
Antilisterial Effectiveness of Origanum vulgare var. hirtum and Coridothymus capitatus Essential Oils and Hydrolates Alone and in Combination
 ,
,  , , and
, , and 
Abstract
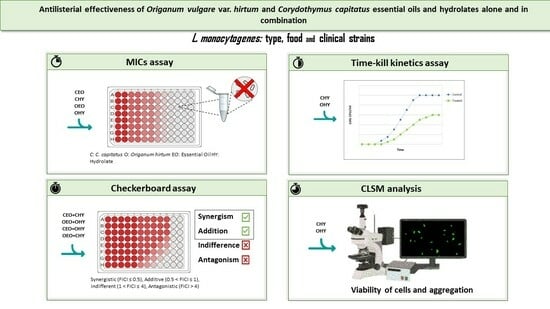
:
1. Introduction
2. Materials and Methods
2.1. Antimicrobial Compounds
2.2. Bacterial Strains and Culture Collection
2.3. Minimum Inhibitory Concentration Determination
2.4. Time-Kill Kinetics Assay
2.5. Confocal Laser Scanning Microscope Analysis
2.6. Synergistic Interaction Evaluation
2.7. Data Analysis
3. Results and Discussion
3.1. EOs and HYs Chemical Composition
3.2. Minimum Inhibitory Concentration Determination
3.3. Time-Kill Kinetics Assay
3.4. Confocal Laser Scanning Microscope Observation
3.5. Evaluation of the Synergistic Interaction of EO and HY Pairs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oraon, L.A.; Jana, P.S.; Prajapati, P.S. Application of herbs in functional dairy products—A review. J. Dairy Vet. Anim. Res. 2017, 5, 109. [Google Scholar]
- Delgado, A.; Gonçalves, S.; Romano, A. Mediterranean Diet: The Role of Phenolic Compounds from Aromatic Plant Foods. Foods 2023, 12, 840. [Google Scholar] [CrossRef]
- Hammoudi Halat, D.; Krayem, M.; Khaled, S.; Younes, S. A Focused Insight into Thyme: Biological, Chemical, and Therapeutic Properties of an Indigenous Mediterranean Herb. Nutrients 2022, 14, 2104. [Google Scholar] [CrossRef]
- Rossi, C.; Chaves-López, C.; Serio, A.; Casaccia, M.; Maggio, F.; Paparella, A. Effectiveness and mechanisms of essential oils for biofilm control on food-contact surfaces: An updated review. Crit. Rev. Food Sci. Nutr. 2020, 62, 2172. [Google Scholar] [CrossRef]
- Değirmenci, H.; Erkurt, H. Relationship between volatile components, antimicrobial and antioxidant properties of the essential oil, hydrosol and extracts of Citrus aurantium L. flowers. J. Infect. Public Health 2020, 13, 58. [Google Scholar] [CrossRef]
- Paparella, A.; Mazzarrino, G.; Chaves-Lopez, C.; Rossi, C.; Sacchetti, G.; Guerrieri, O.; Serio, A. Chitosan boosts the antimicrobial activity of Origanum vulgare essential oil in modified atmosphere packaged pork. Food Microbiol. 2016, 59, 23. [Google Scholar] [CrossRef]
- Rossi, C.; Maggio, F.; Chaves-López, C.; Valbonetti, L.; Berrettoni, M.; Paparella, A.; Serio, A. Effectiveness of selected essential oils and one hydrolate to prevent and remove Listeria monocytogenes biofilms on polystyrene and stainless steel food-contact surfaces. J. Appl. Microbiol. 2021, 132, 1866. [Google Scholar] [CrossRef] [PubMed]
- Heimesaat, M.M.; Mousavi, S.; Weschka, D.; Bereswill, S.S. Anti-Pathogenic and Immune-Modulatory Effects of Peroral Treatment with Cardamom Essential Oil in Acute Murine Campylobacteriosis. Microorganisms 2021, 9, 169. [Google Scholar] [CrossRef] [PubMed]
- Serra, E.; Saubade, F.; Ligorio, C.; Whitehead, K.; Sloan, A.; Williams, D.W.; Hidalgo-Bastida, A.; Verran, J.; Malic, S. Methylcellulose Hydrogel with Melissa officinalis Essential Oil as a Potential Treatment for Oral Candidiasis. Microorganisms 2020, 8, 215. [Google Scholar] [CrossRef] [PubMed]
- Di Vito, M.; Smolka, M.R.A.; Proto, L.; Barbanti, F.; Gelmini, E.; Napoli, M.G.; Bellardi, P.; Mattarelli, G.; Beretta, M.; Sanguinetti, F.; et al. Is the Antimicrobial Activity of Hydrolates Lower than That of Essential Oils? Antibiotics 2021, 10, 88. [Google Scholar] [CrossRef] [PubMed]
- Iordache, A.M.; Nechita, C.; Voica, C.; Roba, C.; Botoran, O.R.; Ionete, R.E. Assessing the Health Risk and the Metal Content of Thirty-Four Plant Essential Oils Using the ICP-MS Technique. Nutrients 2022, 14, 2363. [Google Scholar] [CrossRef]
- Rivera-Pérez, A.; Romero-González, R.; Garrido Frenich, A. Persistent organic pollutants (PCBs and PCDD/Fs), PAHs, and plasticizers in spices, herbs, and tea—A review of chromatographic methods from the last decade (2010–2020). Crit. Rev. Food Sci. Nutr. 2022, 62, 5224–5244. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Kim, H.; Beuchat, L.R.; Ryu, J.H. Synergistic activities of gaseous oregano and thyme thymol essential oils against Listeria monocytogenes on surfaces of a laboratory medium and radish sprouts. Food Microbiol. 2020, 86, 103357. [Google Scholar] [CrossRef]
- D’Amato, S.; Serio, A.; Chaves-López, C.; Paparella, A. Hydrosols: Biological activity and potential as antimicrobials for food applications. Food Control 2018, 86, 126. [Google Scholar] [CrossRef]
- Acimovic, M.; Tešević, V.; Smiljanic, K.; Cvetkovic, M.; Stankovic, J.; Kiprovski, B.; Sikora, V. Hydrolates: By-products of essential oil distillation: Chemical composition, biological activity and potential uses. Adv. Technol. 2020, 9, 54. [Google Scholar] [CrossRef]
- Politi, M.; Menghini, L.; Conti, B.; Bedini, S.; Farina, P.; Cioni, P.L.; Braca, A.; Leo, M. Reconsidering Hydrosols as Main Products of Aromatic Plants Manufactory: The Lavandin (Lavandula × intermedia) Case Study in Tuscany. Molecules 2020, 25, 2225. [Google Scholar] [CrossRef]
- Mazzarrino, G.; Paparella, A.; Chaves-López, C.; Faberi, A.; Sergi, M.; Sigismondi, C.; Compagnone, D.; Serio, A. Salmonella enterica and Listeria monocytogenes inactivation dynamics after treatment with selected essential oils. Food Control 2015, 50, 794–803. [Google Scholar] [CrossRef]
- Tardugno, R.; Serio, A.; Pellati, F.; D’Amato, S.; Chaves López, C.; Bellardi, M.G.; Di Vito, M.; Savini, V.; Paparella, A.; Benvenuti, S. Lavandula × intermedia and Lavandula angustifolia essential oils: Phytochemical composition and antimicrobial activity against foodborne pathogens. Nat. Prod. Res. 2019, 33, 3330. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI 447 Supplement M100S; CLSI: Wayne, PA, USA, 2020. [Google Scholar]
- Qiu, Y.; Chan, S.T.; Lin, L.; Shek, T.L.; Tsang, T.F.; Zhang, Y.; Ip, M.; Chan, P.K.; Blanchard, N.; Hanquet, G.; et al. Nusbiarylins, a new class of antimicrobial agents: Rational design of bacterial transcription inhibitors targeting the interaction between the NusB and NusE proteins. Bioorg. Chem. 2019, 92, 103203. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing; 25th Informational Supplement M100S25; CLSI: Wayne, PA, USA, 2015. [Google Scholar]
- Tyagi, P.; Singh, M.; Kumari, H.; Kumari, A.; Mukhopadhyay, K. Bactericidal activity of CurcuminI is associated with damaging of bacterial membrane. PLoS ONE 2015, 10, e0121313. [Google Scholar] [CrossRef]
- Fratini, F.; Mancini, S.; Turchi, B.; Friscia, E.; Pistelli, L.; Giusti, G.; Cerri, D. A novel interpretation of the fractional inhibitory concentration index: The case Origanum vulgare L. and Leptospermum scoparium J. R. et G. Forst essential oils against Staphylococcus aureus strains. Microbiol. Res. 2017, 195, 11. [Google Scholar] [CrossRef]
- Vasilijević, B.; Mitić-Ćulafić, D.; Djekic, I.; Marković, T.; Knežević-Vukčević, J.; Tomasevic, I.; Velebit, B.; Nikolić, B. Antibacterial effect of Juniperus communis and Satureja montana essential oils against Listeria monocytogenes in vitro and in wine marinated beef. Food Control 2019, 100, 247. [Google Scholar] [CrossRef]
- Granger, R.; Passet, J.; Verdier, R. Gamma-terpinene, precursor of p-cymene in Thymus vulgaris L. Comptes Rendus Hebd. Séances L’académie Sci. 1964, 258, 5539–5541. [Google Scholar]
- Balahbib, A.; El Omari, N.; WEl Hachlafi, N.; Lakhdar, F.; El Menyiy, N.; Salhi, N.; Mrabti, H.N.; Bakrim, S.; Zengin, G.; Bouyahya, A. Health beneficial and pharmacological properties of p-cymene. Food Chem. Toxicol. 2021, 153, 112259. [Google Scholar] [CrossRef]
- Olaimat, A.N.; Al-Holy, M.A.; Shahbaz, H.M.; Al-Nabulsi, A.A.; Abu Ghoush, M.H.; Osaili, T.M.; Ayyash, M.M.; Holley, R.A. Emergence of Antibiotic Resistance in Listeria monocytogenes Isolated from Food Products: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1277. [Google Scholar] [CrossRef] [PubMed]
- Wesche, A.M.; Gurtler, J.B.; Marks, B.P.; Ryser, E.T. Stress, sublethal injury, resuscitation, and virulence of bacterial foodborne pathogens. J. Food Prot. 2009, 72, 1121. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; He, M.; Wu, C. Cross protection of lactic acid bacteria during environmental stresses: Stress responses and underlying mechanisms. LWT 2021, 144, 111203. [Google Scholar] [CrossRef]
- Nostro, A.; Marino, A.; Blanco, A.R.; Cellini, L.; Di Giulio, M.; Pizzimenti, F.; Roccaro, A.S.; Bisignano, G. In vitro activity of carvacrol against staphylococcal preformed biofilm by liquid and vapour contact. J. Med. Microbiol. 2009, 58, 791. [Google Scholar] [CrossRef]
- Nostro, A.; Marino, A.; Blanco, A.R.; Cellini, L.; Di Giulio, M.; Pizzimenti, F.; Roccaro, A.S.; Bisignano, G. Evaluation of antimicrobial activity of the hydrolate of Coridothymus capitatus (L.) Reichenb. fil. (Lamiaceae) alone and in combination with antimicrobial agents. BMC Complement. Med. Ther. 2021, 20, 89. [Google Scholar]
- Sharma, S.; Jaiswal, S.; Duffy, B.; Jaiswal, A.K. Advances in emerging technologies for the decontamination of the food contact surfaces. Food Res. Int. 2022, 151, 110865. [Google Scholar] [CrossRef]
- Bezerra dos Santos Rodrigues, J.; de Carvalho, R.J.; de Souza, N.T.; de Sousa Oliveira, K.; Luiz Franco, O.; Schaffner, D.; Leite de Souza, E.; Magnani, M. Effects of oregano essential oil and carvacrol on biofilms of Staphylococcus aureus from food-contact surfaces. Food Control 2017, 73, 1237–1246. [Google Scholar] [CrossRef]
- Buccioni, F.; Purgatorio, C.; Maggio, F.; Garzoli, S.; Rossi, C.; Valbonetti, L.; Paparella, A.; Serio, A. Unraveling the Antimicrobial Effectiveness of Coridothymus capitatus Hydrolate against Listeria monocytogenes in Environmental Conditions Encountered in Foods: An In Vitro Study. Microorganisms 2022, 10, 920. [Google Scholar] [CrossRef] [PubMed]
- Hollander, A.; Yaron, S. Pore-forming treatments induce aggregation of Salmonella Seftenberg through protein leakage. Food Microbiol. 2021, 96, 103721. [Google Scholar] [CrossRef]
- Maggio, F.; Rossi, C.; Chaves-Lòpez, C.; Valbonetti, L.; Desideri, G.; Paparella, A.; Serio, A. A single exposure to a sublethal concentration of Origanum vulgare essential oil initiates response against food stressors and restoration of antibiotic susceptibility in Listeria monocytogenes. Food Control 2022, 132, 108562. [Google Scholar] [CrossRef]
- Stojkovic, D.; Glamoclija, J.; Ciric, A.; Nikolic, M.; Ristic, M.; Siljegovic, J.; Sokovic, M. Investigation on antibacterial synergism of Origanum vulgare and Thymus vulgaris essential oils. Arch. Biol. Sci. 2013, 65, 639–643. [Google Scholar] [CrossRef]
- Pirintsos, S.; Bariotakis, M.; Kampa, M.; Sourvinos, G.; Lionis, C.; Castanas, E. The Therapeutic Potential of the Essential Oil of Thymbra capitata (L.) Cav., Origanum dictamnus L. and Salvia fruticosa Mill. and a Case of Plant-Based Pharmaceutical Development. Front. Pharmacol. 2020, 11, 522213. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Strain | Origin | References |
|---|---|---|
| ATCC 19114 | Type strain | |
| LM1 | Cured pork meat | Unpublished |
| LM2 | Deli meat sandwich | Unpublished |
| LM4 | Deli meat sandwich | [17] |
| LM6 | Pork ribs | [6] |
| LM12 | Cured pork meat | Unpublished |
| LM13 | Pork meat | Unpublished |
| LM17 | Salami | [17] |
| LM19 | Pork meat | [17] |
| L3 | Clinical environments | [18] |
| L253 | ||
| L239 | ||
| L291 | ||
| L315 | ||
| L317 | ||
| L368 |
| Botanical Species | Chemical Component | EO [%] | HY 1 [%] | Origin | Cultivation Method |
|---|---|---|---|---|---|
| Coridothymus capitatus | Carvacrol | 73.0 | 100 | South coast of Sicily | Organic Farming 2 |
| p-Cymene | 9.48 | ||||
| β-Caryophyllene | 5.1 | ||||
| γ-Terpinene | 4.30 | ||||
| Terpinolene | 1.70 | ||||
| β-Thujene | 1.61 | ||||
| β-Myrcene | 1.37 | ||||
| α-Pinene | 1.20 | ||||
| β-Linalool | 0.47 | ||||
| Borneol | 0.4 | ||||
| α-Phellandrene | 0.33 | ||||
| L-Terpinen-4-ol | 0.3 | ||||
| Camphene | 0.26 | ||||
| Thymol | 0.20 | ||||
| β-Pinene | 0.16 | ||||
| p-Mentha-1,3,8-triene | 0.03 | ||||
| Origanum vulgare subsp. hirtum | Thymol | 44.17 | 100 | ||
| γ-Terpinene | 26.09 | ||||
| p-Cymene | 16.03 | ||||
| Terpinolene | 3.66 | ||||
| Isothymol methyl ether | 2.96 | ||||
| β-Thujene | 1.81 | ||||
| β-Myrcene | 1.36 | ||||
| α-Pinene | 0.95 | ||||
| β-Caryophyllene | 0.73 | ||||
| O-Methylthymol | 0.69 | ||||
| Terpinen-4-ol | 0.44 | ||||
| α-Phellandrene | 0.41 | ||||
| δ-Cadinene | 0.18 | ||||
| β-Pinene | 0.13 | ||||
| β-trans-Ocimene | 0.12 | ||||
| Aromadendrene | 0.10 | ||||
| γ-Muurolene | 0.09 | ||||
| Camphene | 0.08 |
| Strain | EOs | HYs | ||
|---|---|---|---|---|
| CEO | OEO | CHY | OHY | |
| ATCC 19114 | 1.25 | 2.5 | 125 | 125 |
| LM1 | 10.0 | 10.0 | 125 | 500 |
| LM2 | 10.0 | 10.0 | 125 | 500 |
| LM4 | 5.0 | 10.0 | n.e. 1 | 250 |
| LM6 | 2.5 | 2.5 | 250 | 250 |
| LM12 | 10.0 | 10.0 | 125 | 500 |
| LM13 | 2.5 | 2.5 | 500 | 500 |
| LM17 | 1.25 | 0.625 | 250 | 125 |
| LM19 | 0.625 | 1.25 | 250 | 250 |
| L315 | 1.25 | 2.5 | 125 | 125 |
| L253 | 2.5 | 0.625 | 250 | 250 |
| L317 | 0.625 | 0.3125 | 250 | 125 |
| L291 | 0.625 | 0.625 | 500 | 250 |
| L239 | 0.3125 | 0.3125 | 250 | 125 |
| L368 | 5.0 | 10.0 | 250 | 125 |
| L3 | 1.25 | 1.25 | 500 | 500 |
| Combination EO/HY | Strain | FIC | FICI | Outcome | |
|---|---|---|---|---|---|
| EO | HY | ||||
| CEO/CHY | ATCC 19114 | 0.500 | 1.000 | 1.500 | Indifferent |
| LM6 | 0.500 | 0.250 | 0.750 | Additive | |
| LM13 | 0.500 | 0.250 | 0.750 | Additive | |
| L315 | 0.500 | 0.500 | 1.000 | Additive | |
| L3 | 0.500 | 0.500 | 1.000 | Additive | |
| OEO/OHY | ATCC 19114 | 0.500 | 0.250 | 0.750 | Additive |
| LM6 | 0.500 | 0.500 | 1.000 | Additive | |
| LM13 | 0.500 | 0.250 | 0.750 | Additive | |
| L315 | 0.500 | 0.062 | 0.562 | Additive | |
| L3 | 0.500 | 0.500 | 1.000 | Additive | |
| CEO/OHY | ATCC 19114 | 0.250 | 0.500 | 0.750 | Additive |
| LM6 | 0.500 | 0.250 | 0.750 | Additive | |
| LM13 | 0.500 | 0.125 | 0.625 | Additive | |
| L315 | 0.500 | 0.500 | 1.000 | Additive | |
| L3 | 0.500 | 0.500 | 1.000 | Additive | |
| OEO/CHY | ATCC 19114 | 0.500 | 0.500 | 1.000 | Additive |
| LM6 | 0.500 | 0.500 | 1.000 | Additive | |
| LM13 | 0.500 | 0.250 | 0.750 | Additive | |
| L315 | 0.500 | 0.007 | 0.507 | Synergistic | |
| L3 | 0.500 | 0.250 | 0.750 | Additive | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Amato, S.; Rossi, C.; Maggio, F.; Valbonetti, L.; Savini, V.; Paparella, A.; Serio, A. Antilisterial Effectiveness of Origanum vulgare var. hirtum and Coridothymus capitatus Essential Oils and Hydrolates Alone and in Combination. Foods 2024, 13, 860. https://0-doi-org.brum.beds.ac.uk/10.3390/foods13060860
D’Amato S, Rossi C, Maggio F, Valbonetti L, Savini V, Paparella A, Serio A. Antilisterial Effectiveness of Origanum vulgare var. hirtum and Coridothymus capitatus Essential Oils and Hydrolates Alone and in Combination. Foods. 2024; 13(6):860. https://0-doi-org.brum.beds.ac.uk/10.3390/foods13060860
Chicago/Turabian StyleD’Amato, Serena, Chiara Rossi, Francesca Maggio, Luca Valbonetti, Vincenzo Savini, Antonello Paparella, and Annalisa Serio. 2024. "Antilisterial Effectiveness of Origanum vulgare var. hirtum and Coridothymus capitatus Essential Oils and Hydrolates Alone and in Combination" Foods 13, no. 6: 860. https://0-doi-org.brum.beds.ac.uk/10.3390/foods13060860