
DNA-Based Method for Traceability and Authentication of Apis cerana and A. dorsata Honey (Hymenoptera: Apidae), Using the NADH dehydrogenase 2 Gene



Abstract
:
1. Introduction
2. Material and Methods
2.1. Schematic Overview of the Experimental Design
2.2. Designing Species-Specific Primers
2.3. Evaluate the Specificity and Sensitivity of Designed Primer Sets Using Bee DNA
- -
- DNA extraction from honeybees
- -
- Specificity and sensitivitytest of designed primers
- -
- Melting curve analysis by real-time PCR
2.4. PCR-Based Sensitivity Test of Honey Samples
- -
- Preparation of honey samples
- -
- DNA extraction from honey samples
- -
- Polymerase chain reaction (PCR)
2.5. Adulteration Analysis of the Honey Samples
- -
- Honey samples
- -
- DNA extraction and PCR-based authentication of honey samples
- -
- Data analysis
3. Results
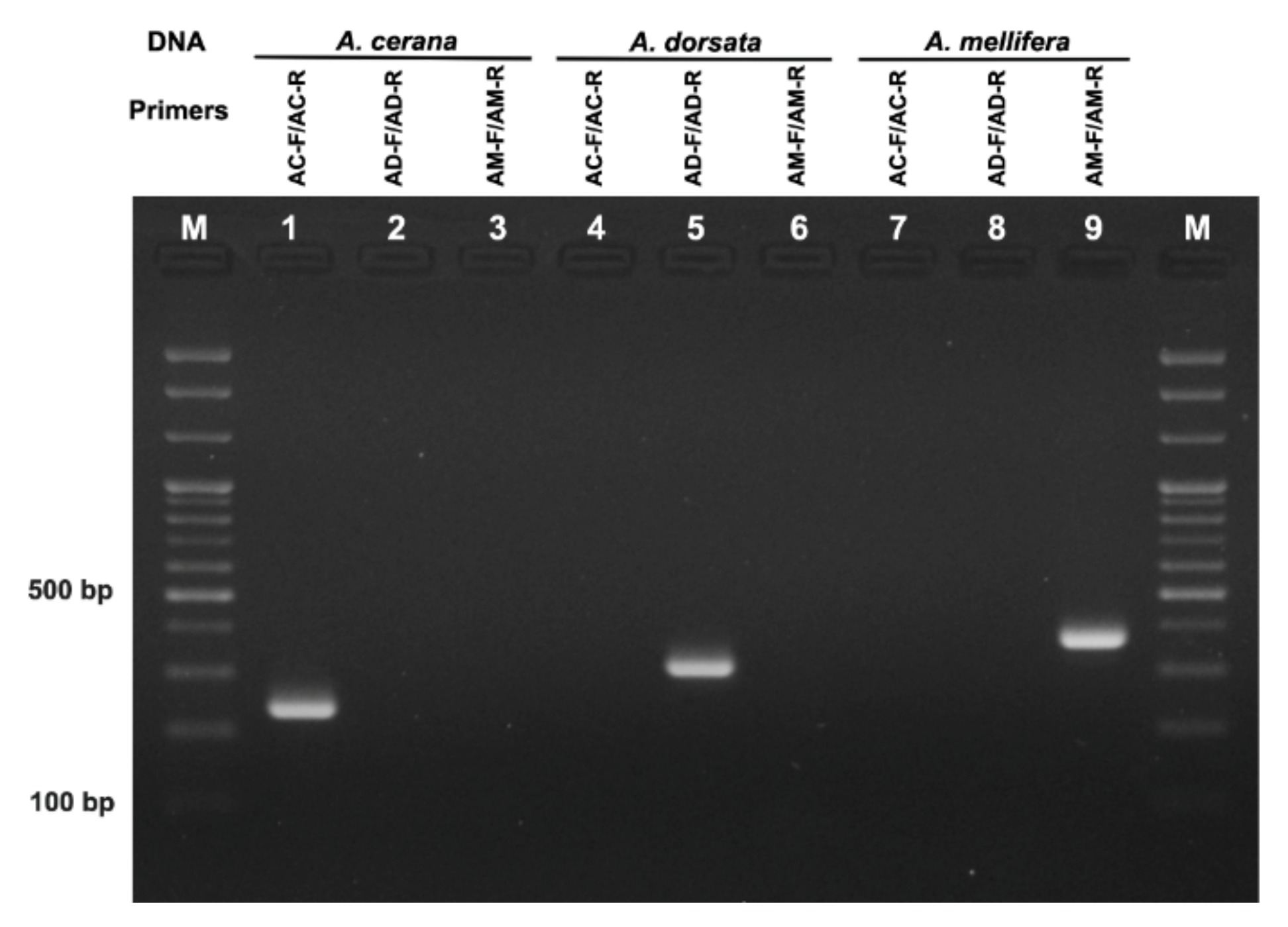
3.1. Specificity Test of Species-Specific Primers
3.2. Sensitivity Test of Primers Using Bee and Honey DNA
3.3. Melting Curve Analysis by Real-Time PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ajibola, A.; Chamunorwa, J.P.; Erlwanger, K.H. Dietary supplementation with natural honey promotes growth and health of male and female rats compared to cane syrup. Sci. Res. Essays 2013, 8, 543–553. [Google Scholar]
- Soares, S.; Amaral, J.S.; Oliveira, M.B.; Mafra, I. A comprehensive review on the main honey authentication issues: Production and origin. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1072–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldroyd, B.P.; Wongsiri, S. Asian Honey Bees: Biology, Conservation and Human Interactions; Harvard University Press: Cambridge, MA, USA, 2006. [Google Scholar]
- Wongsiri, S.; Chanchao, C.; Lekprayoon, C.; Wattanasermkit, K.; Deowanish, S.; Leepitakrat, S. Honeybee diversity and management in the new millennium in Thailand. In Proceedings of the 7th International Conference on Tropical Bees, Chiang Mai, Thailand, 19–25 March 2000; pp. 9–14. [Google Scholar]
- Corlett, R.T. Honeybees in natural ecosystems. In Honeybees of Asia; Hepburn, H.R., Radloff, S.E., Eds.; Springer-Verlag: Berlin, Germany, 2011; pp. 215–226. [Google Scholar]
- Partap, U. The pollination role of honeybees. In Honeybees of Asia; Hepburn, H.R., Radloff, S.E., Eds.; Springer-Verlag: Berlin, Germany, 2011; pp. 227–255. [Google Scholar]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Yong, P.L.; Othman, M.S.H. Economic Value of Honey Bees, Peninsular Malaysia; Forestry Department Peninsular Malaysia (FDPM): Kuala Lumpur, Malaysia, 2007. [Google Scholar]
- Moniruzzaman, M.; Khalil, M.I.; Sulaiman, S.A.; Gan, S.H. Physicochemical and antioxidant properties of Malaysian honeys produced by Apis cerana, Apis dorsata and Apis mellifera. BMC Complement. Altern. Med. 2013, 13, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruttner, F. Biogeography and Taxonomy of Honeybees; Springer: Heidelberg/Berlin, Germany, 1988. [Google Scholar]
- Jung, C.; Lee, M. Beekeeping in Korea: Past, present, and future challenges. In Asian Beekeeping in the 21st Century; Chantawannakul, P., Williams, G., Neumann, P., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 175–197. [Google Scholar]
- Lee, D.C.; Lee, S.Y.; Cha, S.H.; Choi, Y.S.; Rhee, H.I. Discrimination of native bee-honey and foreign bee-honey by SDS–PAGE. Korean J. Food Sci. Technol. 1998, 30, 1–5. [Google Scholar]
- He, X.; Wang, W.; Qin, Q.; Zeng, Z.; Zhang, S.; Barron, A.B. Assessment of flight activity and homing ability in Asian and European honey bee species, Apis cerana and Apis mellifera, measured with radio frequency tags. Apidologie 2012, 44, 38–51. [Google Scholar] [CrossRef] [Green Version]
- Jaafar, M.; Othman, M.; Yaacob, M.; Talip, B.; Ilyas, M.; Ngajikin, N.; Fauzi, N. A review on honey adulteration and the available detection approaches. J. Integr. Eng. 2020, 12, 125–131. [Google Scholar]
- Bogdanov, S.; Ruoff, K.; Oddo, L.P. Physico-chemical methods for the characterisation of unifloral honeys: A review. Apidologie 2004, 35 (Suppl. 1), S4–S17. [Google Scholar] [CrossRef] [Green Version]
- Sahinler, N.; Sahinler, S.; Gul, A. Biochemical composition of honeys produced in Turkey. J. Apic. Res. 2004, 43, 53–56. [Google Scholar] [CrossRef]
- Guler, A.; Bakan, A.; Nisbet, C.; Yavuz, O. Determination of important biochemical properties of honey to discriminate pure and adulterated honey with sucrose (Saccharum officinarum L.) syrup. Food Chem. 2007, 105, 1119–1125. [Google Scholar] [CrossRef]
- Chen, L.; Xue, X.; Ye, Z.; Zhou, J.; Chen, F.; Zhao, J. Determination of Chinese honey adulterated with high fructose corn syrup by near infrared spectroscopy. Food Chem. 2011, 128, 1110–1114. [Google Scholar] [CrossRef]
- Moore, J.C.; Spink, J.; Lipp, M. Development and application of a database of food ingredient fraud and economically motivated adulteration from 1980 to 2010. J. Food Sci. 2012, 77, R118–R126. [Google Scholar] [CrossRef]
- Bottero, M.T.; Dalmasso, A. Animal species identification in food products: Evolution of biomolecular methods. Vet. J. 2011, 190, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kumar, R.R.; Sharma, B.D.; Gokulakrishnan, P.; Mendiratta, S.K.; Sharma, D. Identification of species origin of meat and meat products on the DNA basis: A review. Crit. Rev. Food. Sci. Nutr. 2013, 55, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Amaral, J.; Meira, L.; Oliveira, M.; Mafra, I. Advances in authenticity testing for meat speciation. In Advances in Food Authenticity Testing; Downey, G., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 369–414. [Google Scholar]
- Willette, D.A.; Simmonds, S.E.; Cheng, S.H.; Esteves, S.; Kane, T.L.; Nuetzel, H.; Pilaud, N.; Rachmawati, R.; Barber, P.H. Using DNA barcoding to track seafood mislabeling in Los Angeles restaurants. Conserv. Biol. 2017, 31, 1076–1085. [Google Scholar] [CrossRef] [PubMed]
- Won, S.; Lee, D.; Ko, S.H.; Kim, J.; Rhee, H. Honey major protein characterization and its application to adulteration detection. Food Res. Int. 2008, 41, 952–956. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, S.; Chen, Y.; Wu, Y.; Tian, J.; Si, J.; Zhang, C.; Zheng, H.; Hu, F. Authentication of Apis cerana honey and Apis mellifera honey based on major royal jelly protein 2 gene. Molecules 2019, 24, 289. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-Z.; Chen, Y.-F.; Wu, Y.-Q.; Si, J.-J.; Zhang, C.-P.; Zheng, H.-O.; Hu, F.-L. Discrimination of the entomological origin of honey according to the secretions of the bee (Apis cerana or Apis mellifera). Food Res. Int. 2019, 116, 362–369. [Google Scholar] [CrossRef]
- Moritz, C.; Dowling, T.E.; Brown, W.M. Evolution of animal mitochondrial DNA: Relevance for population biology and systematics. Annu. Rev. Ecol. Evol. Syst. 1987, 18, 269–292. [Google Scholar] [CrossRef]
- Song, S.; Pursell, Z.F.; Copeland, W.C.; Longley, M.J.; Kunkel, T.A.; Mathews, C.K. DNA precursor asymmetries in mammalian tissue mitochondria and possible contribution to mutagenesis through reduced replication fidelity. Proc. Natl. Acad. Sci. USA 2005, 102, 4990–4995. [Google Scholar] [CrossRef] [Green Version]
- Zink, R.M.; Barrowclough, G.F. Mitochondrial DNA under siege in avian phylogeography. Mol. Ecol. 2008, 17, 2107–2121. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Lee, D.; Choi, S. Detection of Korean native honey and European honey by using duplex polymerase chain reaction and Immunochromatographic assay. Food Sci. Anim. Resour. 2017, 37, 599–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, S.; Grazina, L.; Mafra, I.; Costa, J.; Pinto, M.A.; Duc, H.P.; Oliviera, M.B.; Amaral, J. Novel diagnostic tools for Asian (Apis cerana) and European (Apis mellifera) honey authentication. Food Res. Int. 2018, 105, 686–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 20 December 2021).
- Kek, S.P.; Chin, N.L.; Tan, S.W.; Yusof, Y.A.; Chua, L.S. Molecular identification of honey entomological origin based on bee mitochondrial 16S rRNA and COI gene sequences. Food Control 2017, 78, 150–159. [Google Scholar] [CrossRef]
- Brudzynski, K.; Abubaker, K.; Miotto, D. Unraveling a mechanism of honey antibacterial action: Polyphenol/H2O2-induced oxidative effect on bacterial cell growth and on DNA degradation. Food Chem. 2012, 133, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Utzeri, V.J.; Ribani, A.; Fontanesi, L. Authentication of honey based on a DNA method to differentiate Apis mellifera subspecies: Application to Sicilian honey bee (A. M. siciliana) and iberian honey bee (A. M. iberiensis) honeys. Food Control 2018, 91, 294–301. [Google Scholar] [CrossRef]
- Bovo, S.; Utzeri, V.J.; Ribani, A.; Cabbri, R.; Fontanesi, L. Shortgun sequencing of honey DNA can describe honey bee derived environmental signatures and the honey bee hologenome complexity. Sci. Rep. 2020, 10, 9279. [Google Scholar] [CrossRef]
- Schnell, I.B.; Fraser, M.; Willerslev, E.; Gilbert, M.T.P. Characterisation of insect and plant origins using DNA extracted from small volumes of bee honey. Arthropod Plant Interact. 2010, 4, 107–116. [Google Scholar] [CrossRef]
- Jain, S.A.; Jesus, F.T.; Marchioro, G.M.; Araujo, E.D. Extraction of DNA from honey and its amplification by PCR for botanical identification. Food Sci. Technol. 2013, 33, 753–756. [Google Scholar] [CrossRef] [Green Version]
- Eimanifar, A.; Kimball, R.T.; Braun, E.L.; Ellis, J.D. The complete mitochondrial genome of the Cape honey bee Esch., Apis mellifera capensis (Insecta: Hymenoptera: Apidae). Mitochondrial DNA Part B 2016, 1, 817–819. [Google Scholar] [CrossRef] [Green Version]
- Eimanifar, A.; Kimball, R.T.; Braun, E.L.; Fuchs, S.; Grünewald, B.; Ellis, J.D. The complete mitochondrial genome of an east African honey bee, Apis mellifera monticola Smith (Insecta: Hymenoptera: Apidae). Mitochondrial DNA Part B 2017, 2, 589–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eimanifar, A.; Kimball, R.T.; Braun, E.L.; Moustafa, D.M.; Haddad, N.; Fuchs, S.; Grunewald, B.; Ellis, J.D. The complete mitochondrial genome of the Egyptian honey bee, Apis mellifera lamarckii (Insecta: Hymenoptera: Apidae). Mitochondrial DNA Part B 2017, 2, 270–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eimanifar, A.; Kimball, R.T.; Braun, E.L.; Fuchs, S.; Grunewald, B.; Ellis, J.D. The complete mitochondrial genome of Apis mellifera meda (Insecta: Hymenoptera: Apidae). Mitochondrial DNA Part B 2017, 2, 268–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eimanifar, A.; Kimball, R.T.; Braun, E.L.; Ellis, J.D. Mitochondrial genome diversity and population structure of two western honeybee subspecies in the Republic of South Africa. Sci. Rep. 2018, 8, 1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuller, Z.L.; Nino, E.L.; Patch, H.M.; Bedoya-Reina, O.C.; Baumgarten, T.; Muli, E.; Mumoki, F.; Ratan, A.; McGraw, J.; Frazier, M.; et al. Genome-wide analysis of signatures of selection in populations of African honey bees (Apis mellifera) using new web-based tools. BMC Genom. 2015, 16, 518. [Google Scholar] [CrossRef] [Green Version]
- Gibson, J.D.; Hunt, G.J. The complete mitochondrial genome of the invasive Africanized honey bee, Apis mellifera scutellata (Insecta: Hymenoptera: Apidae). Mitochondrial DNA Part A 2016, 27, 561–562. [Google Scholar] [CrossRef]
- Haddad, N.J. Mitochondrial genome of the Levant Region honey bee, Apis mellifera syriaca (Hymenoptera: Apidae). Mitochondrial DNA Part A 2016, 27, 4067–4068. [Google Scholar] [CrossRef]
- Hu, P.; Lu, Z.X.; Haddad, N.; Noureddine, A.; Loucif-Ayad, W.; Wang, Y.Z.; Zhang, R.B.Z.A.L.; Guan, X.; Zhang, H.X.; Niu, H. Complete mitochondrial genome of the Algerian honey bee, Apis mellifera intermissa (Hymenoptera: Apidae). Mitochondrial DNA Part A 2016, 27, 1791–1792. [Google Scholar]
- Ilyasov, R.A.; Park, J.; Takahashi, J.; Kwon, H.W. Phylogenetic uniqueness of honeybee Apis cerana from the Korean peninsula inferred from the mitochondrial, nuclear, and morphological data. J. Apicul. Sci. 2018, 62, 189–214. [Google Scholar] [CrossRef] [Green Version]
- Nakagawaa, I.; Maedaa, M.; Chikanoa, M.; Okuyamaa, H.; Murrayb, R.; Takahashia, J. The complete mitochondrial genome of the yellow coloured honeybee Apis mellifera (Insecta: Hymenoptera: Apidae) of New Zealand. Mitochondrial DNA Part B 2018, 3, 66–67. [Google Scholar] [CrossRef] [Green Version]
- Okuyama, H.; Hill, J.; Martin, S.J.; Takahashi, J. The complete mitochondrial genome of a Buckfast bee, Apis mellifera (Insecta: Hymenoptera: Apidae) in Northern Ireland. Mitochondrial DNA Part B 2018, 3, 338–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuyama, H.; Tingek, S.; Takahashi, J. The complete mitochondrial genome of the cavity-nesting honeybee, Apis cerana (Insecta: Hymenoptera: Apidae) from Borneo. Mitochondrial DNA Part B 2017, 2, 475–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuyama, H.; Jimi, R.; Wakamiya, T.; Takahashi, J. Complete mitochondrial genome of the honeybee Apis cerana native to two remote islands in Japan. Conserv. Genet. Resour. 2017, 9, 557–560. [Google Scholar] [CrossRef]
- Takahashi, J.; Deowanish, S.; Okuyama, H. Analysis of the complete mitochondrial genome of the giant honeybee, Apis dorsata, (Hymenoptera: Apidae) in Thailand. Conserv. Genet. Resour. 2017. [Google Scholar] [CrossRef]
- Takahashi, J.; Wakamiya, T.; Kiyoshi, T.; Uchiyama, H.; Yajima, S.; Kimura, K.; Nomura, T. The complete mitochondrial genome of the Japanese honeybee, Apis cerana japonica (Insecta: Hymenoptera: Apidae). Mitochondrial DNA Part B 2016, 1, 156–157. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.W.; Liu, G.H.; Dong, X.; Lin, R.Q.; Song, H.Q.; Huang, S.Y.; Yuan, Z.G.; Zhao, X.Q. The complete mitochondrial genome of the Asiatic cavity nesting honeybee Apis cerana (Hymenoptera: Apidae). PLoS ONE 2011, 6, e23008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, A.R.; Kim, J.S.; Kim, M.J.; Kim, H.K.; Choi, Y.S.; Kim, I. Comparative description of mitochondrial genomes of the honey bee Apis (Hymenoptera: Apidae): Four new genome sequences and Apis phylogeny using whole genomes and individual genes. J. Apic. Res. 2018, 57, 484–503. [Google Scholar] [CrossRef]
- Yang, J.; Xu, J.; Wu, J.; Zhang, X.; He, S. The complete mitogenome of wild honeybee Apis dorsata (Hymenoptera: Apidae) from South-Western China. Mitochondrial DNA Part B 2019, 4, 231–232. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Primer | 5′-3′ | Length | Target Fragment |
|---|---|---|---|---|
| A. cerana | AC-F | TCATTAGATTTTACAAAATCAGATCA | 26 | 224 bp |
| AC-R | CTTATAACTAAATATGTTAATGATCATA | 28 | ||
| A. dorsata | AD-F | TATATTAATTGTTATAACTTACATAAATAA | 31 | 302 bp |
| AD-R | GGATTAAGAATATATAATATTCATATTTT | 29 | ||
| A. mellifera | AM-F | CTATTAGATTTACTAAAACAGATACT | 26 | 377 bp |
| AM-R | ATAATTAAATGAATATAAAATAATTATAGCA | 31 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamadzade Namin, S.; Yeasmin, F.; Choi, H.W.; Jung, C. DNA-Based Method for Traceability and Authentication of Apis cerana and A. dorsata Honey (Hymenoptera: Apidae), Using the NADH dehydrogenase 2 Gene. Foods 2022, 11, 928. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11070928
Mohamadzade Namin S, Yeasmin F, Choi HW, Jung C. DNA-Based Method for Traceability and Authentication of Apis cerana and A. dorsata Honey (Hymenoptera: Apidae), Using the NADH dehydrogenase 2 Gene. Foods. 2022; 11(7):928. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11070928
Chicago/Turabian StyleMohamadzade Namin, Saeed, Fatema Yeasmin, Hyong Woo Choi, and Chuleui Jung. 2022. "DNA-Based Method for Traceability and Authentication of Apis cerana and A. dorsata Honey (Hymenoptera: Apidae), Using the NADH dehydrogenase 2 Gene" Foods 11, no. 7: 928. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11070928