy Atkins’ mango juice | |||
| U 0 h (Raw) | 1.76 ± 0.05 d | 4.12 ± 0.01 a | 23.00 ± 0.05 d,e |
| U 24 h (un-inoculated) | 1.76 ± 0.05 d | 4.12 ± 0.01 a | 24.00 ± 0.02 d,e |
| L75 24 h | 0.62 ± 0.04 e | 4.01 ± 0.01 c | 28.00 ± 0.02 c |
| L56 24 h | 0.72 ± 0.08 e | 4.07 ± 0.04 b | 26.00 ± 0.02 c,d |
| L56 + 75 24 h | 0.70 ± 0.07 e | 4.03 ± 0.01 c | 27.00 ± 0.04 c,d |
| LSD | 2.44 * | 0.01 ** | 0.04 *** |
| Mango Cultivar Juice | L* | b* | c* | Δε |
|---|---|---|---|---|
| ‘Peach’ mango juice | ||||
| U 0 h (Raw) | 33.62 ± 0.36 a | 18.32 ± 0.65 a | 18.40 ± 0.65 a | |
| U 24 h (un-inoculated) | 31.38 ± 0.62 b | 16.97 ± 0.09 b | 17.01 ± 0.11 b | 6.09 ± 0.05 a |
| L75 24 h | 33.08 ± 0.40 a | 18.42 ± 0.89 a | 18.63 ± 0.85 a | 1.64 ± 0.05 k |
| L56 24 h | 33.73 ± 0.34 a | 18.46 ± 0.71 a | 18.76 ± 0.16 a | 1.41 ± 0.05 l |
| L56 + 75 24 h | 29.52 ± 0.19 b | 15.51 ± 0.45 c | 15.63 ± 0.48 c | 3.49 ± 0.05 f |
| ‘Sabre’ mango juice | ||||
| U 0 h (Raw) | 33.04 ± 0.60 a | 18.25 ± 1.12 a | 18.41 ± 1.14 a | |
| U 24 h (un-inoculated) | 30.46 ± 0.33 b | 15.36 ± 0.30 d | 15.54 ± 0.31 c | 5.64 ± 0.05 b |
| L75 24 h | 30.18 ± 0.48 b | 14.15 ± 0.44 e | 14.32 ± 0.47 d | 4.00 ± 0.05 d |
| L56 24 h | 30.57 ± 0.19 b | 15.98 ± 0.61 b,c | 16.08 ± 0.55 b,c | 3.41 ± 0.05 h |
| L56 + 75 24 h | 30.29 ± 0.27 b | 13.52 ± 0.88 e | 13.77 ± 0.93 d | 5.16 ± 0.05 c |
| ‘Tommy Atkins’ mango juice | ||||
| U 0 h (Raw) | 27.03 ± 0.66 c | 11.12 ± 0.14 f | 9.90 ± 0.25 e | |
| U 24 h (un-inoculated) | 25.02 ± 0.13 d | 9.16 ± 0.45 g | 9.90 ± 0.25 e | 3.54 ± 0.05 e |
| L75 24 h | 25.00 ± 0.21 d | 8.78 ± 0.10 g | 9.42 ± 0.12 e | 2.65 ± 0.05 i |
| L56 24 h | 25.87 ± 0.10 d | 9.55 ± 0.10 g | 10.44 ± 0.09 e | 2.25 ± 0.05 j |
| L56 + 75 24 h | 25.82 ± 0.07 d | 8.93 ± 0.25 g | 9.58 ± 0.27 e | 3.43± 0.05 g |
| LSD | 0.19 ** | 4.65 *** | 10.44 *** | 0.18 *** |
| Juice of Different Mango Cultivar | Ascorbic Acid (mg/100 g) | TPC (mg/100 mL) | Beta-Carotene (µg/mL) | FRAP (µM TEAC/mL) | DPPH IC50 (mg/mL) | ABTS IC50 (mg/mL) |
|---|---|---|---|---|---|---|
| ‘Peach’ mango juice | ||||||
| U 0 h (control) | 10.42 ± 0.06 d | 1171.41 ± 0.01 f | 3.78 ± 0.22 h | 354.19 ± 0.06 g | 16.33 ± 0.01 n | 10.67 ± 0.11 o |
| U 24 h (un-inoculated) | 5.19 ± 0.06 k | 1051.45 ± 0.01 k | 10.85 ± 0.83 d | 254.94 ± 0.01 n | 163.52 ± 0.02 a | 28.29 ± 0.06 e |
| L75 24 h | 4.49 ± 0.06 l | 1393.07 ± 0.01 b | 17.02 ± 1.62 b | 550.39 ± 0.01 b | 95.92 ± 0.01 e | 14.51 ± 0.43 m |
| L56 24 h | 4.13 ± 0.01 m | 1170.07 ± 0.01 h | 19.89 ± 1.16 a | 330.39 ± 0.01 h | 109.51 ± 0.01 c | 27.10 ± 1.01 f |
| L56 + 75 24 h | 3.87 ± 0.01 n | 1364.57 ± 0.01 c | 16.33 ± 1.95 c | 461.33 ± 0.58 d | 104.15 ±0.01 d | 22.16 ± 0.01 l |
| ‘Sabre’ mango juice | ||||||
| U 0 h (control) | 15.51 ± 0.01 a | 1167.03 ± 0.01 i | 0.64 ± 0.52 j | 356.91 ± 0.01 f | 22.17 ± 0.01 m | 24.97 ± 0.01 j |
| U 24 h (un-inoculated) | 10.71 ± 0.01 c | 1007.08 ± 0.01 m | 4.02 ± 0.19 g | 295.06 ± 0.01 e | 56.04 ± 0.01 i | 43.27 ± 2.17 a |
| L75 24 h | 11.23 ± 0.01 b | 1415.71 ± 0.01 a | 4.32 ± 0.81 f | 558.28 ± 0.01 a | 44.77 ± 0.01 l | 25.27 ± 1.11 i |
| L56 24 h | 9.03 ± 0.01 g | 1174.64 ± 0.01 e | 8.03 ± 1.25 e | 390.37 ± 0.06 l | 54.96 ± 0.01 j | 37.38 ± 6.06 d |
| L56 + 75 24 h | 9.29 ± 0.01 f | 1211.33 ± 0.01 d | 0.97 ± 0.42 i | 547.51 ±0.01 c | 52.52 ± 0.01 k | 27.45 ± 4.33 g |
| ‘Tommy Atkins’ mango juice | ||||||
| U 0 h (control) | 9.68 ± 0.01 e | 923.06 ± 0.01 n | 0.02 ± 0.00 o | 297.74 ± 0.01 k | 58.52 ± 0.01 h | 13.79 ± 0.91 n |
| U 24 h (un-inoculated) | 8.52 ± 0.01 h | 1089.72 ± 0.01 j | 0.08 ± 0.01 n | 229.74 ± 0.02 o | 112.53 ± 0.01 b | 40.06 ± 3.51 b |
| L75 24 h | 7.87 ± 0.01 i | 1170.84 ± 0.01 g | 0.12 ±0.00 m | 322.07 ± 0.01 i | 58.28 ± 0.01 h | 23.04 ± 1.24 k |
| L56 24 h | 3.61 ± 0.01 o | 1046.38 ± 0.01 l | 0.15 ± 0.02 k | 284.19 ± 0.01 m | 62.87 ± 0.01 g | 39.86 ± 2.11 c |
| L56 + 75 24 h | 7.23 ± 0.01 j | 823.31 ± 0.01 l | 0.13 ±0.02 l | 298.07 ± 0.01 j | 63.98 ± 0.01 f | 26.86 ± 0.77 i |
| LSD | 3.90 *** | 8.019 *** | 0.56 *** | 1.34 *** | 1.27 *** | 0.14 *** |
| ‘Peach’ | ‘Sabre’ | ‘Tommy Atkins’ | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Volatile Compounds | U 0 h | U 24 h | L75 24 h | L56 24 h | L56 + 75 24 h | U 0 h | U 24 h | L75 24 h | L56 24 h | L56 + 75 24 h | U 0 h | U 24 h | L75 24 h | L56 24 h | L56 + 75 24 h |
| ALKANES | |||||||||||||||
| octane-2-methyl | 9.5 b | 0.7 d | 9.8 b | 2.0 d | 7.1 b,c | 9.5 b | 3.7 c,d | 18.3 a | 7.7 b | 1.2 d | ND | 0.7 d | 9.5 b | ND | ND |
| decane-3,7-dimethyl | 9.1 d,e | 7.3 d,e,f | 20.3 b | 11.4 c,d | 8.3 d,e | 8.7 d,e | 5.9 e,f,g | 26.2 a | 8.6 d,e | 13.9 c | 2.7 f,g | 1.3 g | 15.6 c | 6.3 e,f | 1.6 g |
| Nonadecane | 2.2 e,f,g | 1.2 f,g | 17.9 a | 12.0 b,c | 6.9 d,e | 5.6 d,e,f | 2.5 e,f,g | 18.4 a | 14.4 a,b | 2.7 e,f,g | 2.7 e,f,g | ND | 15.7 a,b | ND | 7.7 c,d |
| hexadecane | 3.1 c | 1.6 c | 8.9 a,b | 7.7 b | 8.2 a,b | 3.0 c | 3.0 c | 10.8 a | 2.9 c | ND | 1.3 c | 1.1 c | 1.6 c | 1.4 c | 2.3 c |
| pentadecane-8-hexyl | 7.8 c,d | 0.8 d | 22.1 b | 8.2 c,d | 10.9 c | 8.6 c,d | ND | 60.9 a | 4.3 e | 1.0 e | 4.5 d,e | ND | 8.9 c,d | 1.3 e | ND |
| heptane-2,6-dimethyl (piney aroma) | 0.3 e | 0.7 e | 3.4 b | 0.9 e | 0.7 e | 2.3 c | 1.5 d | 4.4 a | 2.6 c | 0.55 e | 0.9 e | 0.5 e | 0.4 e | 0.5 e | 0.3 e |
| KETONES | |||||||||||||||
| 2-pentanone (fruity) | ND | ND | ND | ND | ND | ND | ND | 6.3 a | ND | ND | ND | ND | ND | ND | ND |
| 2-heptanone (fruity/floral) | ND | ND | ND | ND | ND | ND | ND | 7.6 a | ND | ND | ND | ND | ND | ND | ND |
| ESTERS | |||||||||||||||
| butyl isobutyrate (fruity type) | ND | ND | 10.3 b | ND | ND | 1.8 d | ND | 15.5 a | ND | 0.1 e,f | ND | ND | 1.8 c,d | 1.6 d,e | ND |
| butyl acetate (banana or apple) | ND | ND | ND | ND | ND | ND | 1.2 b | 22.6 a | ND | ND | ND | ND | ND | ND | ND |
| amyl acetate (fresh-fruity, reminiscent of pear, banana and apple) | ND | ND | ND | ND | ND | ND | ND | 0.3 a | ND | ND | ND | ND | ND | ND | ND |
| ethyl hexanoate (fruity type) | ND | ND | 12.4 a | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| methyl acetate (fruity) | ND | ND | 4.7 a | ND | 0.6 b | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| ethyl acetate (fruity) | ND | ND | 9.5 a | 5.7 b | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| 2,6-di-t-butyl-methylphenol acetate | 56.5 b | 14.0 d | 83.6 a | 57.4 b | 30.3 c,d | 18.6 d | 20.1 d | 84.9 a | 48.1 b,c | 22.5 d | 19.1 d | 18.9 d | 57.4 b | 34.2 c,d | 29.0 c,d |
| ethyl butyrate (fruity) | 24.0 c,d | 16.4 f | 49.0 a | 43.4 a | 46.7 a | 31.8 b | 18.0 e,f | 49.0 a | 44.7 a | 47.7 a | ND | 21.7 d,e,f | 33.4 b | 22.0 d,e | 28.7 b,c |
| ethyl octanoate | 3.28 h | 2.6 h | 21.3 d | 13.6 f | 13.3 f | 12.5 f | 5.5 g | 30.6 a | 26.6 b | 24.3 c | 2.6 h | ND | 17.9 e | 2.7 h | 13.6 f |
| isobutyl acetate (waxy type) | 5.5 d | ND | 4.8 d,e | 2.9 e | 3.3 e | 11.8 c | ND | 16.9 a | 14.1 b | 14.1 b | ND | ND | 3.9 d,e | 2.9 e | 4.0 d,e |
| ethyl decanoate (waxy type) | 2.9 c | ND | 5.2 b | ND | ND | ND | ND | 8.9 a | 5.2 b | 8.2 a | ND | ND | 4.3 b | ND | ND |
| methyl benzoate (floral) | ND | ND | ND | ND | ND | ND | ND | 3.8 a | ND | ND | ND | ND | ND | ND | ND |
| geranyl isovalerate (floral type, green) | ND | ND | ND | ND | ND | 0.9 b | ND | 1.3 a | 1.0 a,b | 0.9 b | ND | ND | ND | ND | ND |
| linalyl butyrate (floral type) | ND | ND | 1.5 c | ND | 0.9 d | ND | ND | 2.3 a | ND | 1.9 b | ND | ND | 0.9 d | ND | 0.7 d |
| ALDEHYDES | |||||||||||||||
| nonanal (rose-orange) | ND | ND | ND | ND | ND | ND | ND | 2.67 a | ND | ND | ND | ND | ND | ND | ND |
| e-2 hexenal (green) | 2.9 e,f | 0.9 f,g | 15.6 a,b | 1.2 f,g | 5.9 c,d | 0.9 f,g | 1.1 f,g | 17.6 a | 1.2 f,g | 7.2 c | 1.2 f,g | ND | 14.2 b | 4.6 d,e | 3.6 e |
| pentanal (fruity) | ND | ND | 25.9 a | ND | ND | ND | ND | 25.9 a | 0.04 d | 20.7 b | ND | ND | 0.04 d | 3.9 c | 0.03 d |
| decanal (green) | ND | ND | 1.7 b | ND | ND | ND | ND | 2.7 a | ND | ND | ND | ND | ND | ND | ND |
| ALCOHOL | |||||||||||||||
| e-3 hexanol (green) | 27.3 c,d | 13.9 e | 33.9 a,b | 27.3 c,d | 30.3 b,c | 27.3 c,d | 14.6 e | 37.3 a | 29.6 b,c | 37.3 a | 23.3 d | 11.9 e | 30.6 b,c | 27.9 c,d | 27.6 c,d |
| penten-3-ol (fruity, and green) | 13.3 c,d | 13.4 c,d | 23.3 b | 14.1 c,d | 18.0 b,c | 20.4 b | 17.4 b,c | 36.7 a | 22.7 b | 22.7 b | 20.5 b | 8.7 d | 21.4 b | 18.7 b,c | 20.3 b |
| 1-dodecanol (floral, soapy, waxy) | ND | ND | ND | ND | ND | 2.40 b | ND | 5.8 a | 2.8 b | 5.7 a | ND | ND | ND | ND | ND |
| TERPENE HYDROCARBONS | |||||||||||||||
| β-myrcene (earthy, fruity, and clove-like) | 4.1 d,e | 2.9 e | 9.1 c | 5.4 d,e | 5.4 d,e | 3.4 d,e | 1.9 e | 25.4 a | 5.4 d,e | 16.0 b | 5.4 e | 3.0 e | 7.0 c,d | 5.4 d,e | 5.4 d,e |
| TOTAL VOLATILE COMPOUNDS | 171.7 | 76.3 | 388.9 | 213.1 | 196.7 | 169.2 | 96.3 | 543.0 | 541.9 | 248.6 | 84.1 | 67.8 | 244.4 | 133.4 | 144.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cele, N.P.; Akinola, S.A.; Manhivi, V.E.; Shoko, T.; Remize, F.; Sivakumar, D. Influence of Lactic Acid Bacterium Strains on Changes in Quality, Functional Compounds and Volatile Compounds of Mango Juice from Different Cultivars during Fermentation. Foods 2022, 11, 682. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11050682
Cele NP, Akinola SA, Manhivi VE, Shoko T, Remize F, Sivakumar D. Influence of Lactic Acid Bacterium Strains on Changes in Quality, Functional Compounds and Volatile Compounds of Mango Juice from Different Cultivars during Fermentation. Foods. 2022; 11(5):682. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11050682
Chicago/Turabian StyleCele, Nobahle P., Stephen A. Akinola, Vimbainashe E. Manhivi, Tinotenda Shoko, Fabienne Remize, and Dharini Sivakumar. 2022. "Influence of Lactic Acid Bacterium Strains on Changes in Quality, Functional Compounds and Volatile Compounds of Mango Juice from Different Cultivars during Fermentation" Foods 11, no. 5: 682. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11050682