
Efficacy of the Yeast Wickerhamomyces anomalus in Biocontrol of Gray Mold Decay of Tomatoes and Study of the Mechanisms Involved



Abstract
:1. Introduction
2. Materials and Methods
2.1. Fruits
2.2. Antagonist
2.3. Fungal Pathogen
2.4. Inhibitory Activity of Different Yeasts against Tomato Gray Mold Caused by B. cinerea
2.5. Efficacy of W. anomalus at Different Concentrations against Tomato Gray Mold Caused by B. cinerea
2.6. Colonization Trends of W. anomalus in Tomato Tissue and Epidermis at 20 °C and 4 °C
2.7. Efficacy of W. anomalus on Enzyme Activities and Content of Total Phenols and Flavonoid Compounds in Tomatoes
2.8. Statistical Analysis
3. Results
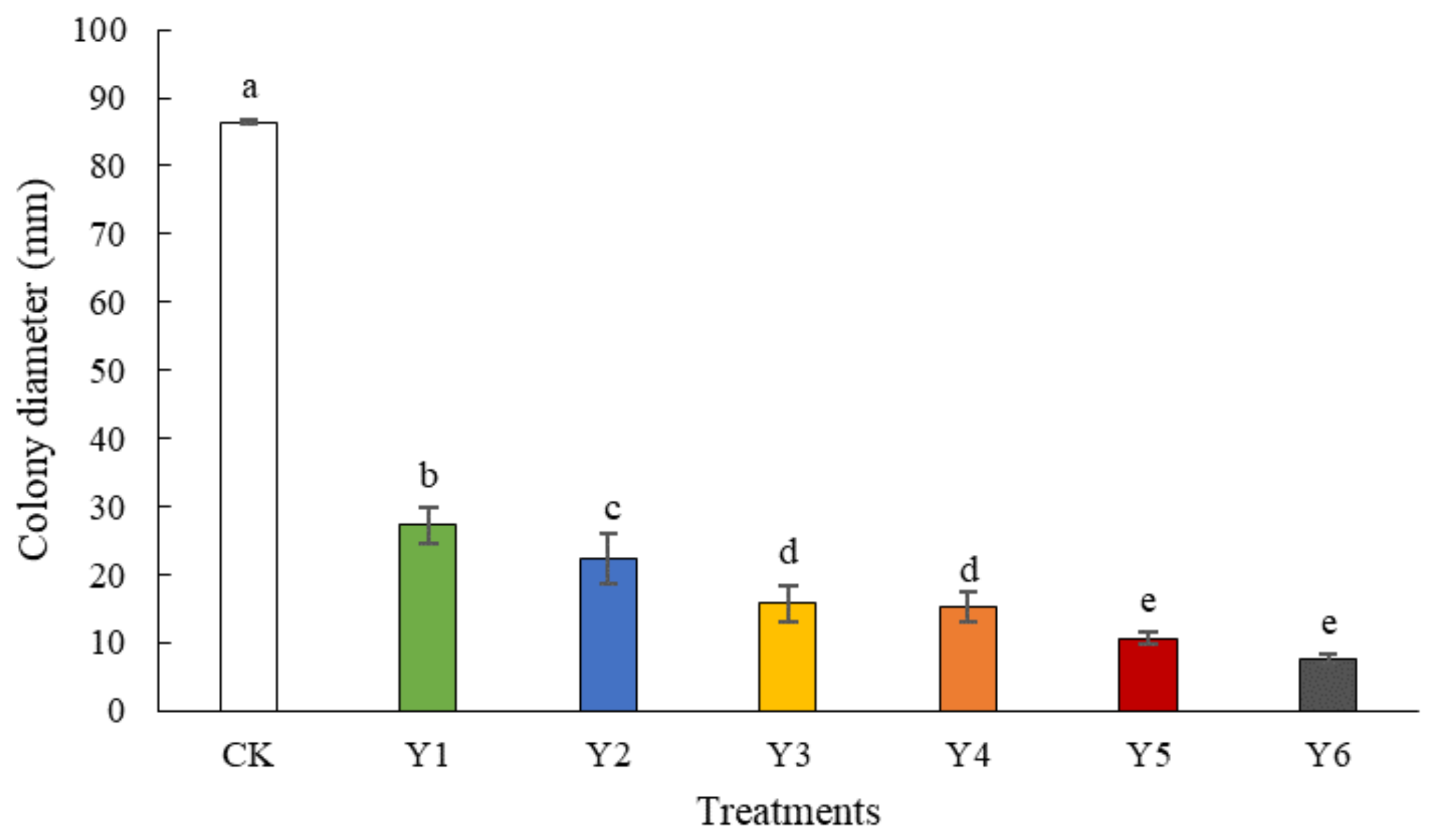
3.1. Inhibitory Effect of Different Yeasts against Tomato Gray Mold Caused by B. cinerea
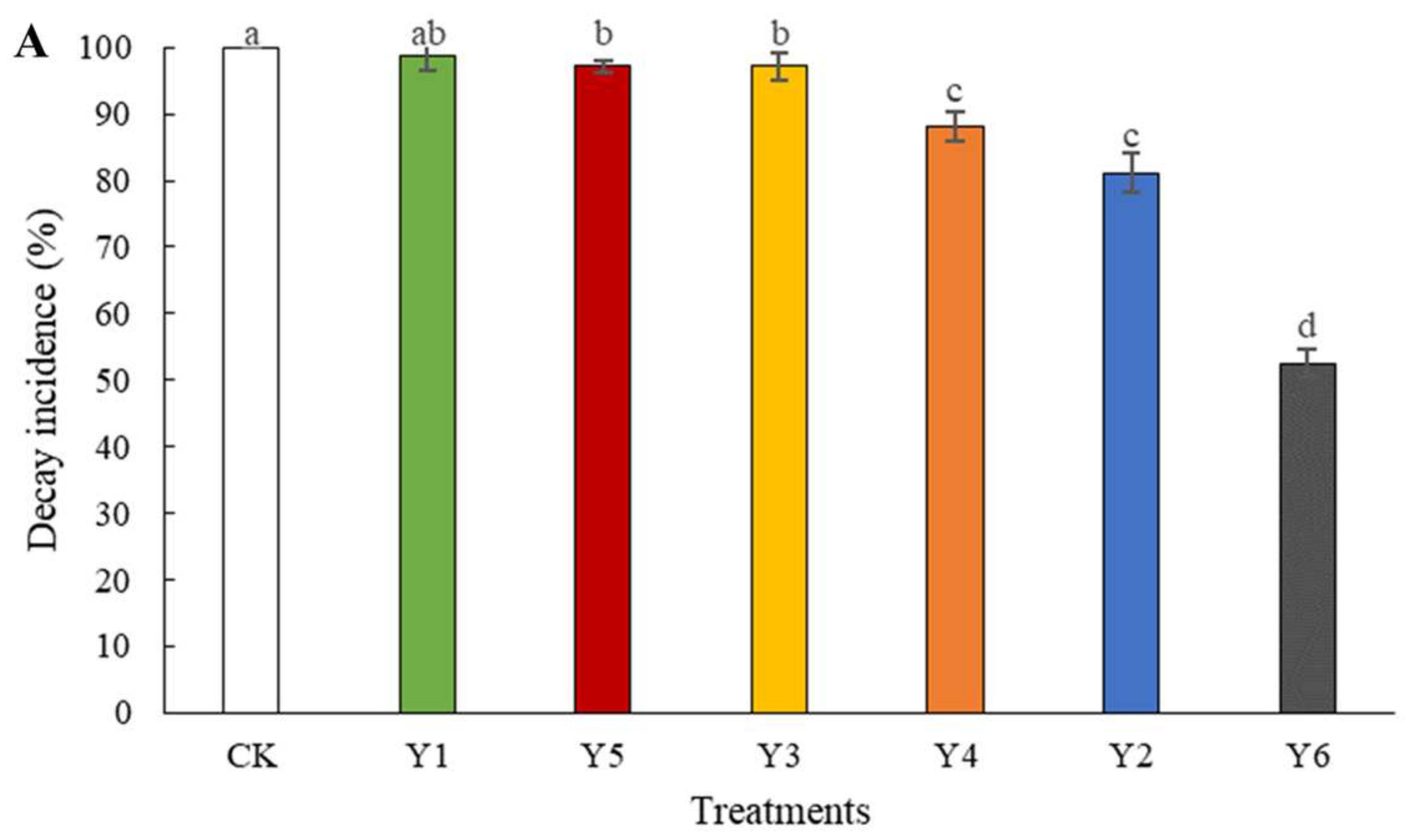
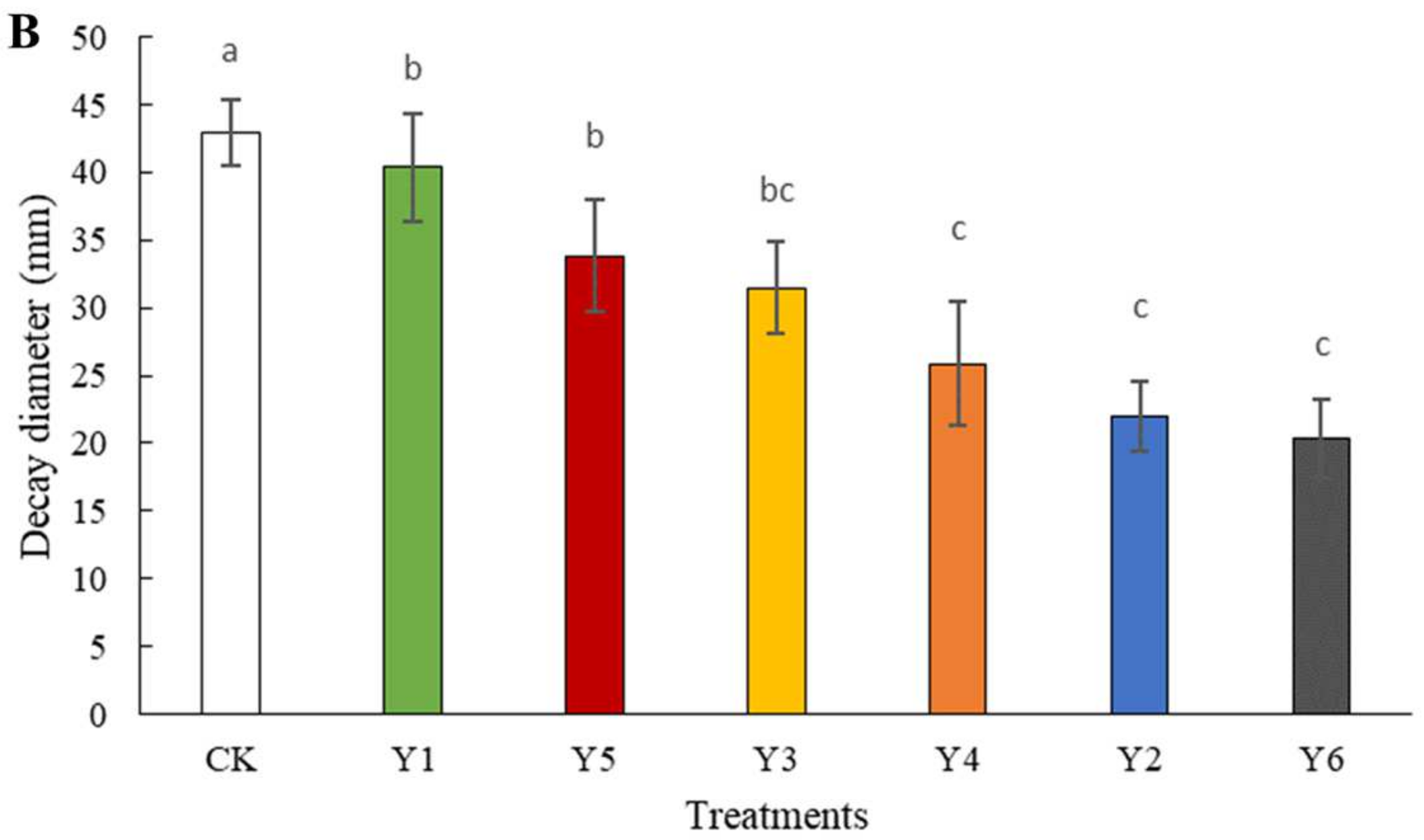
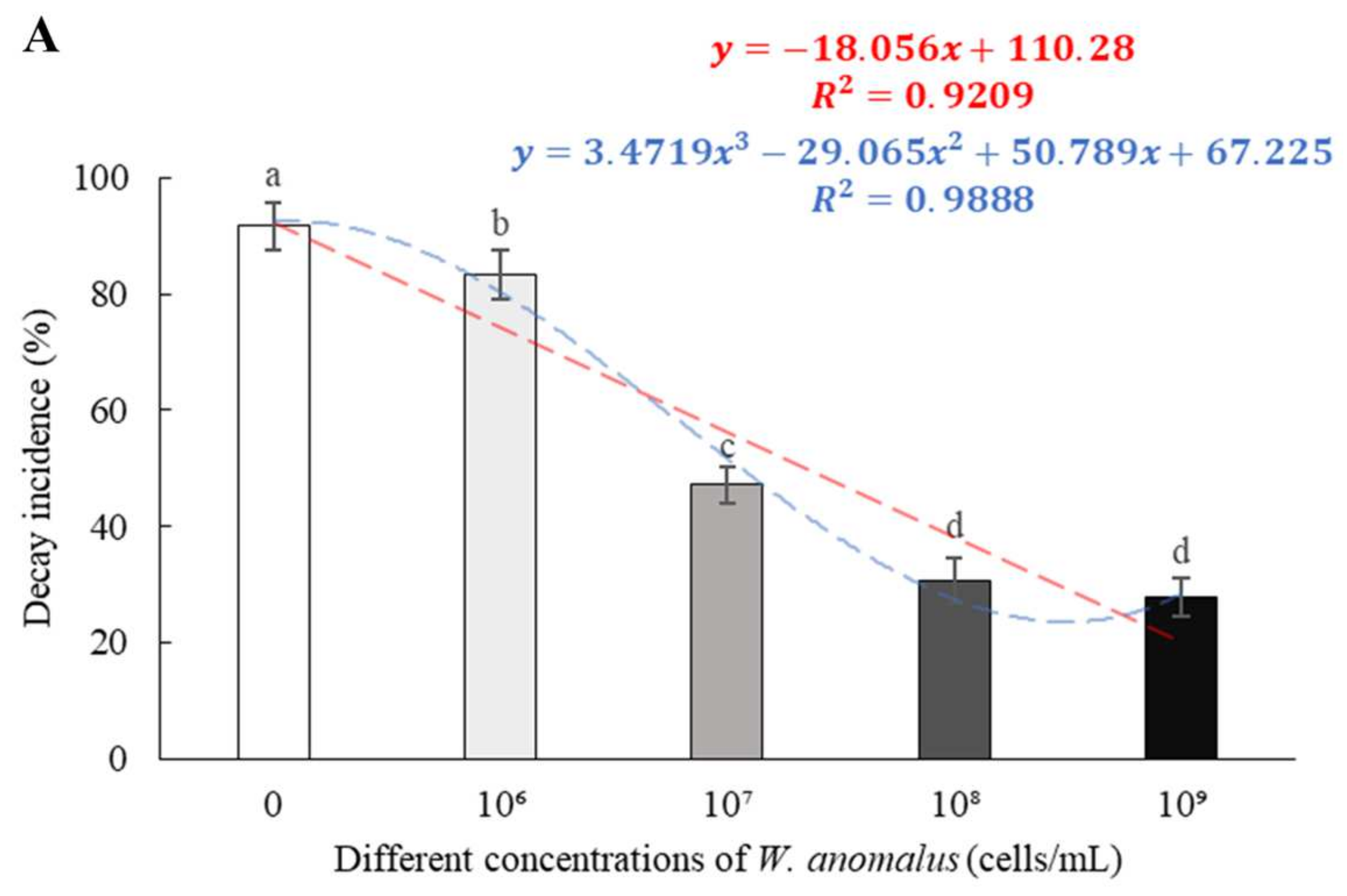
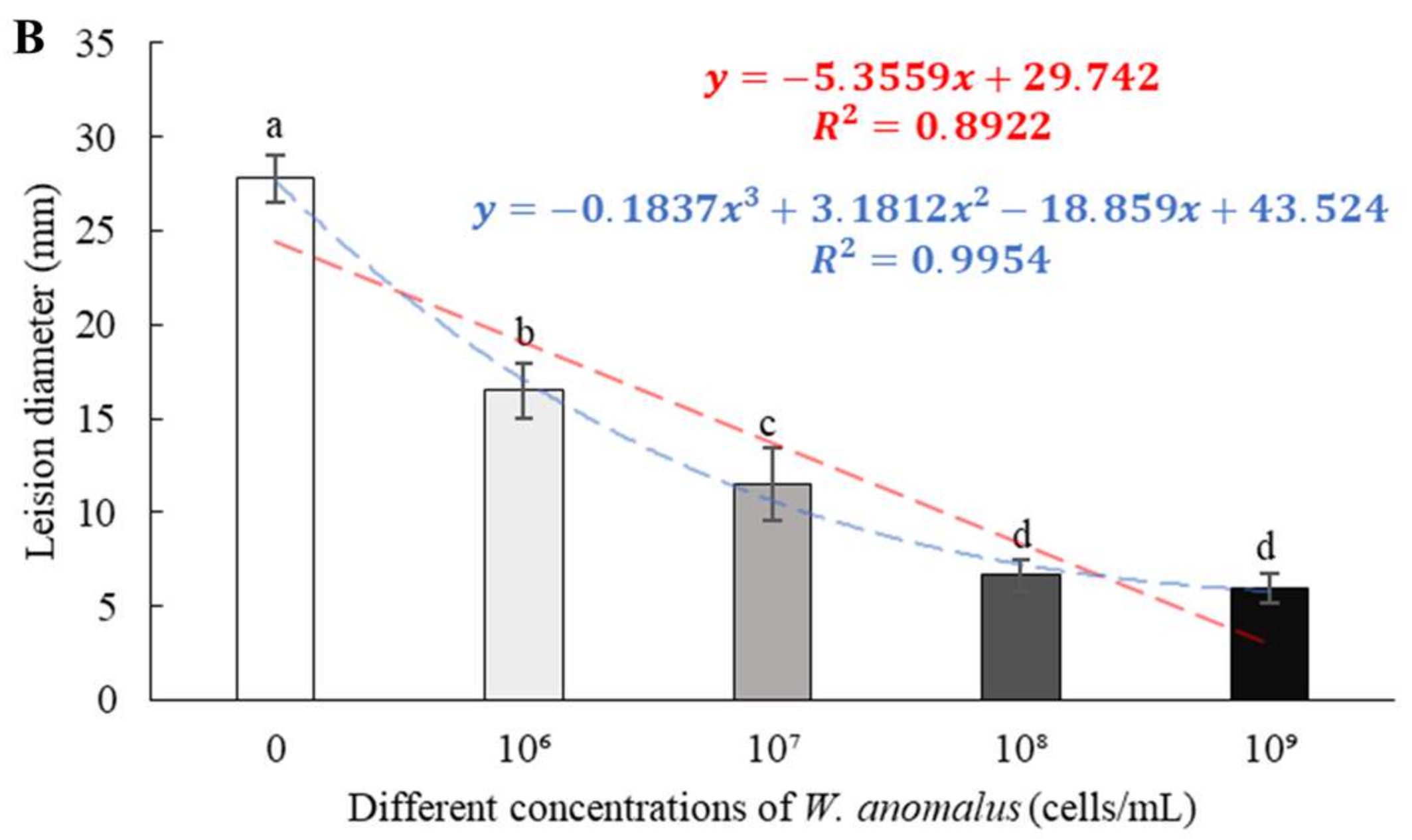
3.2. Efficacy of W. anomalus at Different Concentrations against the Tomato Gray Mold Caused by B. cinerea
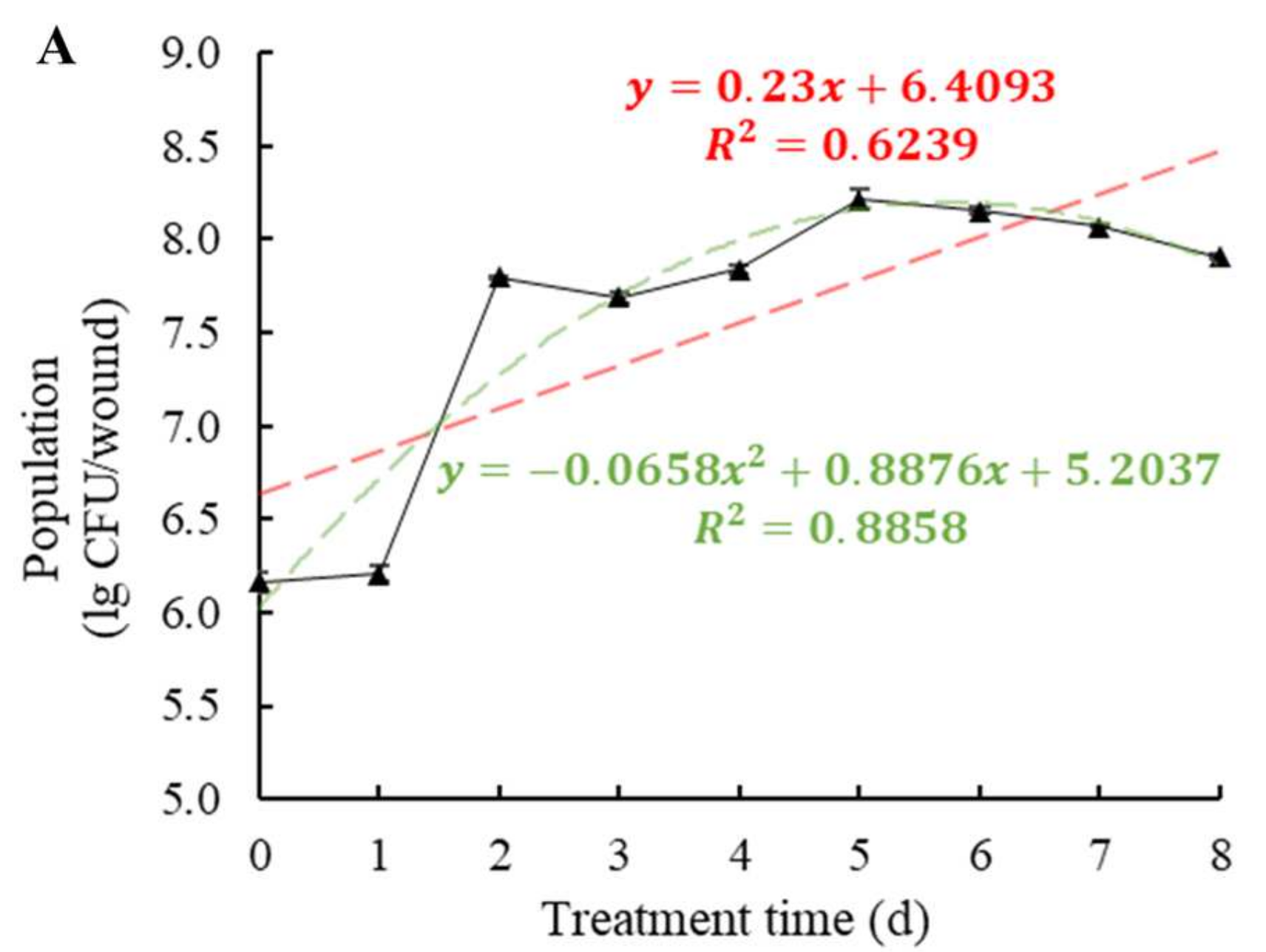
3.3. Colonization Trends of W. anomalus in Tomato Tissue and Epidermis at 20 °C and 4 °C
3.4. Effects of W. anomalus on Enzyme Activities and Total Phenolic and Flavonoid Compounds Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Feng, W.; Zheng, X. Essential oils to control Alternaria alternata in vitro and in vivo. Food Control. 2007, 18, 1126–1130. [Google Scholar] [CrossRef]
- Liu, S.; Che, Z.; Chen, G. Multiple-fungicide resistance to carbendazim, diethofencarb, procymidone, and pyrimethanil in field isolates of Botrytis cinerea from tomato in Henan Province, China. Crop. Prot. 2016, 84, 56–61. [Google Scholar] [CrossRef]
- Dik, A.J.; Elad, Y. Comparison of antagonists of Botrytis cinerea in greenhouse-grown cucumber and tomato under different climatic conditions. Eur. J. Plant. Pathol. 1999, 105, 123–137. [Google Scholar] [CrossRef]
- Małolepsza, U. Induction of disease resistance by acibenzolar-S-methyl and o-hydroxyethylorutin against Botrytis cinerea in tomato plants. Crop. Prot. 2006, 25, 956–962. [Google Scholar] [CrossRef]
- Dal Bello, G.; Mónaco, C.; Rollan, M.C.; Lampugnani, G.; Arteta, N.; Abramoff, C.; Ronco, L.; Stocco, M. Biocontrol of postharvest grey mould on tomato by yeasts. J. Phytopathol. 2008, 156, 257–263. [Google Scholar] [CrossRef]
- Eckert, J.W.; Ogawa, J.M. The chemical control of postharvest diseases: Subtropical and tropical fruits. Annu. Rev. Phytopathol. 1985, 23, 421–454. [Google Scholar] [CrossRef]
- Leroux, P. Chemical control of Botrytis and its resistance to chemical fungicides. In Botrytis: Biology, Pathology and Control; Elad, Y., Williamson, B., Tudzynski, P., Delen, N., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 195–222. [Google Scholar]
- Sun, H.-Y.; Wang, H.-C.; Chen, Y.; Li, H.-X.; Chen, C.-J.; Zhou, M.-G. Multiple resistance of Botrytis cinerea from vegetable crops to Carbendazim, Diethofencarb, Procymidone, and Pyrimethanil in China. Plant. Dis. 2010, 94, 551–556. [Google Scholar] [CrossRef] [Green Version]
- De Curtis, F.; Lima, G.; Vitullo, D.; De Cicco, V. Biocontrol of Rhizoctonia solani and Sclerotium rolfsii on tomato by delivering antagonistic bacteria through a drip irrigation system. Crop. Prot. 2010, 29, 663–670. [Google Scholar] [CrossRef]
- Yang, R.; Fan, X.; Cai, X.; Hu, F. The inhibitory mechanisms by mixtures of two endophytic bacterial strains isolated from Ginkgo biloba against pepper phytophthora blight. Biol. Control. 2015, 85, 59–67. [Google Scholar] [CrossRef]
- De Simone, N.; Pace, B.; Grieco, F.; Chimienti, M.; Tyibilika, V.; Santoro, V.; Capozzi, V.; Colelli, G.; Spano, G.; Russo, P. Botrytis cinerea and table grapes: A review of the main physical, chemical, and bio-based control treatments in post-harvest. Foods 2020, 9, 1138. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, T.; Xia, J.; Yu, D.; Wang, J.; Zheng, X. Biocontrol of postharvest gray mold of cherry tomatoes with the marine yeast Rhodosporidium paludigenum. Biol. Control. 2010, 53, 178–182. [Google Scholar] [CrossRef]
- Guo, H.; Qin, X.; Wu, Y.; Yu, W.; Liu, J.; Xi, Y.; Dou, G.; Wang, L.; Xiao, H. Biocontrol of gray mold of cherry tomatoes with the volatile organic monomer from Hanseniaspora uvarum, trans-Cinnamaldehyde. Food Bioprocess. Technol. 2019, 12, 1809–1820. [Google Scholar] [CrossRef]
- Fernandez-San Millan, A.; Larraya, L.; Farran, I.; Ancin, M.; Veramendi, J. Successful biocontrol of major postharvest and soil-borne plant pathogenic fungi by antagonistic yeasts. Biol. Control. 2021, 160, 104683. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhao, L.; Li, Z.; Li, C.; Li, B.; Gu, X.; Zhang, X.; Zhang, H. Screening and identification of an antagonistic yeast controlling postharvest blue mold decay of pears and the possible mechanisms involved. Biol. Control. 2019, 133, 26–33. [Google Scholar] [CrossRef]
- He, F.; Zhao, L.; Zheng, X.; Abdelhai, M.H.; Boateng, N.S.; Zhang, X.; Zhang, H. Investigating the effect of methyl jasmonate on the biocontrol activity of Meyerozyma guilliermondii against blue mold decay of apples and the possible mechanisms involved. Physiol. Mol. Plant. Pathol. 2020, 109, 101454. [Google Scholar] [CrossRef]
- Apaliya, M.T.; Zhang, H.; Yang, Q.; Zheng, X.; Zhao, L.; Kwaw, E.; Mahunu, G.K. Hanseniaspora uvarum enhanced with trehalose induced defense-related enzyme activities and relative genes expression levels against Aspergillus tubingensis in table grapes. Postharvest Biol. Technol. 2017, 132, 162–170. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Xu, W.; Zheng, X.; Zhang, X.; Abdelhai, M.H.; Zhao, L.; Li, H.; Diao, J.; Zhang, H. Exploring the effect of β-glucan on the biocontrol activity of Cryptococcus podzolicus against postharvest decay of apples and the possible mechanisms involved. Biol. Control. 2018, 121, 14–22. [Google Scholar] [CrossRef]
- Wang, M.; Zhao, L.; Zhang, X.; Dhanasekaran, S.; Abdelhai, M.H.; Yang, Q.; Jiang, Z.; Zhang, H. Study on biocontrol of postharvest decay of table grapes caused by Penicillium rubens and the possible resistance mechanisms by Yarrowia lipolytica. Biol. Control. 2019, 130, 110–117. [Google Scholar] [CrossRef]
- Li, J.; Yan, J.; Ritenour, M.A.; Wang, J.; Cao, J.; Jiang, W. Effects of 1-methylcyclopropene on the physiological response of Yali pears to bruise damage. Sci. Hortic. 2016, 200, 137–142. [Google Scholar] [CrossRef]
- Deng, L.; Zeng, K.; Zhou, Y.; Huang, Y. Effects of postharvest oligochitosan treatment on anthracnose disease in citrus (Citrus sinensis L. Osbeck) fruit. Eur. Food Res. Technol. 2015, 240, 795–804. [Google Scholar] [CrossRef]
- Zhang, H.; Apaliya, M.T.; Mahunu, G.K.; Chen, L.; Li, W. Control of ochratoxin A-producing fungi in grape berry by microbial antagonists: A review. Trends Food Sci. Technol. 2016, 51, 88–97. [Google Scholar] [CrossRef]
- Amina, K.; Essghaier, B.; Abdeljabar, H.; Sadfi Zouaoui, N. New epiphytic yeasts able to reduce grey mold disease on apples. Plant. Prot. Sci. 2018, 54, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Dunlap, C.A.; Schisler, D.A.; Bowman, M.J.; Rooney, A.P. Genomic analysis of Bacillus subtilis OH 131.1 and co-culturing with Cryptococcus flavescens for control of Fusarium head blight. Plant. Gene 2015, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Li, C.; Li, P.; Zhang, H.; Zhang, X.; Zheng, X.; Yang, Q.; Apaliya, M.T.; Boateng, N.A.S.; Sun, Y. The biocontrol effect of Sporidiobolus pararoseus Y16 against postharvest diseases in table grapes caused by Aspergillus niger and the possible mechanisms involved. Biol. Control. 2017, 113, 18–25. [Google Scholar] [CrossRef]
- Huang, R.; Che, H.J.; Zhang, J.; Yang, L.; Jiang, D.H.; Li, G.Q. Evaluation of Sporidiobolus pararoseus strain YCXT3 as biocontrol agent of Botrytis cinerea on post-harvest strawberry fruits. Biol. Control. 2012, 62, 53–63. [Google Scholar] [CrossRef]
- Oro, L.; Feliziani, E.; Ciani, M.; Romanazzi, G.; Comitini, F. Volatile organic compounds from Wickerhamomyces anomalus, Metschnikowia pulcherrima and Saccharomyces cerevisiae inhibit growth of decay causing fungi and control postharvest diseases of strawberries. Int. J. Food Microbiol. 2018, 265, 18–22. [Google Scholar] [CrossRef]
- Toffano, L.; Fialho, M.B.; Pascholati, S.F. Potential of fumigation of orange fruits with volatile organic compounds produced by Saccharomyces cerevisiae to control citrus black spot disease at postharvest. Biol. Control. 2017, 108, 77–82. [Google Scholar] [CrossRef]
- Cao, J.; Zhang, H.; Yang, Q.; Ren, R. Efficacy of Pichia caribbica in controlling blue mold rot and patulin degradation in apples. Int. J. Food Microbiol. 2013, 162, 167–173. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, G.; Li, P.; Yang, Q.; Chen, K.; Zhao, L.; Apaliya, M.T.; Gu, X.; Zhang, H. Mechanisms of glycine betaine enhancing oxidative stress tolerance and biocontrol efficacy of Pichia caribbica against blue mold on apples. Biol. Control. 2017, 108, 55–63. [Google Scholar] [CrossRef]
- Mahunu, G.K.; Zhang, H.; Yang, Q.; Zhang, X.; Li, D.; Zhou, Y. Improving the biocontrol efficacy of Pichia caribbica with phytic acid against postharvest blue mold and natural decay in apples. Biol. Control. 2016, 92, 172–180. [Google Scholar] [CrossRef]
- Czarnecka, M.; Żarowska, B.; Połomska, X.; Restuccia, C.; Cirvilleri, G. Role of biocontrol yeasts Debaryomyces hansenii and Wickerhamomyces anomalus in plants’ defence mechanisms against Monilinia fructicola in apple fruits. Food Microbiol. 2019, 83, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Sun, Y.; Yang, D.; Li, J.; Gu, X.; Zhang, X.; Zhang, H. Effects of Sporidiobolus pararoseus Y16 on postharvest blue mold decay and the defense response of apples. J. Food Qual. 2018, 2018, 6731762. [Google Scholar] [CrossRef] [Green Version]
- El-Ghaouth, A. Biologically-based alternatives to synthetic fungicides for the control of postharvest diseases. J. Ind. Microbiol. Biotechnol. 1997, 19, 160–162. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, Y.; Dhanasekaran, S.; Guo, Z.; Chen, S.; Zhang, X.; Zhang, H. Efficacy of Wickerhamomyces anomalus yeast in the biocontrol of blue mold decay in apples and investigation of the mechanisms involved. Biocontrol 2021, 66, 547–558. [Google Scholar] [CrossRef]
- Sharma, R.R.; Singh, D.; Singh, R. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: A review. Biol. Control. 2009, 50, 205–221. [Google Scholar] [CrossRef]
- Chen, L.; Tan, J.T.G.; Zhao, X.; Yang, D.; Yang, H. Energy regulated enzyme and non-enzyme-based antioxidant properties of harvested organic mung bean sprouts (Vigna radiata). LWT 2019, 107, 228–235. [Google Scholar] [CrossRef]
- Farkas, G.L.; Stahmann, M.A. On the nature of changes in peroxidase isoenzymes in bean leaves infected by southern bean mosaic virus. Phytopathology 1966, 56, 669–677. [Google Scholar]
- Choudhary, D.K.; Prakash, A.; Johri, B.N. Induced systemic resistance (ISR) in plants: Mechanism of action. Indian J. Microbiol. 2007, 47, 289–297. [Google Scholar] [CrossRef] [Green Version]
- Prabhukarthikeyan, S.R.; Keerthana, U.; Raguchander, T. Antibiotic-producing Pseudomonas fluorescens mediates rhizome rot disease resistance and promotes plant growth in turmeric plants. Microbiol. Res. 2018, 210, 65–73. [Google Scholar] [CrossRef]
- Boeckx, T.; Winters, A.L.; Webb, K.J.; Kingston-Smith, A.H. Polyphenol oxidase in leaves: Is there any significance to the chloroplastic localization? J. Exp. Bot. 2015, 66, 3571–3579. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Ming, J.; Deng, L.; Zeng, K. Effect of Pichia membranaefaciens in combination with salicylic acid on postharvest blue and green mold decay in citrus fruits. Biol. Control. 2014, 74, 21–29. [Google Scholar] [CrossRef]
- Liu, J.; Wisniewski, M.; Droby, S.; Vero, S.; Tian, S.; Hershkovitz, V. Glycine betaine improves oxidative stress tolerance and biocontrol efficacy of the antagonistic yeast Cystofilobasidium infirmominiatum. Int. J. Food Microbiol. 2011, 146, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Hernández, J.A.; Ferrer, M.A.; Jiménez, A.; Barceló, A.R.; Sevilla, F. Antioxidant systems and O2−/H2O2 production in the apoplast of pea leaves. Its relation with salt-induced necrotic lesions in minor veins. Plant. Physiol. 2001, 127, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Skrypnik, L.; Novikova, A.; Tokupova, E. Improvement of phenolic compounds, sssentialoOil content and antioxidant properties of sweet basil (Ocimum basilicum L.) depending on type and concentration of selenium application. Plants 2019, 8, 458. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Duan, X.; Jing, G.; OuYang, Q.; Tao, N. Cinnamaldehyde inhibits the mycelial growth of Geotrichum citri-aurantii and induces defense responses against sour rot in citrus fruit. Postharvest Biol. Technol. 2017, 129, 23–28. [Google Scholar] [CrossRef]
- Ge, Y.; Deng, H.; Bi, Y.; Li, C.; Liu, Y.; Dong, B. Postharvest ASM dipping and DPI pre-treatment regulated reactive oxygen species metabolism in muskmelon (Cucumis melo L.) fruit. Postharvest Biol. Technol. 2015, 99, 160–167. [Google Scholar] [CrossRef]
- Yan, Y.; Zhang, X.; Zheng, X.; Apaliya, M.T.; Yang, Q.; Zhao, L.; Gu, X.; Zhang, H. Control of postharvest blue mold decay in pears by Meyerozyma guilliermondii and it’s effects on the protein expression profile of pears. Postharvest Biol. Technol. 2018, 136, 124–131. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant. Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Ahima, J.; Zhang, X.; Yang, Q.; Zhao, L.; Tibiru, A.M.; Zhang, H. Biocontrol activity of Rhodotorula mucilaginosa combined with salicylic acid against Penicillium digitatum infection in oranges. Biol. Control. 2019, 135, 23–32. [Google Scholar] [CrossRef]
- Shah, K.; Nahakpam, S. Heat exposure alters the expression of SOD, POD, APX and CAT isozymes and mitigates low cadmium toxicity in seedlings of sensitive and tolerant rice cultivars. Plant. Physiol. Biochem. 2012, 57, 106–113. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, Y.; Wang, Y.; Li, B.; Gu, X.; Zhang, X.; Boateng, N.A.S.; Zhang, H. Effect of β-glucan on the biocontrol efficacy of Cryptococcus podzolicus against postharvest decay of pears and the possible mechanisms involved. Postharvest Biol. Technol. 2020, 160, 111057. [Google Scholar] [CrossRef]
- Han, J.; Zhao, L.; Zhu, H.; Dhanasekaran, S.; Zhang, X.; Zhang, H. Study on the effect of alginate oligosaccharide combined with Meyerozyma guilliermondii against Penicillium expansum in pears and the possible mechanisms involved. Physiol. Mol. Plant. Pathol. 2021, 115, 101654. [Google Scholar] [CrossRef]
- Preciado-Rangel, P.; Reyes-Pérez, J.J.; Ramírez-Rodríguez, S.C.; Salas-Pérez, L.; Fortis-Hernández, M.; Murillo-Amador, B.; Troyo-Diéguez, E. Foliar aspersion of salicylic acid improves phenolic and flavonoid compounds, and also the fruit yield in cucumber (Cucumis sativus L.). Plants 2019, 8, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Extracting Buffer | Reference |
|---|---|---|
| PPO | Phosphate buffer with 1% PVP and 1.33 mM ethylene diamine tetraacetic acid (EDTA) | Apaliya et al. [17] |
| POD | Phosphate buffer with 1% PVP and 1.33 mM EDTA | Wang, Y. et al. [18] |
| APX | Phosphate buffer with 0.1 mM EDTA, 1 mM ascorbic acid and 2% polyvinyl-poly-pyrrolidone (PVPP) | Wang, M. et al. [19] |
| SOD | Phosphate buffer with 5 mM dithiothreitol (DTT) and 5% PVP | Li and Yan [20] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lanhuang, B.; Yang, Q.; Godana, E.A.; Zhang, H. Efficacy of the Yeast Wickerhamomyces anomalus in Biocontrol of Gray Mold Decay of Tomatoes and Study of the Mechanisms Involved. Foods 2022, 11, 720. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11050720
Lanhuang B, Yang Q, Godana EA, Zhang H. Efficacy of the Yeast Wickerhamomyces anomalus in Biocontrol of Gray Mold Decay of Tomatoes and Study of the Mechanisms Involved. Foods. 2022; 11(5):720. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11050720
Chicago/Turabian StyleLanhuang, Boen, Qiya Yang, Esa Abiso Godana, and Hongyin Zhang. 2022. "Efficacy of the Yeast Wickerhamomyces anomalus in Biocontrol of Gray Mold Decay of Tomatoes and Study of the Mechanisms Involved" Foods 11, no. 5: 720. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11050720