
Disentangling the Tissue-Specific Variations of Volatile Flavor Profiles of the Lentinula edodes Fruiting Body
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Volatile Compound Extraction
2.3. Volatile Compound Measurements with GC–MS
2.4. Statistics
3. Results
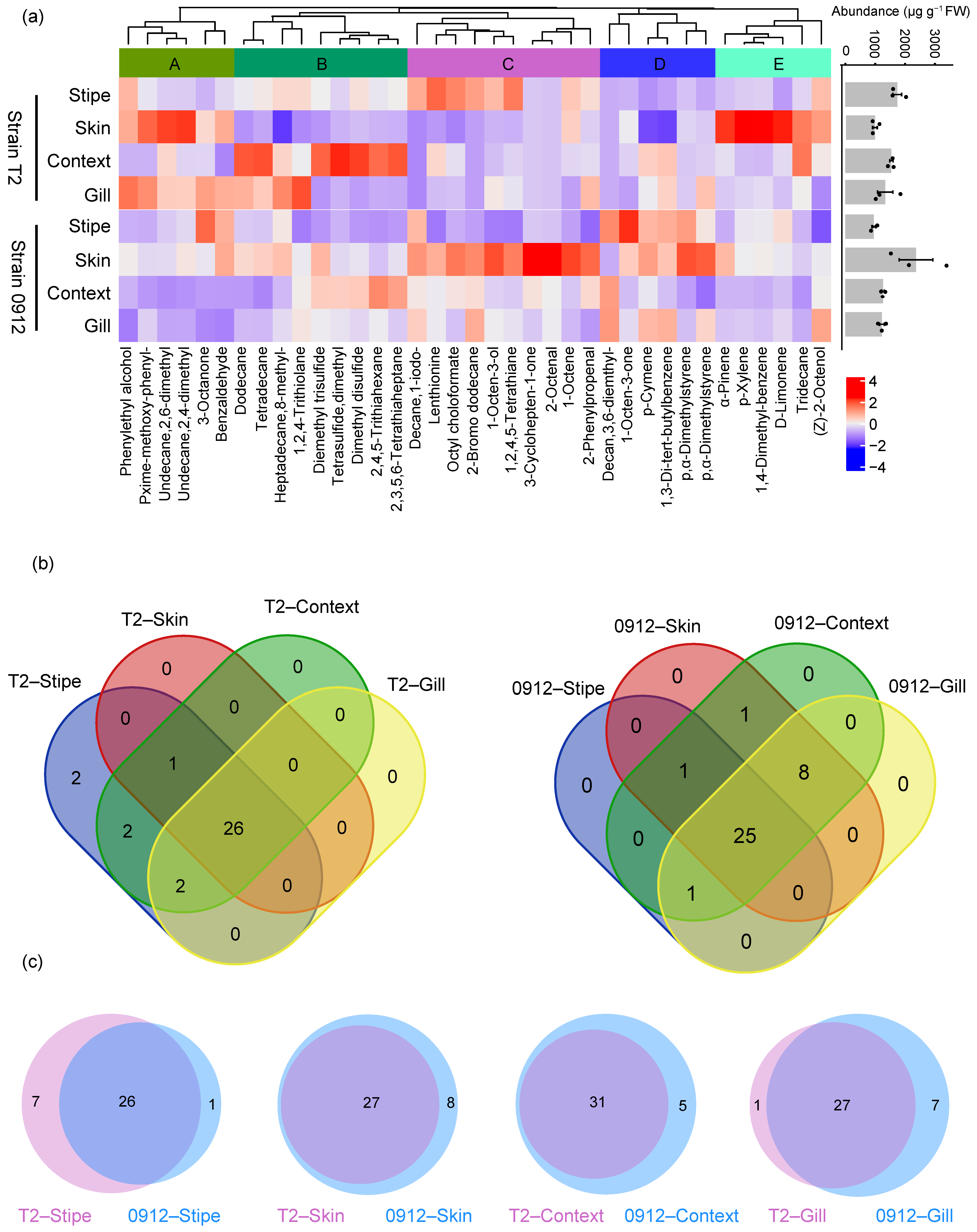
3.1. Overall Volatile Emissions from Different Tissues of L. edodes Fruiting Body
3.2. Chemical Diversity of the Volatile Compounds in Different Tissues of L. edodes Fruiting Body
3.3. The Profiles of C8 and Sulfur Compounds Differed in Different Tissues and Strains
3.4. Correlations of C8 and Sulfur Compounds
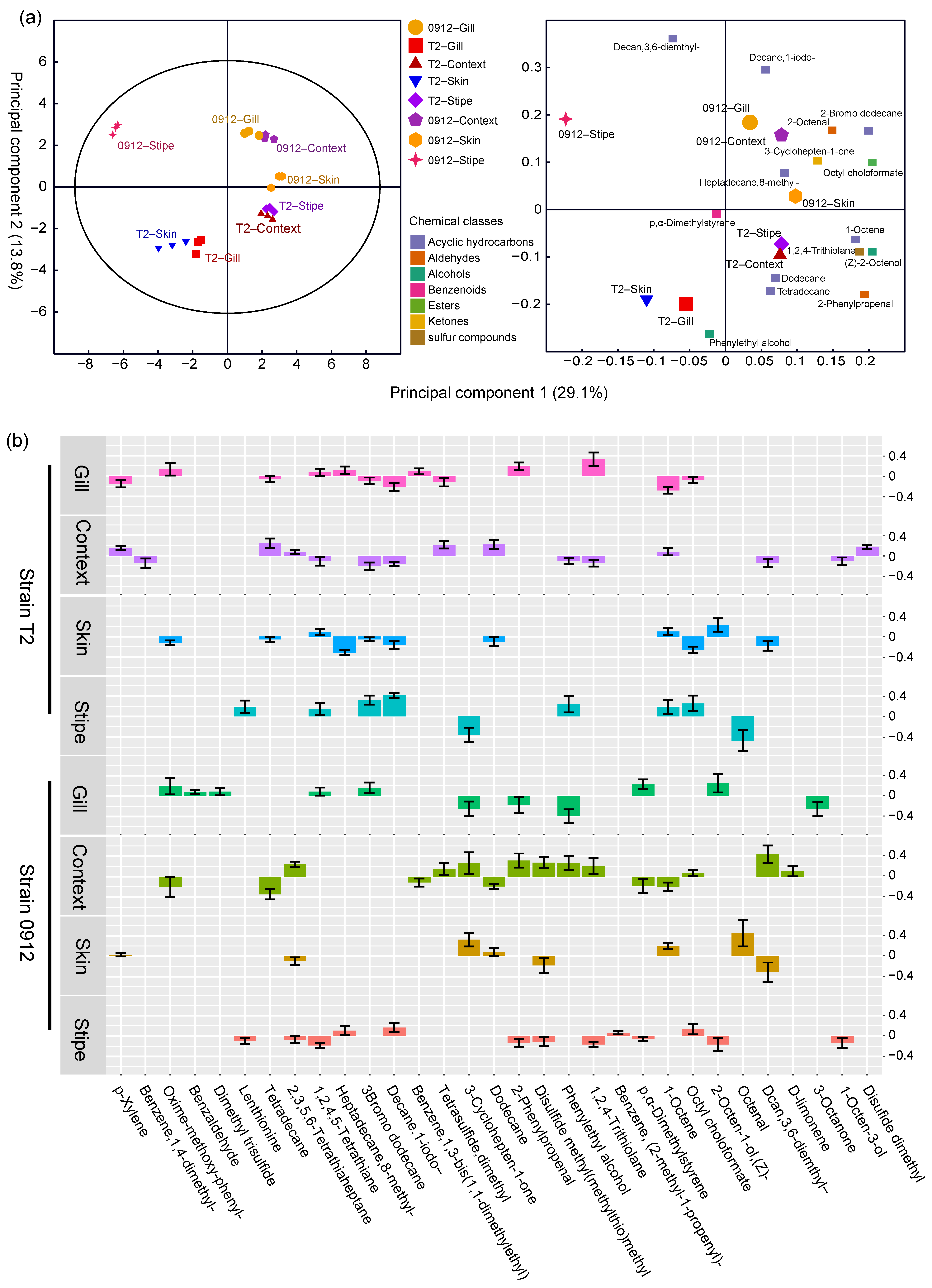
3.5. The Tissue-Dependent Volatile Profile Patterns Predict Different L. edodes Strains and Tissues
3.6. Volatile Biomarkers of L. edodes Strains and Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yu, H.; Zhang, L.; Shang, X.; Peng, B.; Li, Y.; Xiao, S.; Tan, Q.; Fu, Y. Chromosomal genome and population genetic analyses to reveal genetic architecture, breeding history and genes related to cadmium accumulation in Lentinula edodes. BMC Genom. 2022, 23, 120. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Gong, W.; Zhang, L.; Yang, Z.; Nong, W.; Bian, Y.; Kwan, H.-S.; Cheung, M.K.; Xiao, Y. Association mapping reveals genetic loci associated with important agronomic traits in Lentinula edodes, shiitake mushroom. Front. Microbiol. 2017, 8, 237. [Google Scholar] [CrossRef] [PubMed]
- China Edible Fungi Association. Available online: https://mp.weixin.qq.com/s/M6ZQN5IYkkMCOmsojfLtUw (accessed on 30 October 2023).
- Ahmad, I.; Arif, M.; Xu, M.; Zhang, J.; Ding, Y.; Lyu, F. Therapeutic values and nutraceutical properties of shiitake mushroom (Lentinula edodes): A review. Trends Food Sci. Technol. 2023, 134, 123–135. [Google Scholar] [CrossRef]
- Bisen, P.S.; Baghel, R.K.; Sanodiya, B.S.; Thakur, G.S.; Prasad, G.B. Lentinus edodes: A macrofungus with pharmacological activities. Curr. Med. Chem. 2010, 17, 2419–2430. [Google Scholar] [CrossRef] [PubMed]
- Finimundy, T.C.; Dillon, A.J.P.; Henriques, J.A.P.; Ely, M.R. A review on general nutritional compounds and pharmacological properties of the Lentinula edodes mushroom. Food Sci. Nutr. 2014, 5, 12. [Google Scholar]
- Chung, I.M.; Kim, S.Y.; Han, J.G.; Kong, W.-S.; Jung, M.Y.; Kim, S.H. Fatty acids and stable isotope ratios in shiitake mushrooms (Lentinula edodes) indicate the origin of the cultivation substrate used: A preliminary case study in Korea. Foods 2020, 9, 1210. [Google Scholar] [CrossRef]
- Chen, C.C.; Liu, S.E.; Wu, C.M.; Ho, C.T. Enzymic Formation of Volatile Compounds in Shiitake Mushroom (Lentinus edodes Sing.). In Biogeneration of Aromas; Parliament, T.H., Croteau, R., Eds.; ACS Symposium Series; ACS: Washington, DC, USA, 1986; Volume 317, pp. 176–183. [Google Scholar]
- Wu, C.M.; Wang, Z. Volatile compounds in fresh and processed shiitake mushrooms (Lentinus edodes Sing.). Food Sci. Technol. Res. 2000, 6, 166–170. [Google Scholar] [CrossRef]
- Cho, D.B.; Seo, H.Y.; Kim, K.S. Analysis of the volatile flavor compounds produced during the growth stages of the shiitake mushrooms (Lentinus edodes). Prev. Nutr. Food Sci. 2003, 8, 306–314. [Google Scholar] [CrossRef]
- Li, W.; Wang, J.; Chen, W.; Yang, Y.; Zhang, J.; Feng, J.; Yu, H.; Li, Q. Analysis of volatile compounds of Lentinula edodes grown in different culture substrate formulations. Food Res. Int. 2019, 125, 108517. [Google Scholar] [CrossRef]
- Sun, L.; Xin, G.; Hou, Z.; Zhao, X.; Xu, H.; Bao, X.; Xia, R.; Li, Y.; Li, L. Biosynthetic mechanism of key volatile biomarkers of harvested Lentinula edodes triggered by spore release. J. Agric. Food Chem. 2021, 69, 9350–9361. [Google Scholar] [CrossRef]
- Combet, E.; Eastwood, D.C.; Burton, K.S.; Henderson, J. Eight-carbon volatiles in mushrooms and fungi: Properties, analysis, and biosynthesis. Mycoscience 2006, 47, 317–326. [Google Scholar] [CrossRef]
- Xiao, Z.; Liu, L.; Niu, Y.; Zhang, J.; Wang, D.; Zhou, C. Mushroom alcohol (1-octen-3-ol) and other 7 aroma compounds selected from Chinese dry-cured hams can enhance saltiness perception. Meat Sci. 2023, 208, 109398. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, C.; Oliveira, I.; Silva, J.A.; Martins, C.; Ventanas, J.; García, C. Implementation of multivariate techniques for the selection of volatile compounds as indicators of sensory quality of raw beef. J. Food Sci. Technol. 2015, 52, 3887–3898. [Google Scholar] [CrossRef] [PubMed]
- Ahamed, Z.; Seo, J.K.; Eom, J.U.; Yang, H.S. Optimization of volatile compound extraction on cooked meat using HS-SPME-GC-MS, and evaluation of diagnosis to meat species using volatile compound by multivariate data analysis. LWT 2023, 188, 115374. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, L.; Liu, X.; Wang, Y.; Yang, W.; Zhao, W.; Zhao, G.; Cui, H.; Wen, J. Identification of characteristic aroma compounds in chicken meat and their metabolic mechanisms using gas chromatography-olfactometry, odor activity values, and metabolomics. Food Res. Int. 2023, 175, 113782. [Google Scholar] [CrossRef] [PubMed]
- Venkateshwarlu, G.; Chandravadana, M.; Tewari, R. Volatile flavour components of some edible mushrooms (Basidiomycetes). Flavour Fragr. J. 1999, 14, 191–194. [Google Scholar] [CrossRef]
- Zhu, R.; Wen, Y.; Wu, W.; Zhang, L.; Salman, F.M.; Shan, S.; Wen, J.; Farag, M.A.; Zhang, Y.; Zhao, C. The flavors of edible mushrooms: A comprehensive review of volatile organic compounds and their analytical methods. Crit. Rev. Food Sci. Nutr. 2022, 15, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Moliszewska, E. Mushroom flavour. Acta Univ. Lodz. Folia Biol. Oecol. 2014, 10, 80–88. [Google Scholar] [CrossRef]
- Yasumoto, K. Enzyme-catalyzed evolution of lenthionine from lenthinic acid. Agric. Biol. Chem. 1971, 35, 2070–2080. [Google Scholar] [CrossRef]
- Yasumoto, K.; Iwami, K.; Mitsuda, H. A new sulfur-containing peptide from Lentinus edodes acting as a precursor for lenthionine. Agric. Biol. Chem. 1971, 35, 2059–2069. [Google Scholar] [CrossRef]
- Li, W.; Li, R.; Chen, W.; Feng, J.; Wu, D.; Zhang, Z.; Zhang, J.; Yang, Y. The anabolism of sulphur aroma volatiles responds to enzymatic and non-enzymatic reactions during the drying process of shiitake mushrooms. Food Chem. 2022, 371, 131123. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, W.C.; Wang, J.B.; Feng, J.; Wu, D.; Zhang, Z.; Zhang, J.S.; Yang, Y. Effects of enzymatic reaction on the generation of key aroma volatiles in shiitake mushroom at different cultivation substrates. Food Sci. Nutr. 2021, 9, 2247–2256. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Bao, D.; Kang, Q.; Wu, Y. Research progress of volatiles in Lentinula edodes. Acta Edulis Fungi. 2018, 25, 105–114. [Google Scholar]
- Sneeden, E.Y.; Harris, H.H.; Pickering, I.J.; Prince, R.C.; Johnson, S.; Li, X.; Block, E.; George, G.N. The Sulfur Chemistry of Shiitake Mushroom. J. Am. Chem. Soc. 2004, 126, 458–459. [Google Scholar] [CrossRef] [PubMed]
- Hiraide, M.; Yokoyama, I. The smell and odorous components of dried shiitake mushroom, Lentinula edodes IV: Survey of trends in consumer preferences and changes in sensory evaluation. J. Wood Sci. 2007, 53, 458–461. [Google Scholar] [CrossRef]
- Hiraide, M.; Yokoyama, I.; Miyazaki, Y. The smell and odorous components of dried shiitake mushroom, Lentinula edodes II: Sensory evaluation by ordinary people. J. Wood Sci. 2005, 51, 628–633. [Google Scholar] [CrossRef]
- Hiraide, M.; Miyazaki, Y.; Shibata, Y. The smell and odorous components of dried shiitake mushroom, Lentinula edodes I: Relationship between sensory evaluations and amounts of odorous components. J. Wood Sci. 2004, 50, 358–364. [Google Scholar] [CrossRef]
- França, F.; dos Santos, H.-P.S.; Frasceto, R.A.; Saldana, E.; Lorenzo, J.M.; de Souza, V.T.M.F.; Selani, M.M. Umami ingredient from shiitake (Lentinula edodes) by-products as a flavor enhancer in low-salt beef burgers: Effects on physicochemical and technological properties. LWT 2022, 154, 112724. [Google Scholar] [CrossRef]
- Deng, G.; Li, J.; Liu, H.; Wang, Y. Volatile compounds and aroma characteristics of mushrooms: A review. Crit. Rev. Food Sci. Nutr. 2023, 3, 1–18. [Google Scholar] [CrossRef]
- Li, Y. Analytical methods for the analysis of volatile natural products. Nat. Prod. Rep. 2023, 40, 922–956. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, D.; Wu, Y.; Tong, X.; Qin, Y.; Wang, L. Difference in volatile aroma components of Stropharia rugosoannulata under two cultivated environments investigated by SPME-GC-MS. Foods 2023, 12, 2656. [Google Scholar] [CrossRef]
- Li, W.; Chen, W.; Wu, D.; Zhang, Z.; Yang, Y.; Zhang, J. Understanding the promotion of heat treatment on the flavor of Lentinula edodes using metabolomics integrated with transcriptomics. Food Res. Int. 2022, 162, 112051. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Dong, X.; Feng, X.; Ibrahim, S.A.; Huang, W.; Liu, Y. Effects of drying process on the volatile and non-volatile flavor compounds of Lentinula edodes. Foods 2021, 10, 2836. [Google Scholar] [CrossRef] [PubMed]
- Wurzenberger, M.; Grosch, W. Bestimmung von 1-oeten-3-ol in pilzen und pilzprodukten. Z. Lebensm. Unters. Forsch. 1983, 176, 16–19. [Google Scholar] [CrossRef]
- Tasaki, Y.; Kobayashi, D.; Sato, R.; Hayashi, S.; Joh, T. Variations in 1-octen-3-ol and lipoxygenase gene expression in the oyster mushroom Pleurotus ostreatus according to fruiting body development, tissue specificity, maturity, and postharvest storage. Mycoscience 2019, 60, 170–176. [Google Scholar] [CrossRef]
- Chen, D.; Wang, S.; Li, M.; Hao, T.; Lin, S. The dynamic changes in product attributes of shiitake mushroom pilei and stipes during dehydration by hot air drying. J. Food Process. Preserv. 2021, 45, e15648. [Google Scholar] [CrossRef]
- Li, S.; Wang, A.; Liu, L.; Tian, G.; Wei, S.; Xu, F. Evaluation of nutritional values of shiitake mushroom (Lentinus edodes) stipes. J. Food Meas. Charact. 2018, 12, 2012–2019. [Google Scholar] [CrossRef]
- van Den Dool, H.; Kratz, P.D. A generalization of the retention index system including linear temperature programmed gas-liquid partition chromatography. J. Chromatogr. 1963, 11, 463–471. [Google Scholar] [CrossRef]
- Guo, Y.; Ghirardo, A.; Weber, B.; Schnitzler, J.P.; Benz, J.P.; Rosenkranz, M. Trichoderma species differ in their volatile profiles and in antagonism toward ectomycorrhiza Laccaria bicolor. Front. Microbiol. 2019, 10, 891. [Google Scholar] [CrossRef]
- Kreuzwieser, J.; Scheerer, U.; Kruse, J.; Burzlaff, T.; Honsel, A.; Alfarraj, S.; Georgiev, P.; Schnitzler, J.P.; Ghirardo, A.; Kreuzer, I.; et al. The Venus flytrap attracts insects by the release of volatile organic compounds. J. Exp. Bot. 2014, 65, 755–766. [Google Scholar] [CrossRef]
- Xu, L.; Fang, X.; Wu, W.; Chen, H.; Mu, H.; Gao, H. Effects of high-temperature pre-drying on the quality of air-dried shiitake mushrooms (Lentinula edodes). Food Chem. 2019, 285, 406–413. [Google Scholar] [CrossRef] [PubMed]
- RStudio. RStudio: Integrated Development Environment for R; PBC: Boston, MA, USA, 2023. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 20 October 2023).
- Wei, T.; Simko, V. Package ‘corrplot’: Visualization of a Correlation Matrix, version 0.92. 2021. Available online: https://github.com/taiyun/corrplot (accessed on 20 October 2023).
- Pedersen, T. ggraph: An Implementation of Grammar of Graphics for Graphs and Networks. 2022. Available online: https://github.com/thomasp85/ggraph (accessed on 20 October 2023).
- Fruchterman, T.M.; Reingold, E.M. Graph drawing by force-directed placement. Softw. Pract. Exp. 1991, 21, 1129–1164. [Google Scholar] [CrossRef]
- Csárdi, G.; Nepusz, T.; Traag, V.; Horvát, S.; Zanini, F.; Noom, D.; Müller, K. igraph: Network Analysis and Visualization in R. R Package Version 1.6.0. 2023. Available online: https://CRAN.R-project.org/package=igraph (accessed on 20 October 2023).
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Darst, B.F.; Malecki, K.C.; Engelman, C.D. Using recursive feature elimination in random forest to account for correlated variables in high dimensional data. BMC Genet. 2018, 19, 65. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Jud, W.; Ghirardo, A.; Antritter, F.; Benz, J.P.; Schnitzler, J.P.; Rosenkranz, M. Sniffing fungi–phenotyping of volatile chemical diversity in Trichoderma species. New Phyto. 2020, 227, 244–259. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, M. Building predictive models in R using the caret package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar] [CrossRef]
- Deviaene, M.; Testelmans, D.; Borzée, P.; Buyse, B.; Van Huffel, S.; Varon, C. Feature selection algorithm based on random forest applied to sleep apnea detection. In Proceedings of the 2019 41st Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Berlin, Germany, 23–27 July 2019; pp. 2580–2583. [Google Scholar]
- Qin, L.; Gao, J.X.; Xue, J.; Chen, D.; Lin, S.Y.; Dong, X.P.; Zhu, B.W. Changes in aroma profile of Shiitake mushroom (Lentinus edodes) during different stages of hot air drying. Foods 2020, 9, 444. [Google Scholar] [CrossRef]
- Xie, L.; Jiang, Y.S.; Wang, Y.B.; Xiao, H.W.; Liu, W.; Ma, Y.; Zhao, X.Y. Changes in the physical properties and volatile odor characteristics of shiitake mushrooms (Lentinula edodes) in far infrared radiation drying. Foods 2023, 12, 3213. [Google Scholar] [CrossRef]
- Zhuang, J.; Xiao, Q.; Feng, T.; Huang, Q.; Ho, C.T.; Song, S. Comparative flavor profile analysis of four different varieties of Boletus mushrooms by instrumental and sensory techniques. Food Res. Int. 2020, 136, 109485. [Google Scholar] [CrossRef]
- Luo, D.; Wu, J.; Ma, Z.; Tang, P.; Liao, X.; Lao, F. Production of high sensory quality Shiitake mushroom (Lentinus edodes) by pulsed air-impingement jet drying (AID) technique. Food Chem. 2021, 341, 128290. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, Z.; Chen, X.; Han, D.; Han, J.; Wang, L.; Ren, A.; Yu, H.; Zhao, M. Lenthionine, a key flavor substance in Lentinula edodes, is regulated by cysteine under drought Stress. J. Agric. Food Chem. 2021, 69, 12645–12653. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, W.-C.; Wang, J.-B.; Feng, J.; Wu, D.; Zhang, Z.; Zhang, J.-S.; Yang, Y. Screening candidate genes related to volatile synthesis in shiitake mushrooms and construction of regulatory networks to effectively improve mushroom aroma. J. Sci. Food Agric. 2021, 101, 5618–5626. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.K.; Brunton, N.P.; Lyng, J.G.; Harrison, S.M.; Carpes, S.T.; Papoutsis, K. Volatile and non-volatile compounds of shiitake mushrooms treated with pulsed light after twenty-four hour storage at different conditions. Food Biosci. 2020, 36, 100619. [Google Scholar]
- Xu, S.; Wang, F.; Fu, Y.; Li, D.; Sun, X.; Li, C.; Song, B.; Li, Y. Effects of mixed agro-residues (corn crop waste) on lignin-degrading enzyme activities, growth, and quality of Lentinula edodes. RSC Adv. 2020, 10, 9798–9807. [Google Scholar] [CrossRef]
- Usami, A.; Nakaya, S.; Nakahashi, H.; Miyazawa, M. Chemical composition and aroma evaluation of volatile oils from edible mushrooms (Pleurotus salmoneostramineus and Pleurotus sajor-caju). J. Oleo Sci. 2014, 63, 1323–1332. [Google Scholar] [CrossRef]
- Feng, T.; Yang, M.; Ma, B.; Zhao, Y.; Zhuang, H.; Zhang, J.; Chen, D. Volatile profiles of two genotype Agaricus bisporus species at different growth stages. Food Res. Int. 2021, 140, 109761. [Google Scholar] [CrossRef]
- Culleré, L.; Ferreira, V.; Venturini, M.E.; Marco, P.; Blanco, D. Potential aromatic compounds as markers to differentiate between Tuber melanosporum and Tuber indicum truffles. Food Chem. 2013, 141, 105–110. [Google Scholar] [CrossRef]







{kind=link}
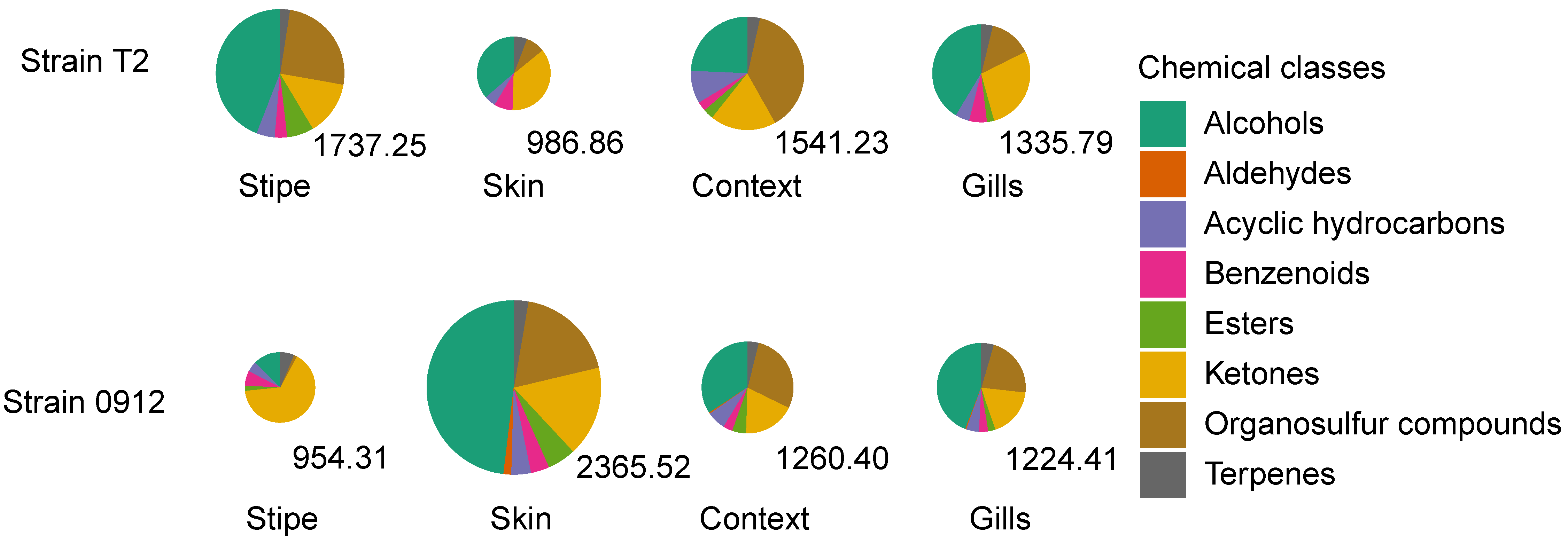{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Models | Accuracy (%) | Sensitivity (%) | Specificity (%) | Accuracy p-Value | Cohen’s Kappa (%) |
|---|---|---|---|---|---|
| Strain T2 vs. 0912 | 100 | 100 | 100 | 0.016 | 100 |
| Different tissues of strain T2 | 100 | 0.004 | 100 | ||
| Stipe | 100 | 100 | 100 | ||
| Skin | 100 | 100 | 100 | ||
| Context | 100 | 100 | 100 | ||
| Gill | 100 | 100 | 100 | ||
| Different tissues of strain 0912 | 0.050 | 67 | |||
| Stipe | 100 | 100 | 100 | ||
| Skin | 50 | 0 | 100 | ||
| Context | 100 | 100 | 100 | ||
| Gill | 83 | 100 | 67 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Zhao, J.; Wei, H.; Gao, Q.; Song, S.; Fan, Y.; Yan, D.; Liu, Y.; Wang, S. Disentangling the Tissue-Specific Variations of Volatile Flavor Profiles of the Lentinula edodes Fruiting Body. Foods 2024, 13, 86. https://0-doi-org.brum.beds.ac.uk/10.3390/foods13010086
Guo Y, Zhao J, Wei H, Gao Q, Song S, Fan Y, Yan D, Liu Y, Wang S. Disentangling the Tissue-Specific Variations of Volatile Flavor Profiles of the Lentinula edodes Fruiting Body. Foods. 2024; 13(1):86. https://0-doi-org.brum.beds.ac.uk/10.3390/foods13010086
Chicago/Turabian StyleGuo, Yuan, Jing Zhao, Huixian Wei, Qi Gao, Shuang Song, Yangyang Fan, Dong Yan, Yu Liu, and Shouxian Wang. 2024. "Disentangling the Tissue-Specific Variations of Volatile Flavor Profiles of the Lentinula edodes Fruiting Body" Foods 13, no. 1: 86. https://0-doi-org.brum.beds.ac.uk/10.3390/foods13010086