Proteolysis and Rheological Properties of Cream Cheese Made with a Plant-Derived Coagulant from Solanum elaeagnifolium

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Extraction of Proteases from Fruit Powder
2.3. Proteolytic Properties of the Plant-Derived Coagulant
2.4. Capacity of the Plant-Derived Coagulant to Clot Milk
2.5. Manufacture of Cream Cheese
2.6. Evaluation of Proteolysis in Cream Cheeses
2.7. Spreadability of Cream Cheese
2.8. Viscoelastic Properties of Cream Cheese
3. Results and Discussion
3.1. Gross Composition of S. elaeagnifolium Fruits
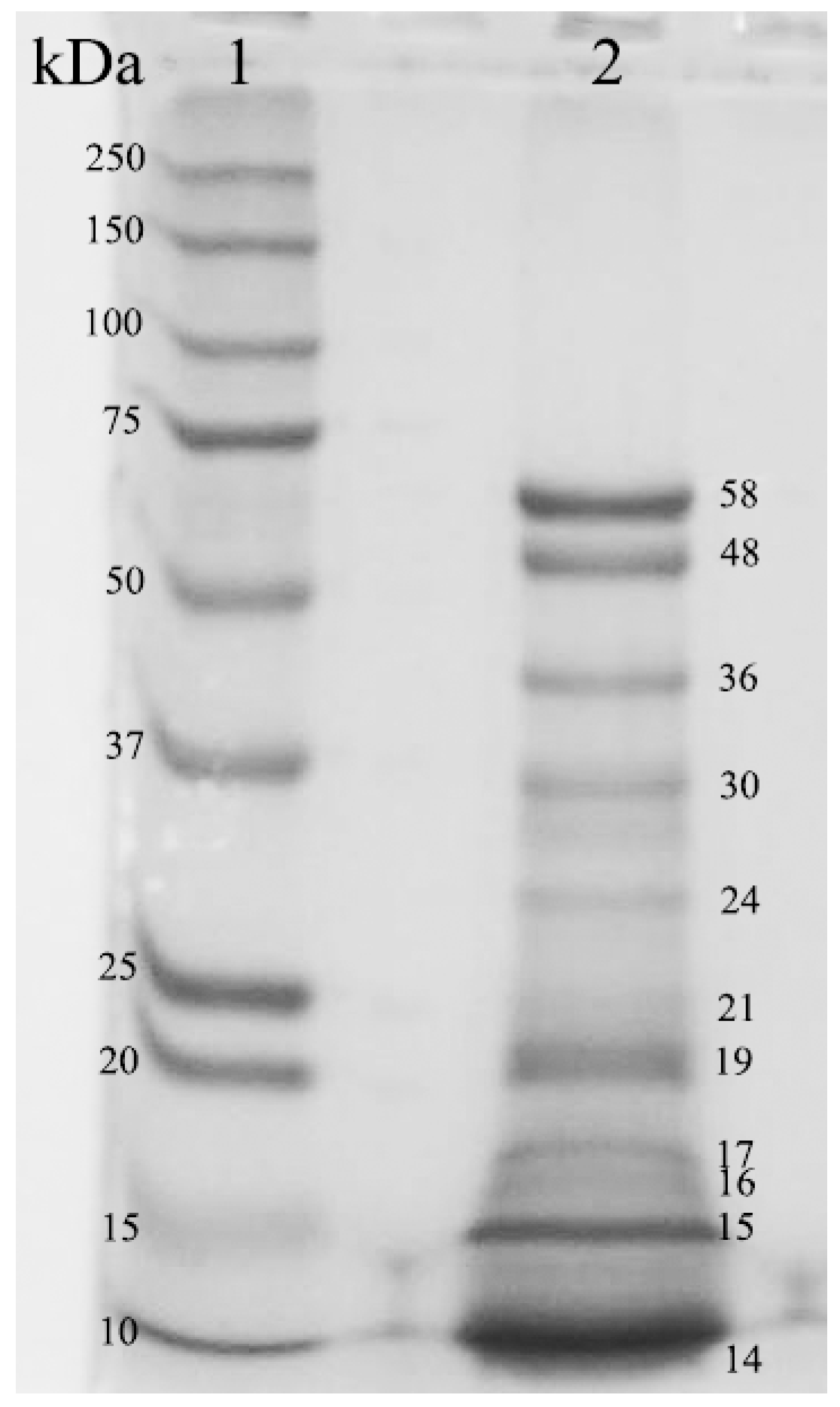
3.2. Proteolytic Properties of the Plant-Derived Coagulant
3.3. Milk Clotting Properties of the Plant-Derived Coagulant
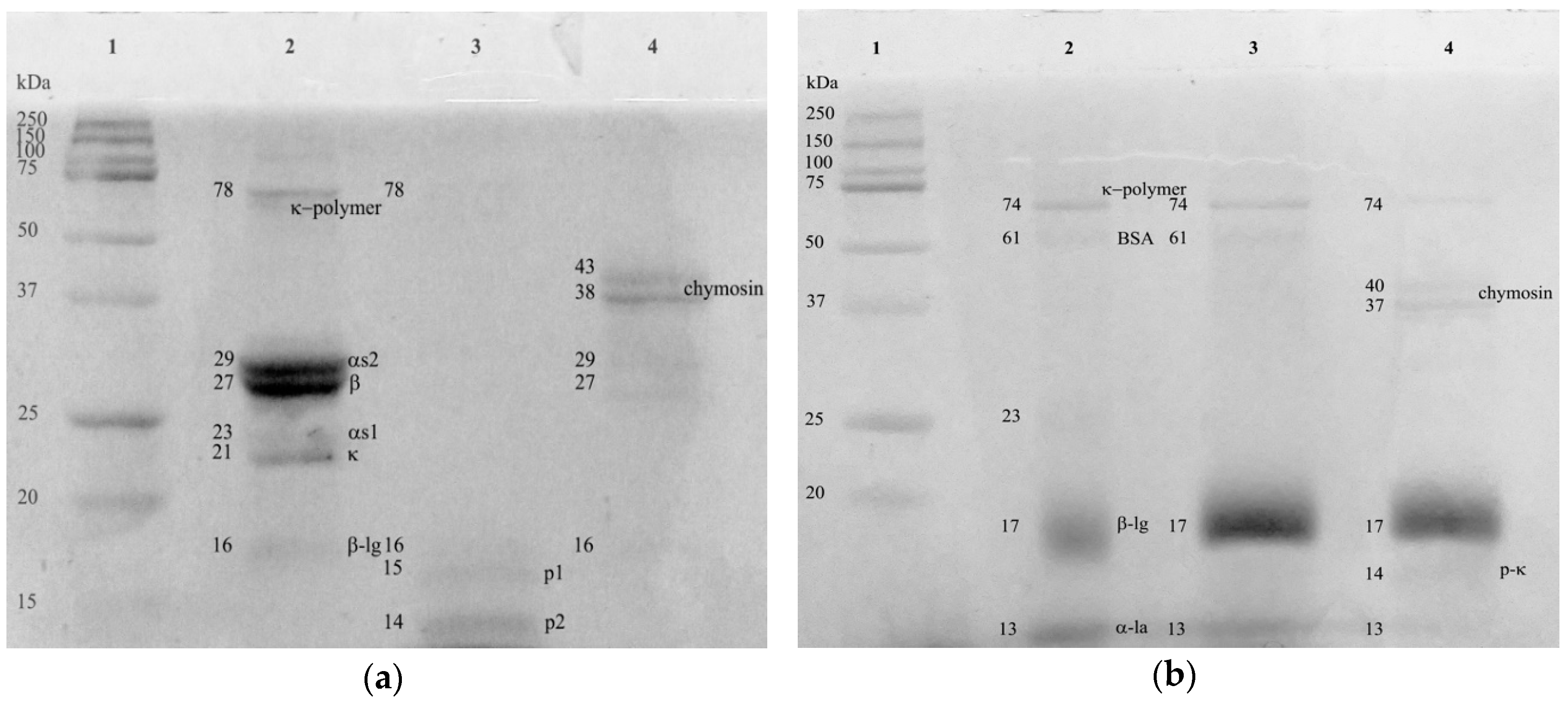
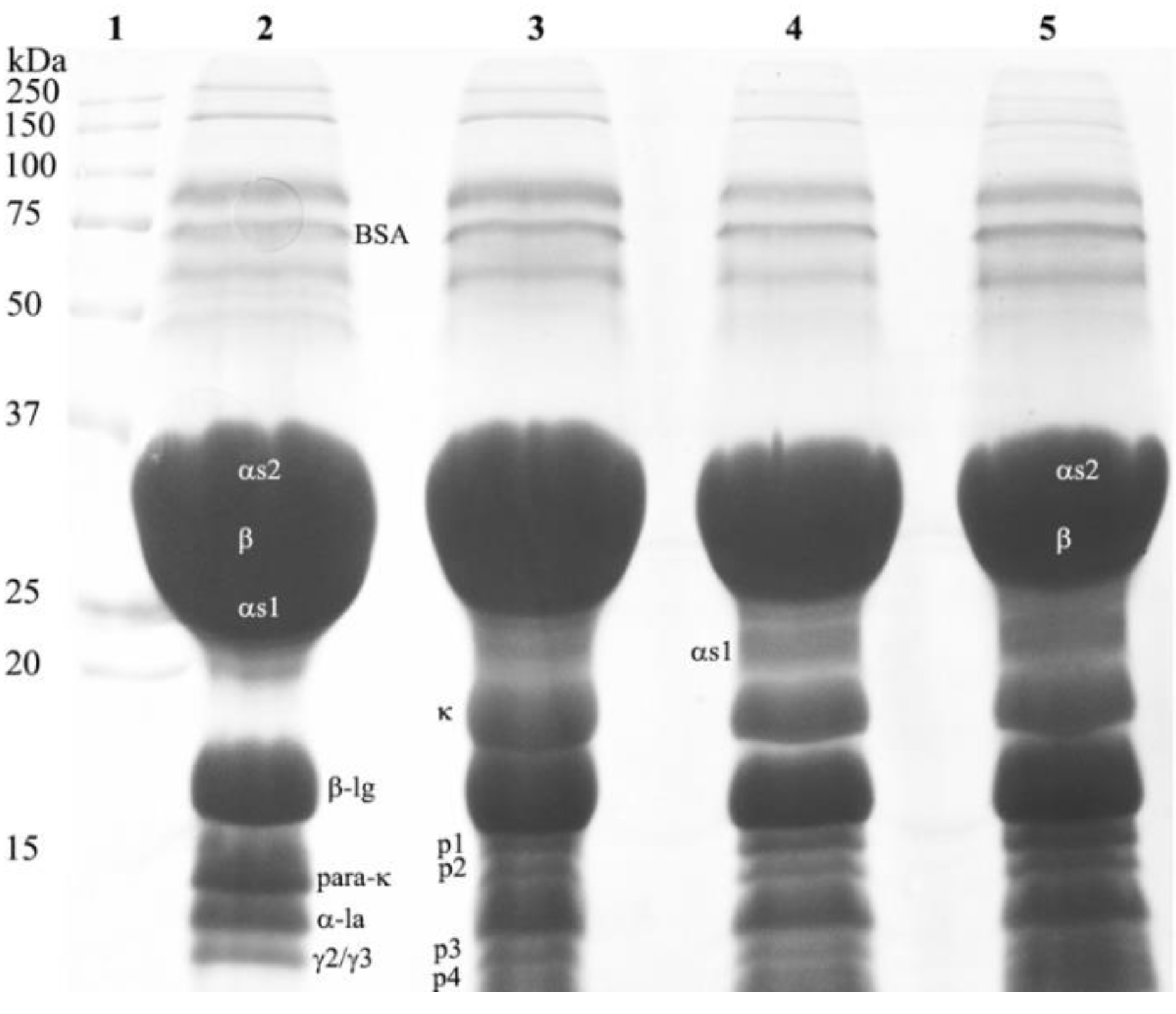
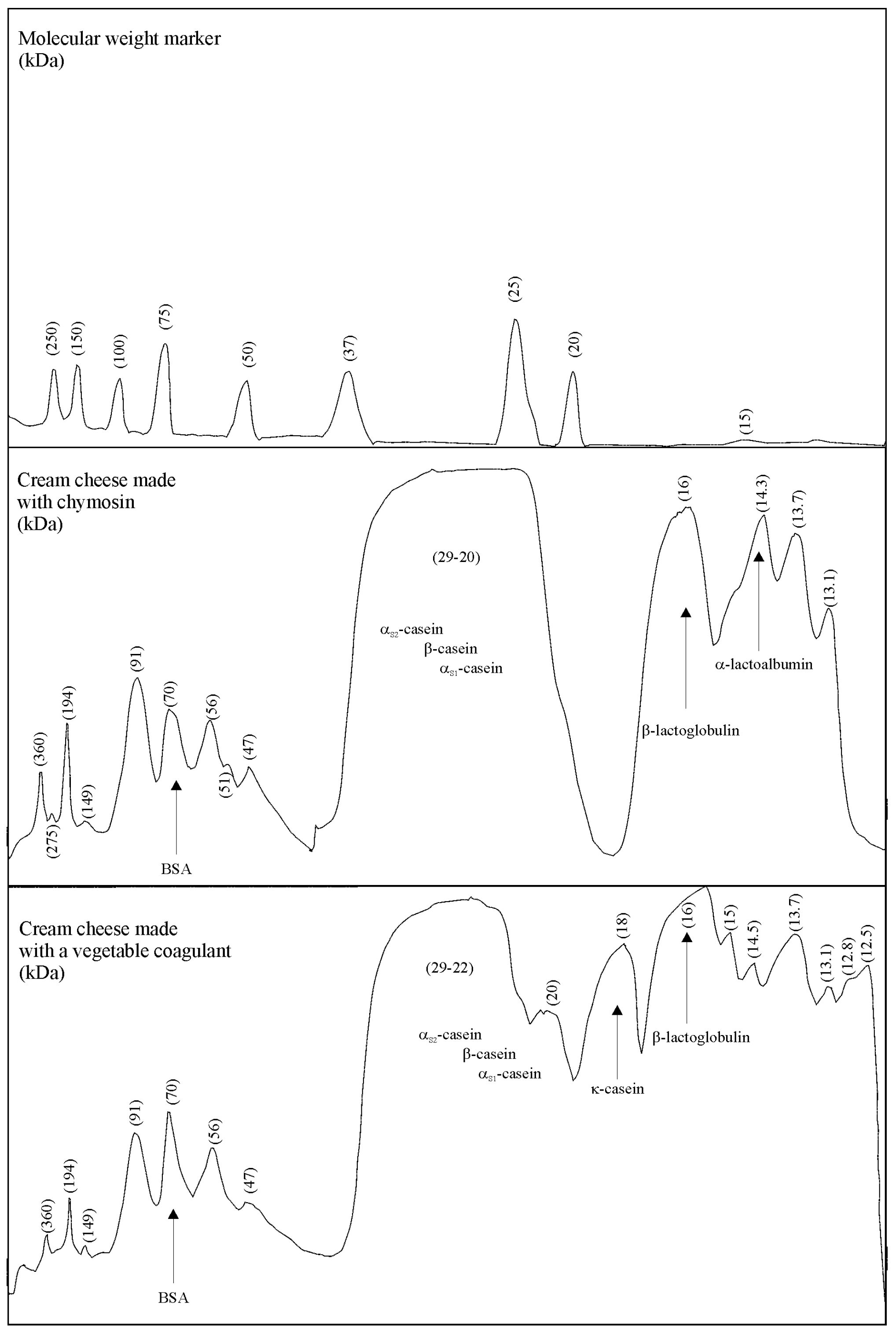
3.4. Composition of Cream Cheese Made with the Plant-Derived Coagulant
3.5. Proteolysis in Cream Cheeses
3.6. Rheological Properties of Cream Cheese
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Brighenti, M.; Govindasamy-Lucey, S.; Lim, K.; Nelson, K.; Lucey, J.A. Characterization of the rheological, textural, and sensory properties of samples of commercial US cream cheese with different fat contents. J. Dairy Sci. 2008, 91, 4501–4517. [Google Scholar] [CrossRef] [PubMed]
- Coutouly, A.; Riaublanc, A.; Axelos, M.; Gaucher, I. Effect of heat treatment, final pH of acidification, and homogenization pressure on the texture properties of cream cheese. Dairy Sci. Technol. 2013, 94, 125–144. [Google Scholar] [CrossRef] [Green Version]
- Schulz-Collins, D.; Senge, B. Acid-and acid/rennet-curd cheeses part A: Quark, cream cheese and related varieties. In Cheese: Chemistry, Physics and Microbiology; Elsevier: New York, NY, USA, 2004; pp. 301–328. [Google Scholar]
- Lucey, J.A.; Singh, H. Formation and physical properties of acid milk gels: A review. Food Res. Int. 1997, 30, 529–542. [Google Scholar] [CrossRef]
- Cosentino, C.; Paolino, R.; Freschi, P.; Calluso, A.M. Short communication: Jenny milk as an inhibitor of late blowing in cheese: A preliminary report. J. Dairy Sci. 2013, 96, 3547–3550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucey, J.; Singh, H. Acid coagulation of milk. In Advanced Dairy Chemistry—1 Proteins; Fox, P.F., McSweeney, P.L.H., Eds.; Springer: New York, NY, USA, 2003; pp. 1001–1025. [Google Scholar]
- Salvatore, E.; Pirisi, A.; Corredig, M. Gelation properties of casein micelles during combined renneting and bacterial fermentation: Effect of concentration by ultrafiltration. Int. Dairy J. 2011, 21, 848–856. [Google Scholar] [CrossRef]
- Jacob, M.; Jaros, D.; Rohm, H. Recent advances in milk clotting enzymes. Int. J. Dairy Technol. 2011, 64, 14–33. [Google Scholar] [CrossRef]
- Shah, M.A.; Mir, S.A.; Paray, M.A. Plant proteases as milk clotting enzymes in cheese-making: A review. Dairy Sci. Technol. 2014, 94, 5–16. [Google Scholar] [CrossRef]
- Ahmed, I.A.M.; Morishina, I.; Babiker, E.E.; Mori, N.; Morishima, I.; Babiker, E.E.; Mori, N. Characterisation of partially purified milk clotting enzyme from Solanum dubium Fresen seeds. Food Chem. 2009, 116, 395–400. [Google Scholar] [CrossRef]
- Chávez-Garay, D.R.; Gutiérrez-Méndez, N.; Valenzuela-Soto, M.E.; García-Triana, A. Partial characterization of a plant coagulant obtained from the berries of Solanum elaeagnifolium. CyTA J. Food 2016, 14, 200–205. [Google Scholar] [CrossRef]
- Gutiérrez-Méndez, N.; Chávez-Garay, D.R.; Jiménez-Campos, H. Exploring the milk clotting properties of a plant coagulant from the berries of S. elaeagnifolium var. Cavanilles. J. Food Sci. 2012, 71, 89–94. [Google Scholar] [CrossRef]
- Bodansky, A. The chymase of Solanum elaeagnifolium a preliminary note. J. Biol. Chem. 1916, 27, 103–105. [Google Scholar]
- Bodansky, A. A study of a milk-coagulating enzyme of Solanum elaeagnifolium. J. Biol. Chem. 1924, 61, 365–375. [Google Scholar]
- AOAC International. Official Methods of Analysis of AOAC International, 16th ed.; Cunnif, P., Ed.; Association of Official Analytical Chemists International: Arlington, MA, USA, 1998. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- McDonald, C.E.; Chen, L.L. The Lowry modification of the Folin reagent for determination of proteinase activity. Anal. Biochem. 1965, 10, 175–177. [Google Scholar] [CrossRef]
- Almanza-Rubio, J.L.; Gutiérrez-Méndez, N.; Leal-Ramos, M.Y.; Sepulveda, D.; Salmeron, I. Modification of the textural and rheological properties of cream cheese using thermosonicated milk. J. Food Eng. 2016, 168, 223–230. [Google Scholar] [CrossRef]
- OEPP. Solanum elaeagnifolium. Bull. OEPP 2007, 37, 236–245. [Google Scholar] [CrossRef]
- Ahmed, I.A.M.; Morishima, I.; Babiker, E.E.; Mori, N. Dubiumin, a chymotrypsin-like serine protease from the seeds of Solanum dubium Fresen. Phytochemistry 2009, 70, 483–491. [Google Scholar] [CrossRef]
- Horne, D.S.; Banks, J.M. Rennet-induced coagulation of milk. In Cheese Chemistry, Physics and Microbiology; Fox, P.F., McSweeney, P.L.H., Cogan, T.M., Guinee, T.P., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2004; pp. 47–70. [Google Scholar]
- Crabbe, M. Rennets: General and molecular aspects. In Cheese: Chemistry, Physics and Microbiology; Elsevier Academic Press: San Diego, CA, USA, 2004; pp. 19–45. [Google Scholar]
- Walstra, P.; Wouters, J.T.M.; Geurst, T.M. Dairy Science and Technology, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Yadav, R.P.; Patel, A.K.; Jagannadham, M.V. Neriifolin S, A dimeric serine protease from Euphorbia Neriifolia Linn.: Purification and biochemical characterisation. Food Chem. 2012, 132, 1296–1304. [Google Scholar] [CrossRef]
- Kumari, M.; Sharma, A.; Jagannadham, M.V. Decolorization of crude latex by activated charcoal, purification and physico-chemical characterization of religiosin, a milk clotting serine protease from the latex of Ficus religiosa. J. Agric. Food Chem. 2010, 58, 8027–8034. [Google Scholar] [CrossRef]
- Martínez-Ruiz, N.R.; López-Díaz, J.A. Optimización de la extracción y estandarización de un cuajo vegetal para la elaboración de queso asadero. Cienc. Front. 2008, 6, 173–176. [Google Scholar]
- Egito, A.S.; Girardet, J.M.; Laguna, L.E.; Poirson, C.; Mollé, D.; Miclo, L.; Humbert, G.; Gaillard, J.L. Milk clotting activity of enzyme extracts from sunflower and albizia seeds and specific hydrolysis of bovine κ-casein. Int. Dairy J. 2007, 17, 816–825. [Google Scholar] [CrossRef]
- Puglisi, I.; Petrone, G.; Lo Piero, A.R. A kiwi juice aqueous solution as coagulant of bovine milk and its potential in Mozzarella cheese manufacture. Food Bioprod. Process. 2014, 92, 67–72. [Google Scholar] [CrossRef]
- Agboola, S.; Chen, S.; Zhao, J. Formation of bitter peptides during ripening of ovine milk cheese made with different coagulants. Dairy Sci. Technol. 2004, 84, 567–578. [Google Scholar] [CrossRef]
- Karametsi, K.; Kokkinidou, S.; Ronningen, I.; Peterson, D.G. Identification of bitter peptides in aged cheddar cheese. J. Agric. Food Chem. 2014, 62, 8034–8041. [Google Scholar] [CrossRef]
- Cosentino, C.; Paolino, R.; Musto, M.; Freschi, P. Innovative use of jenny milk from sustainable rearing. In The Sustainability of Agro-Food and Natural Resource Systems in the Mediterranean Basin; Vastola, A., Ed.; Springer International Publishing: Cham, Switzerland, 2015. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | g/100 g of Dried Fruits |
|---|---|
| Moisture | 5.8 ± 0.1 |
| Fat | 5.0 ± 0.1 |
| Ash | 5.8 ± 0.08 |
| Protein | 13.8 ± 0.3 |
| Total fiber | 56.8 ± 2 |
| Soluble fiber | 2.9 ± 0.9 |
| Insoluble fiber | 53.8 ± 1.1 |
| Carbohydrates 1 | 12.8 |
| (PROTEASE Added) μg of Protein/mL of Milk | (0.022) | (0.4) | (0.8) | (1.6) |
|---|---|---|---|---|
| Component | Control 1 | Cheese made with plant coagulant | ||
| Moisture (g/100 g) | 59.7 ± 8.8 a | 59.5 ± 6.5 a | 58.5 ± 4.4 a | 61.9 ± 2.8 a |
| Fat (g/100 g) | 14.4 ± 0.6 a | 14.4 ± 0.7 a | 14.9 ± 0.1 a | 14.3 ± 0.7 a |
| Protein (g/100 g) | 6.0 ± 0.4 a | 5.1 ± 0.2 ab | 5.3 ± 0.8 ab | 4.7 ± 0.6 b |
| Ash (g/100 g) | 1.1 ± 0.1 a | 1.0 ± 0.05 a | 1.0 ± 0.1 a | 0.8 ± 0.2 a |
| Carbohydrates 2 (g/100 g) | 18.8 ± 2.0 a | 20.1 ± 1.8 a | 20.3 ± 1.9 a | 18.3 ± 2.1 a |
| Yield (%) | 16.1 ± 1.21 a | 14.9 ± 0.74 a | 14.0 ± 2.4 a | 15.0 ± 2.1 a |
| (Protease Added) μg of Protein/mL of Milk | (0.022) | (0.4) | (0.8) | (1.6) |
|---|---|---|---|---|
| Item | Control 1 | Cheese made with plant coagulant | ||
| Spreadability 2 (N) | 151.3 ± 27.8 a | 117.7 ± 8.1 a | 151.6 ± 23.3 a | 101.3 ± 25.8 a |
| η* (kPa×s) | 20.4 ± 7.8 ab | 10.5± 3.9 bc | 29.2 ± 11.2a | 4.4 ± 0.77 c |
| G′ (kPa) | 124.3 ± 28.9 b | 64.1 ± 35.5 bc | 178.6 ± 18.9 a | 26.8 ± 4.8 c |
| G″ (kPa) | 32.1± 13.9 a | 15.8 ± 4.4 b | 42.8 ± 13.3 a | 7.4 ± 2.9 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez-Méndez, N.; Balderrama-Carmona, A.; García-Sandoval, S.E.; Ramírez-Vigil, P.; Leal-Ramos, M.Y.; García-Triana, A. Proteolysis and Rheological Properties of Cream Cheese Made with a Plant-Derived Coagulant from Solanum elaeagnifolium. Foods 2019, 8, 44. https://0-doi-org.brum.beds.ac.uk/10.3390/foods8020044
Gutiérrez-Méndez N, Balderrama-Carmona A, García-Sandoval SE, Ramírez-Vigil P, Leal-Ramos MY, García-Triana A. Proteolysis and Rheological Properties of Cream Cheese Made with a Plant-Derived Coagulant from Solanum elaeagnifolium. Foods. 2019; 8(2):44. https://0-doi-org.brum.beds.ac.uk/10.3390/foods8020044
Chicago/Turabian StyleGutiérrez-Méndez, Néstor, Alejandro Balderrama-Carmona, Socorro E. García-Sandoval, Pamela Ramírez-Vigil, Martha Y. Leal-Ramos, and Antonio García-Triana. 2019. "Proteolysis and Rheological Properties of Cream Cheese Made with a Plant-Derived Coagulant from Solanum elaeagnifolium" Foods 8, no. 2: 44. https://0-doi-org.brum.beds.ac.uk/10.3390/foods8020044