
Metagenomic Analysis of the Effects of Lactiplantibacillus plantarum and Fructooligosaccharides (FOS) on the Fecal Microbiota Structure in Mice

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals Reagents, and Preparation of L. plantarum ST-III
2.2. Animal Design
2.3. SCFAs Analysis
2.4. DNA Extraction and Sequencing
2.5. Bioinformatics Analysis
2.6. Statistical Analysis
3. Results
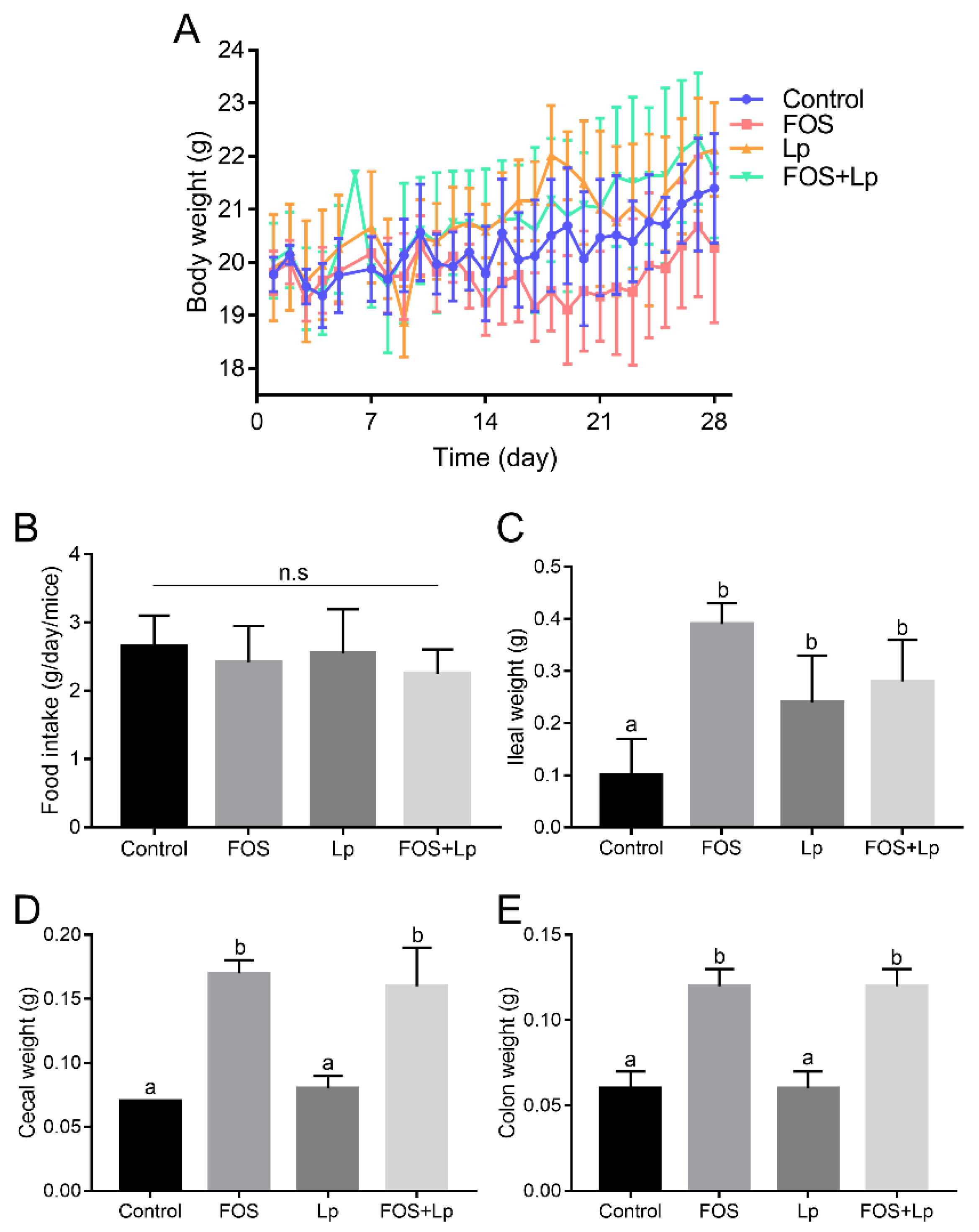
3.1. Influence of L. plantarum ST-III and FOS Intervention on Body Weight and Organ Index
3.2. Influence of L. plantarum ST-III and FOS Intervention on SCFAs
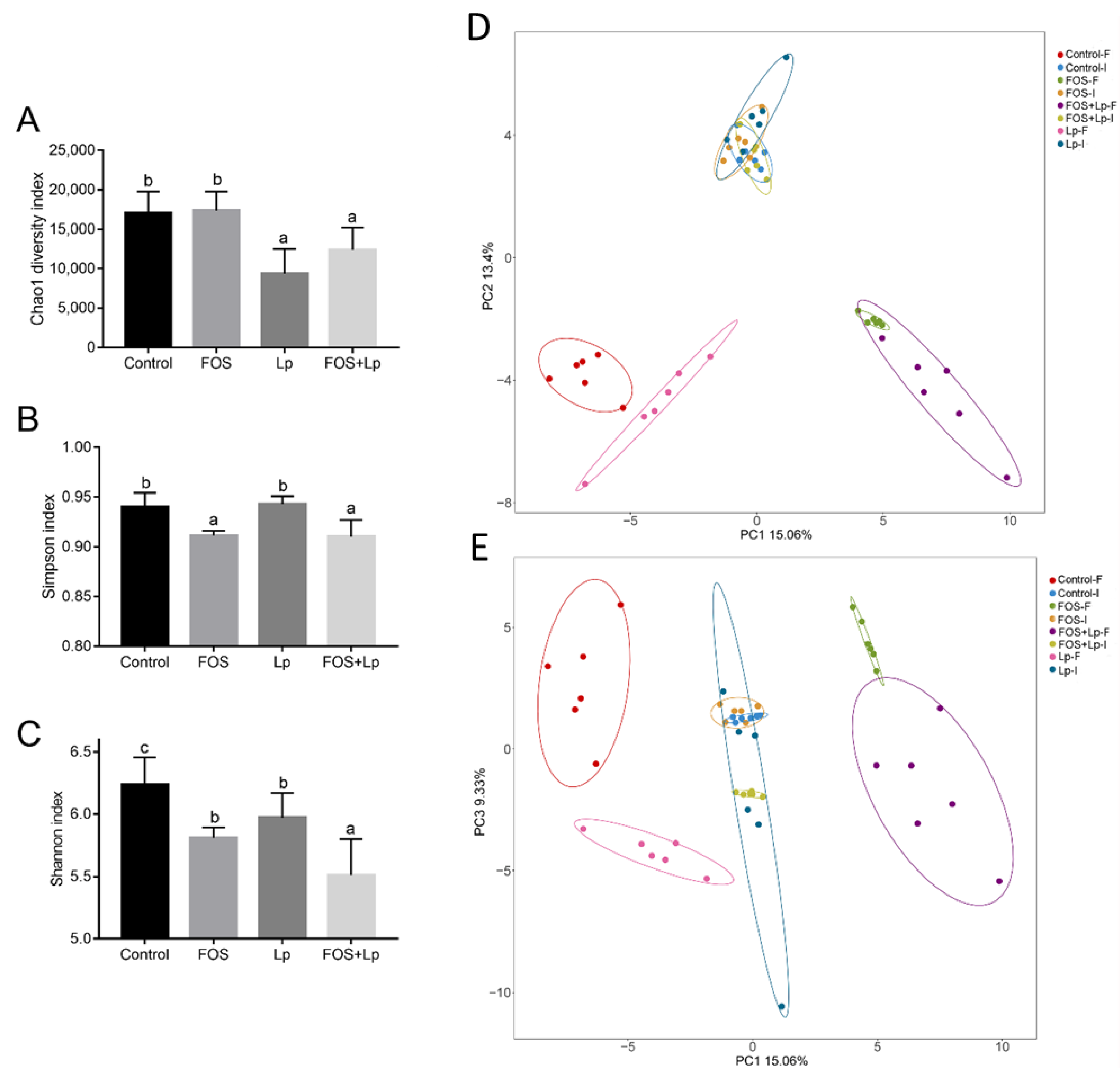
3.3. Alpha and Beta Diversity of Intestinal Microbiota
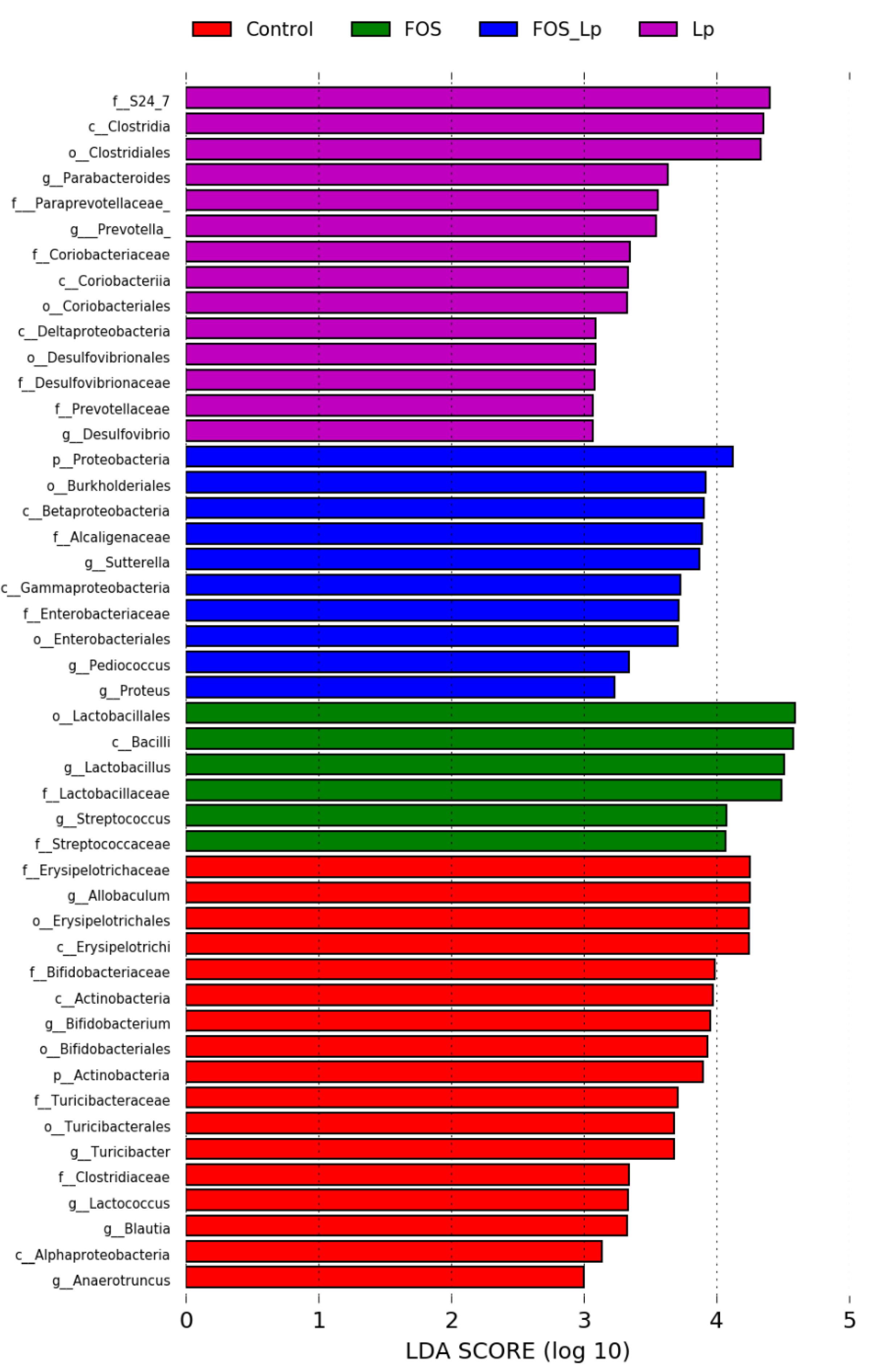
3.4. Influence of L. plantarum ST-III and FOS on the Abundance of Intestinal Microbiota
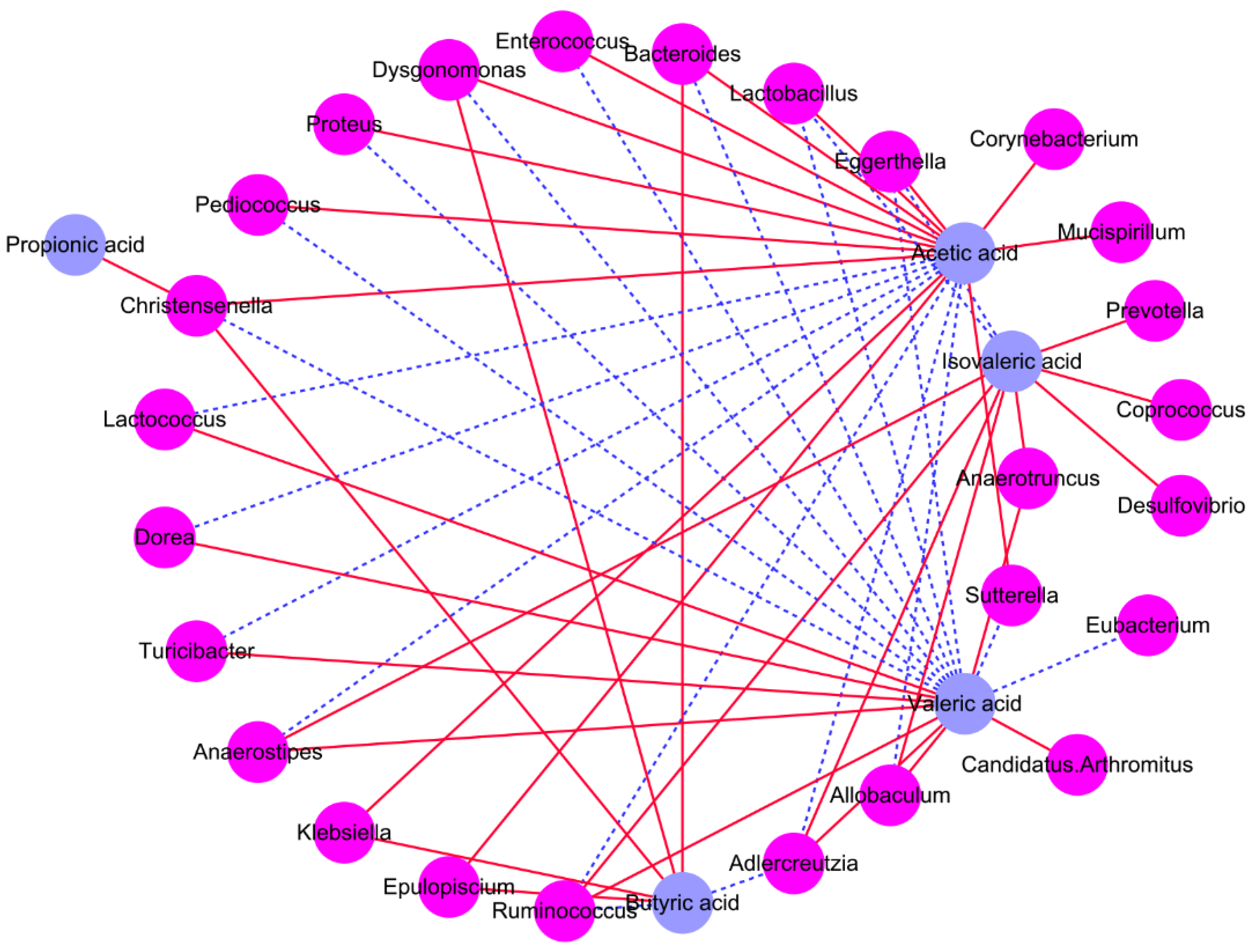
3.5. Association between Intestinal Microbiota and SCFAs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Choi, H.; Rao, M.C.; Chang, E.B. Gut microbiota as a transducer of dietary cues to regulate host circadian rhythms and metabolism. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Crovesy, L.; Masterson, D.; Rosado, E.L. Profile of the gut microbiota of adults with obesity: A systematic review. Eur. J. Clin. Nutr. 2020, 74, 1251–1262. [Google Scholar] [CrossRef] [PubMed]
- Lavelle, A.; Sokol, H. Gut microbiota-derived metabolites as key actors in inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Li, J.; Yin, J.; Li, C.; Zhang, S.; Guo, J.; Duan, J. Fructooligosaccharide decreases the production of uremic toxin precursor through modulating gut microbes mediated tyrosine metabolism pathway. Future Foods 2021, 4, 100069. [Google Scholar] [CrossRef]
- Mao, B.; Li, D.; Zhao, J.; Liu, X.; Gu, Z.; Chen, Y.Q.; Zhang, H.; Chen, W. Metagenomic insights into the effects of fructo-oligosaccharides (FOS) on the composition of fecal microbiota in mice. J. Agric. Food Chem. 2015, 63, 856–863. [Google Scholar] [CrossRef]
- Singhal, N.; Singh, N.S.; Mohanty, S.; Kumar, M.; Virdi, J.S. Rhizospheric Lactobacillus plantarum (Lactiplantibacillus plantarum) strains exhibit bile salt hydrolysis, hypocholestrolemic and probiotic capabilities in vitro. Sci. Rep. 2021, 11, 15288. [Google Scholar] [CrossRef]
- Li, X.; Xiao, Y.; Song, L.; Huang, Y.; Chu, Q.; Zhu, S.; Lu, S.; Hou, L.; Li, Z.; Li, J.; et al. Effect of Lactobacillus plantarum HT121 on serum lipid profile, gut microbiota, and liver transcriptome and metabolomics in a high-cholesterol diet–induced hypercholesterolemia rat model. Nutrition 2020, 79–80, 110966. [Google Scholar] [CrossRef]
- Chen, C.; Zhao, G.; Chen, W.; Guo, B. Metabolism of fructooligosaccharides in Lactobacillus plantarum ST-III via differential gene transcription and alteration of cell membrane fluidity. Appl. Environ. Microbiol. 2015, 81, 7697–7707. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Gu, Y.; Ren, H.; Wang, S.; Zhong, H.; Zhao, X.; Ma, J.; Gu, X.; Xue, Y.; Huang, S.; et al. Gut microbiome-related effects of berberine and probiotics on type 2 diabetes (the premote study). Nat. Commun. 2020, 11, 5015. [Google Scholar] [CrossRef]
- Guo, W.; Xiang, Q.; Mao, B.; Tang, X.; Cui, S.; Li, X.; Zhao, J.; Zhang, H.; Chen, W. Protective effects of microbiome-derived inosine on lipopolysaccharide-induced acute liver damage and inflammation in mice via mediating the TLR4/NF-κB pathway. J. Agric. Food Chem. 2021, 69, 7619–7628. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.; Chen, C.; Gu, J.; Mao, B.; Zhang, H.; Zhao, J.; Chen, W. Tracing Lactobacillus plantarum within the intestinal tract of mice: Green fluorescent protein-based fluorescent tagging. J. Sci. Food Agric. 2021, 101, 1758–1766. [Google Scholar] [CrossRef] [PubMed]
- Le Bourgot, C.; Ferret-Bernard, S.; Blat, S.; Apper, E.; Le Huërou-Luron, I. Short-chain fructooligosaccharide supplementation during gestation and lactation or after weaning differentially impacts pig growth and IgA response to influenza vaccination. J. Funct. Foods 2016, 24, 307–315. [Google Scholar] [CrossRef]
- Meksawan, K.; Chaotrakul, C.; Leeaphorn, N.; Gonlchanvit, S.; Eiam-Ong, S.; Kanjanabuch, T. Effects of Fructo-oligosaccharide supplementation on constipation in elderly continuous ambulatory peritoneal dialysis patients. Perit. Dial. Int. 2016, 36, 60–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Park, J.H.; Kim, I.H. Effects of probiotics containing (Lactobacillus planetarium) and chlortetracycline on growth performance, nutrient digestibility, fecal microflora, diarrhea score and fecal gas emission in weanling pigs. Livest. Sci. 2020, 241, 104186. [Google Scholar] [CrossRef]
- Fukasawa, T.; Kamei, A.; Watanabe, Y.; Koga, J.; Abe, K. Short-chain fructooligosaccharide regulates hepatic peroxisome proliferator-activated receptor α and farnesoid X receptor target gene expression in rats. J. Agric. Food Chem. 2010, 58, 7007–7012. [Google Scholar] [CrossRef]
- Ma, H.; Zhao, Y.; Li, L.; Liu, Y. Effects of combinations of goat milk and oligosaccharides on altering the microbiota, immune responses, and short chain fatty acid levels in the small intestines of mice. J. Agric. Food Chem. 2021, 69, 8828–8837. [Google Scholar] [CrossRef]
- Wang, M.; Wichienchot, S.; He, X.; Fu, X.; Huang, Q.; Zhang, B. In vitro colonic fermentation of dietary fibers: Fermentation rate, short-chain fatty acid production and changes in microbiota. Trends Food Sci. Technol. 2019, 88, 1–9. [Google Scholar] [CrossRef]
- Doo, E.H.; Schwab, C.; Chassard, C.; Lacroix, C. Cumulative effect of yeast extract and fructooligosaccharide supplementation on composition and metabolic activity of elderly colonic microbiota in vitro. J. Funct. Foods 2019, 52, 43–53. [Google Scholar] [CrossRef]
- McKenzie, C.I.; Mackay, C.R.; Macia, L. GPR43-A prototypic metabolite sensor linking metabolic and inflammatory diseases. Trends Endocrinol. Metab. 2015, 26, 511–512. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, X.; Zhu, L.; Yang, X.; He, F.; Wang, T.; Bao, T.; Lu, H.; Wang, H.; Yang, S. Inulin alleviates inflammation of alcoholic liver disease via SCFAs-inducing suppression of M1 and facilitation of M2 macrophages in mice. Int. Immunopharmacol. 2020, 78, 106062. [Google Scholar] [CrossRef]
- Chen, R.; Xu, Y.; Wu, P.; Zhou, H.; Lasanajak, Y.; Fang, Y.; Tang, L.; Ye, L.; Li, X.; Cai, Z.; et al. Transplantation of fecal microbiota rich in short chain fatty acids and butyric acid treat cerebral ischemic stroke by regulating gut microbiota. Pharmacol. Res. 2019, 148, 104403. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Yu, T.; Huang, X.; Bilotta, A.J.; Xu, L.; Lu, Y.; Sun, J.; Pan, F.; Zhou, J.; Zhang, W.; et al. Intestinal microbiota-derived short-chain fatty acids regulation of immune cell IL-22 production and gut immunity. Nat. Commun. 2020, 11, 4457. [Google Scholar] [CrossRef] [PubMed]
- Gentile, C.L.; Weir, T.L. The gut microbiota at the intersection of diet and human health. Science 2018, 362, 776–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Teng, H.; An, F.; Huang, Q.; Chen, L.; Song, H. The beneficial effects of purple yam (Dioscorea alata L.) resistant starch on hyperlipidemia in high-fat-fed hamsters. Food Funct. 2019, 10, 2642–2650. [Google Scholar] [CrossRef] [PubMed]
- Aranaz, P.; Ramos-Lopez, O.; Cuevas-Sierra, A.; Martinez, J.A.; Milagro, F.I.; Riezu-Boj, J.I. A predictive regression model of the obesity-related inflammatory status based on gut microbiota composition. Int. J. Obes. 2021, 45, 2261–2268. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Fan, S.T.; Huang, D.F.; Yu, Q.; Liu, X.Z.; Li, C.; Wang, S.; Xiong, T.; Nie, S.P.; Xie, M.Y. Effect of Lactobacillus plantarum NCU116 fermentation on asparagus officinalis polysaccharide: Characterization, antioxidative, and immunoregulatory activities. J. Agric. Food Chem. 2018, 66, 10703–10711. [Google Scholar] [CrossRef]
- Rasmussen, S.B.; Reinert, L.S.; Paludan, S.R. Innate recognition of intracellular pathogens: Detection and activation of the first line of defense. APMIS 2009, 117, 323–337. [Google Scholar] [CrossRef]
- Hadad, S.E.L.; Alsolami, M.; Aldahlawi, A.; Alrahimi, J.; Basingab, F.; Hassoubah, S.; Alothaid, H. In vivo evidence: Repression of mucosal immune responses in mice with colon cancer following sustained administration of Streptococcus thermophiles. Saudi J. Biol. Sci. 2021, 28, 4751–4761. [Google Scholar] [CrossRef]
- Cai, W.; Xu, J.; Li, G.; Liu, T.; Guo, X.; Wang, H.; Luo, L. Ethanol extract of propolis prevents high-fat diet-induced insulin resistance and obesity in association with modulation of gut microbiota in mice. Food Res. Int. 2020, 130, 108939. [Google Scholar] [CrossRef]
- Wang, Y.; You, Y.; Tian, Y.; Sun, H.; Li, X.; Wang, X.; Wang, Y.; Liu, J. Pediococcus pentosaceus PP04 ameliorates high-fat diet-induced hyperlipidemia by regulating lipid metabolism in C57BL/6N mice. J. Agric. Food Chem. 2020, 68, 15154–15163. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | FOS | Lp | FOS + Lp | |
|---|---|---|---|---|
| Acetic acid (μmol/g) | 10.92 ± 0.77 a | 35.23 ± 20.09 b | 12.35 ± 5.56 a | 31.63 ± 4.32 b |
| Propionic acid (μmol/g) | 3.05 ± 0.23 a | 7.15 ± 3.78 b | 2.91 ± 1.31 a | 5.04 ± 1.02 a |
| Butyric acid (μmol/g) | 0.96 ± 0.06 a | 8.05 ± 4.15 b | 1.38 ± 0.58 a | 10.53 ± 2.78 b |
| Valeric acid (μmol/g) | 0.77 ± 0.05 a | 2.91 ± 1.70 b | 0.80 ± 0.38 a | 1.42 ± 0.48 b |
| Isovaleric acid (μmol/g) | 0.70 ± 0.05 a | 1.70 ± 1.26 b | 1.24 ± 0.97 a,b | 0.89 ± 0.04 a,b |
| SCFAs (μmol/g) | 16.41 ± 1.10 a | 55.03 ± 30.92 b | 18.68 ± 8.80 a | 49.52 ± 7.83 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, S.; Guo, W.; Chen, C.; Tang, X.; Zhao, J.; Mao, B.; Zhang, H. Metagenomic Analysis of the Effects of Lactiplantibacillus plantarum and Fructooligosaccharides (FOS) on the Fecal Microbiota Structure in Mice. Foods 2022, 11, 1187. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11091187
Cui S, Guo W, Chen C, Tang X, Zhao J, Mao B, Zhang H. Metagenomic Analysis of the Effects of Lactiplantibacillus plantarum and Fructooligosaccharides (FOS) on the Fecal Microbiota Structure in Mice. Foods. 2022; 11(9):1187. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11091187
Chicago/Turabian StyleCui, Shumao, Weiling Guo, Cailing Chen, Xin Tang, Jianxin Zhao, Bingyong Mao, and Hao Zhang. 2022. "Metagenomic Analysis of the Effects of Lactiplantibacillus plantarum and Fructooligosaccharides (FOS) on the Fecal Microbiota Structure in Mice" Foods 11, no. 9: 1187. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11091187