Exploring the Brine Microbiota of a Traditional Norwegian Fermented Fish Product (Rakfisk) from Six Different Producers during Two Consecutive Seasonal Productions

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Production Conditions
2.2. Experimental Design and Sampling
2.3. Chemical Analysis
2.4. Culture-Dependent Microbial Characterization
2.5. Culture-Independent Microbial Characterization
2.5.1. DNA Isolation from Rakfisk Brine
2.5.2. Microbiota Analyses Using a Direct Dideoxy Sequencing Method
2.5.3. MANOVA
2.5.4. Microbiota Analysis Using Illumina Sequencing
2.5.5. Total Number of Bacteria Using Quantitative Real-Time PCR
2.6. 16S rDNA Sequencing of LAB Isolated from MRS Agar
3. Results
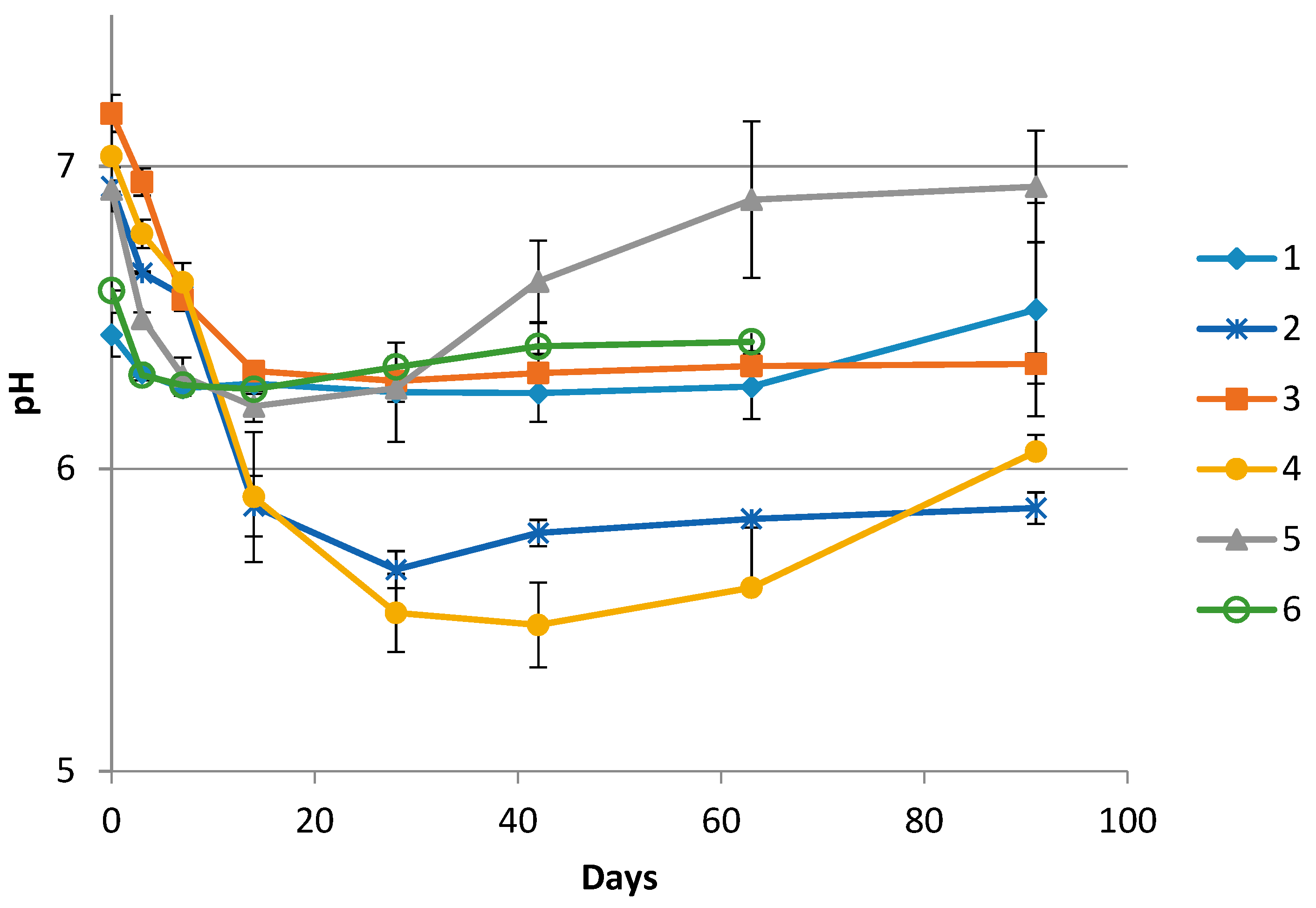
3.1. Chemical Measurements
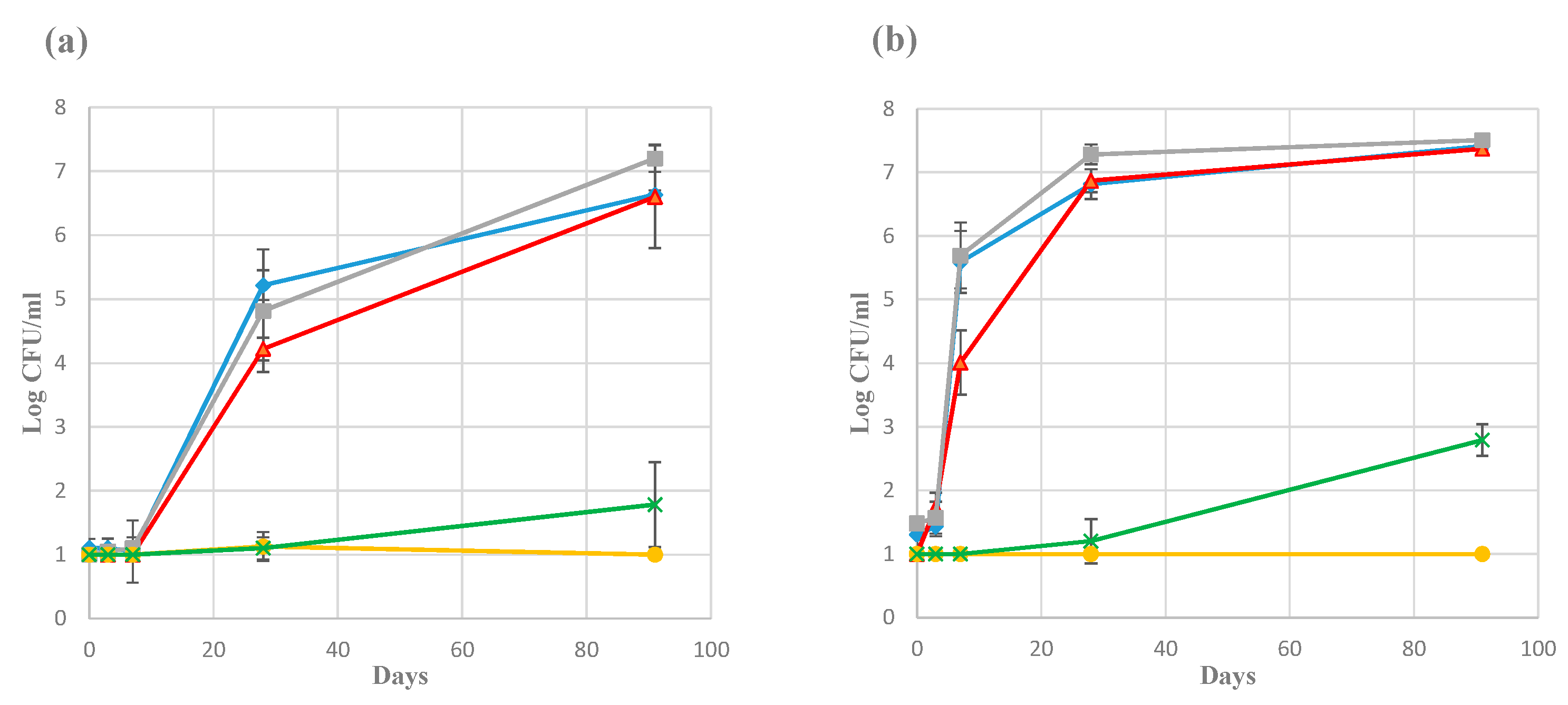
3.2. Plate Counts and Total Number of Bacteria Using qPCR
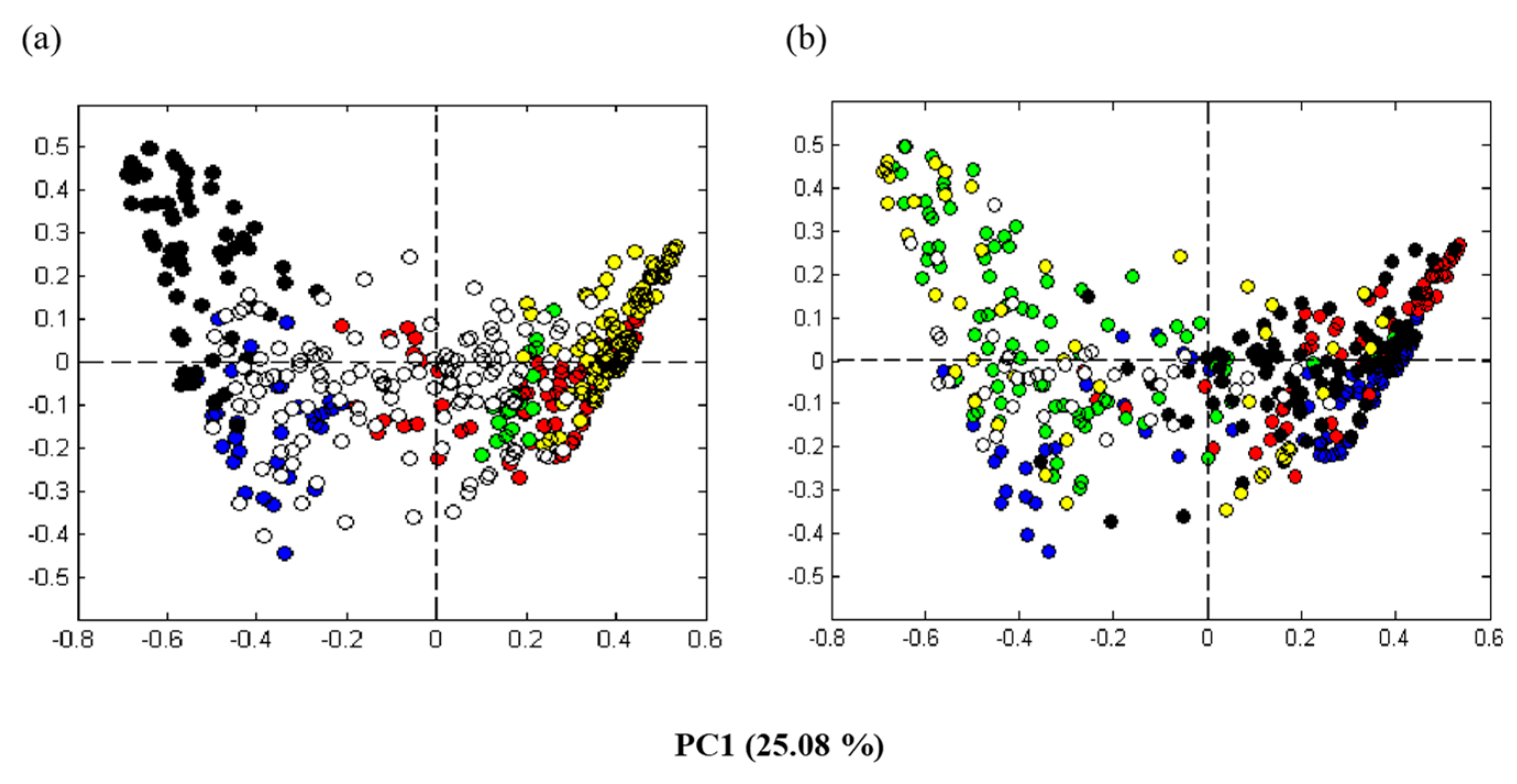
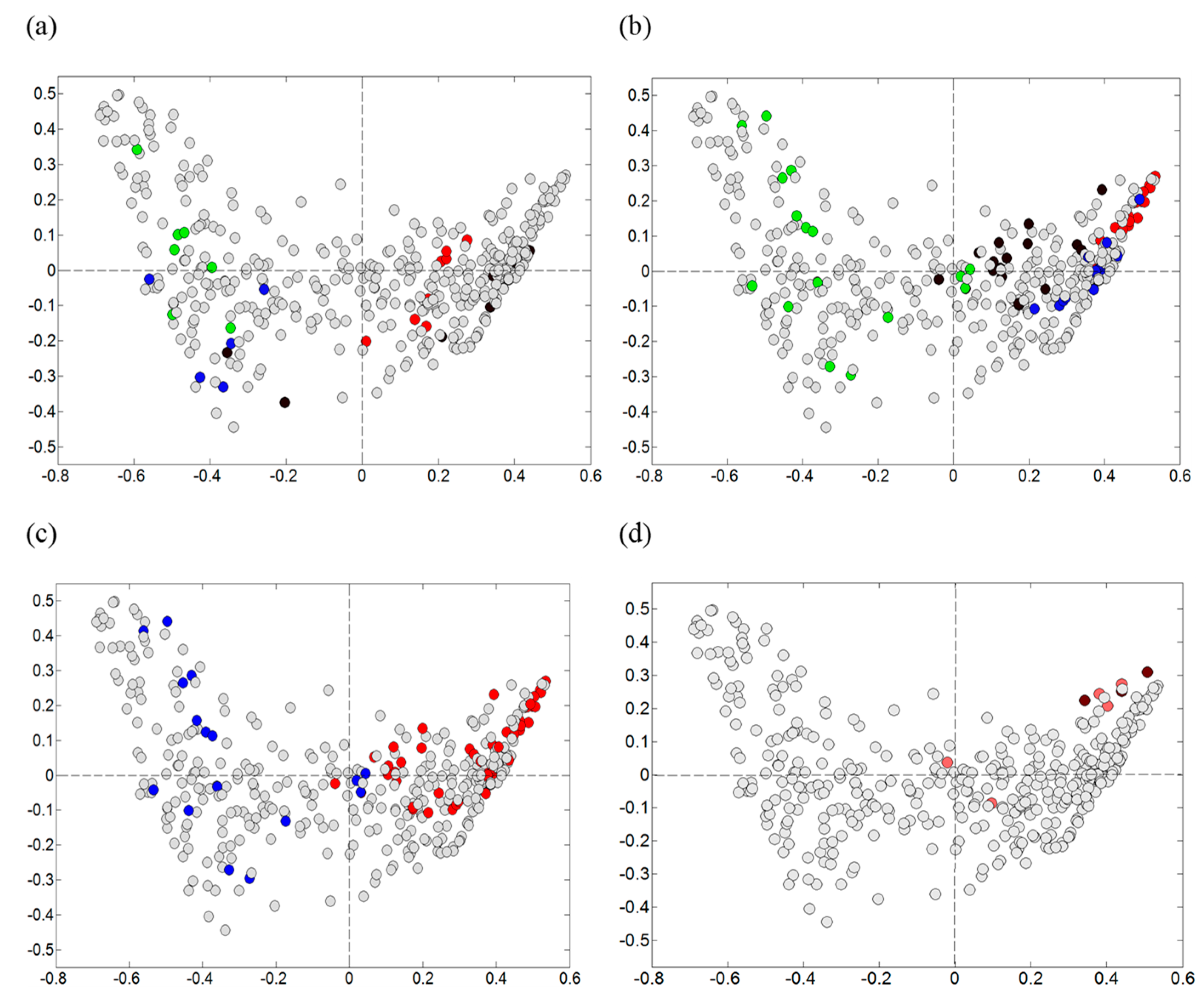
3.3. Microbiota Analysis Using Culture-Independent Characterization
3.3.1. Direct Dideoxy Sequencing
3.3.2. Illumina (MiSeq) Sequencing
3.4. Species Determination of LAB Isolated from MRS Agar
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgements
Conflicts of Interest
References
- Campbell-Platt, G. Fermented foods—A world perspective. Food Res. Int. 1994, 27, 253–257. [Google Scholar] [CrossRef]
- Caplice, E.; Fitzgerald, G.F. Food fermentations: Role of microorganisms in food production and preservation. Int. J. Food Microbiol. 1999, 50, 131–149. [Google Scholar] [CrossRef]
- Steinkraus, K.H. Classification of fermented foods: Worldwide review of household fermentation techniques. Food Control 1997, 8, 311–317. [Google Scholar] [CrossRef]
- Ravyts, F.; De Vuyst, L.; Leroy, F. Bacterial diversity and functionalities in food fermentations. Eng. Life Sci. 2012, 12, 356–367. [Google Scholar] [CrossRef]
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligné, B.; Gänzle, M.; Kort, R.; Pasin, G.; Pihlanto, A.; et al. Health benefits of fermented foods: Microbiota and beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.Y.; Li, Y.; Wu, J.J.; Zhao, Q.L. Diversity of lactic acid bacteria during fermentation of a traditional Chinese fish product, Chouguiyu (stinky mandarinfish). J. Food Sci. 2013, 78, M1778–M1783. [Google Scholar] [CrossRef] [PubMed]
- Fukui, Y.; Yoshida, M.; Shozen, K.; Funatsu, Y.; Takano, T.; Oikawa, H.; Yano, Y.; Satomi, M. Bacterial communities in fish sauce mash using culture-dependent and -independent methods. J. Gen. Appl. Microbiol. 2012, 58, 273–281. [Google Scholar] [CrossRef]
- Kobayashi, T.; Kimura, B.; Fujii, T. Strictly anaerobic halophiles isolated from canned Swedish fermented herrings (Surströmming). Int. J. Food Microbiol. 2000, 54, 81–89. [Google Scholar] [CrossRef]
- Tsuda, H.; Kubota, K.; Matsumoto, T.; Ishimi, Y. Isolation and Identification of lactic acid bacteria in traditional fermented sushi, funazushi, from Japan. Food Sci. Technol. Res. 2012, 18, 77–82. [Google Scholar] [CrossRef]
- Zeng, X.F.; Xia, W.S.; Jiang, Q.X.; Yang, F. Chemical and microbial properties of Chinese traditional low-salt fermented whole fish product Suan yu. Food Control 2013, 30, 590–595. [Google Scholar] [CrossRef]
- Gao, P.; Wang, W.; Jiang, Q.; Xu, Y.; Xia, W. Effect of autochthonous starter cultures on the volatile flavour compounds of Chinese traditional fermented fish (Suan yu). Int. J. Food Sci. Technol. 2016, 51, 1630–1637. [Google Scholar] [CrossRef]
- Skåra, T.; Axelsson, L.; Stefánsson, G.; Ekstrand, B.; Hagen, H. Fermented and ripened fish products in the northern European countries. J. Ethnic Foods 2015, 2, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Axelsson, L. Rakfisk—trygg mat? (“Rakfisk—safe food?”). Toksikologen 2008, 18, 10–13. (In Norwegian) [Google Scholar]
- Schmidt-Nilsen, S.; Bøhmer, T. Zur Kenntnis der wenig gezalzenen Forellen, ‘Rakörret’ (“On the science of the mild-salted trout, ‘rakfisk’”). Kgl. Nor. Vidensk. Selsk. Skr. 1935, 8, 83–86. (In German) [Google Scholar]
- Kim, H.J.; Kim, M.J.; Turner, T.L.; Lee, M.K. Pyrosequencing-based analysis of the bacterial community during fermentation of Alaska pollock sikhae: Traditional Korean seafood. J. Gen. Appl. Microbiol. 2014, 60, 227–233. [Google Scholar]
- Ji, C.; Zhang, J.; Lin, X.; Han, J.; Dong, X.; Yang, S.; Yan, X.; Zhu, B. Metaproteomic analysis of microbiota in the fermented fish, Siniperca chuatsi. LWT Food Sci. Technol. 2017, 80, 479–484. [Google Scholar] [CrossRef]
- Mayo, B.; Rachid, C.T.C.C.; Alegría, A.; Leite, A.M.O.; Peixoto, R.S.; Delgado, S. Impact of next generation sequencing techniques in food microbiology. Curr. Genom. 2014, 15, 293–309. [Google Scholar] [CrossRef] [PubMed]
- Roh, S.W.; Kim, K.H.; Nam, Y.D.; Chang, H.W.; Park, E.J.; Bae, J.W. Investigation of archaeal and bacterial diversity in fermented seafood using barcoded pyrosequencing. ISME J. 2010, 4, 1–16. [Google Scholar] [CrossRef] [PubMed]
- AOAC International. AOAC Official Method 937.09—Salt (Chlorine as Sodium Chloride) in Seafood. Available online: http://www.eoma.aoac.org/methods/info.asp?ID=19783 (accessed on 14 January 2019).
- Frisli, T.; Haverkamp, T.H.A.; Jakobsen, K.S.; Stenseth, N.C.; Rudi, K. Estimation of metagenome size and structure in an experimental soil microbiota from low coverage next-generation sequence data. J. Appl. Microbiol. 2013, 114, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Nadkarni, M.A.; Martin, F.E.; Jacques, N.A.; Hunter, N. Determination of bacterial load by real-time PCR using a broad-range (universal) probe and primers set. Microbiology 2002, 148, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Chaillou, S.; Champomier-Vergès, M.-C.; Cornet, M.; Crutz-Le Coq, A.-M.; Dudez, A.-M.; Martin, V.; Beaufils, S.; Darbon-Rongère, E.; Bossy, R.; Loux, V.; et al. The complete genome sequence of the meat-borne lactic acid bacterium Lactobacillus sakei 23K. Nat. Biotechnol. 2005, 23, 1527–1533. [Google Scholar] [CrossRef] [PubMed]
- Zimonja, M.; Rudi, K.; Trosvik, P.; Næs, T. Multivariate curve resolution of mixed bacterial DNA sequence spectra: Identification and quantification of bacteria in undefined mixture samples. J. Chemom. 2008, 22, 309–322. [Google Scholar] [CrossRef]
- Avershina, E.; Storrø, O.; Oien, T.; Johnsen, R.; Wilson, R.; Egeland, T.; Rudi, K. Bifidobacterial succession and correlation networks in a large unselected cohort of mothers and their children. Appl. Environ. Microbiol. 2013, 79, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Ribosomal Database Project. Available online: http://rdp.cme.msu.edu/ (accessed on 14 January 2019).
- Langsrud, Ø. 50-50 multivariate analysis of variance for collinear responses. J. R. Statist. Soc. D 2002, 51, 305–317. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-Baeza, Y.; Pirrung, M.; Gonzalez, A.; Knight, R. EMPeror: A tool for visualizing high-throughput microbial community data. Gigascience 2013, 2, 16. [Google Scholar] [CrossRef]
- Mao, D.P.; Zhou, Q.; Chen, C.Y.; Quan, Z.X. Coverage evaluation of universal bacterial primers using the metagenomic datasets. BMC Microbiol. 2012, 12, 66. [Google Scholar] [CrossRef]
- van Pijkeren, J.; University of Wisconsin, Madison, WI, USA. Personal communication, 2014.
- McClean, M.N. Colony PCR (Yeast). Available online: https://openwetware.org/wiki/McClean:_Colony_PCR_(Yeast) (accessed on 14 January 2019).
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.L.; McGarrell, D.M.; Sun, Y.N.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef] [PubMed]
- McLeod, A.; Nyquist, O.L.; Snipen, L.; Naterstad, K.; Axelsson, L. Diversity of Lactobacillus sakei strains investigated by phenotypic and genotypic methods. Syst. Appl. Microbiol. 2008, 31, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Hammes, W.P.; Vogel, R.F. The genus Lactobacillus. In The Genera of Lactic Acid Bacteria; Wood, B.J.B., Holzapfel, W.H., Eds.; Chapman & Hall: London, UK, 1995; pp. 19–54. [Google Scholar]
- Torriani, S.; Van Reenen, C.A.; Klein, G.; Reuter, G.; Dellaglio, F.; Dicks, L.M.T. Lactobacillus curvatus subsp. curvatus subsp. nov. and Lactobacillus curvatus subsp. melibiosus subsp. nov. and Lactobacillus sake subsp. sake subsp. nov. and Lactobacillus sake subsp. carnosus subsp. nov., new subspecies of Lactobacillus curvatus Abo-Elnaga and Kandler 1965 and Lactobacillus sake Katagiri, Kitahara, and Fukami 1934 (Klein et al. 1996, emended descriptions), respectively. Int. J. Syst. Bacteriol. 1996, 46, 1158–1163. [Google Scholar] [PubMed]
- Zang, J.; Xu, Y.; Xia, W.; Yu, D.; Gao, P.; Jiang, Q.; Yang, F. Dynamics and diversity of microbial community succession during fermentation of Suan yu, a Chinese traditional fermented fish, determined by high throughput sequencing. Food Res. Int. 2018, 111, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Park, K.Y.; Kim, B.K. Lactic acid bacteria in vegetable fermentations. In Lactic Acid Bacteria, Microbial and Functional Aspects, 4th ed.; Lahtinen, S., Ouwehand, A.C., Salminen, S., Von Wright, A., Eds.; CRC Press: Boca Raton, FL, USA, 2011; pp. 187–211. [Google Scholar]
- Reynisson, E.; Thornor Marteinsson, V.; Jonsdottir, R.; Magnusson, S.H.; Hreggvidsson, G.O. Bacterial succession during curing process of a skate (Dipturus batis) and isolation of novel strains. J. Appl. Microbiol. 2012, 113, 329–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, Y.D.; Yi, S.H.; Lim, S.I. Bacterial diversity of cheonggukjang, a traditional Korean fermented food, analyzed by barcoded pyrosequencing. Food Control 2012, 28, 135–142. [Google Scholar] [CrossRef]
- Humblot, C.; Guyot, J.P. Pyrosequencing of tagged 16S rRNA gene amplicons for rapid deciphering of the microbiomes of fermented foods such as pearl millet slurries. Appl. Environ. Microbiol. 2009, 75, 4354–4361. [Google Scholar] [CrossRef]
- Park, E.J.; Chun, J.; Cha, C.J.; Park, W.S.; Jeon, C.O.; Bae, J.W. Bacterial community analysis during fermentation of ten representative kinds of kimchi with barcoded pyrosequencing. Food Microbiol. 2012, 30, 197–204. [Google Scholar] [CrossRef]
- Lee, S.H.; Jung, J.Y.; Jeon, C.O. Effects of temperature on microbial succession and metabolite change during saeu-jeot fermentation. Food Microbiol. 2014, 38, 16–25. [Google Scholar] [CrossRef]
- Jung, J.Y.; Lee, S.H.; Lee, H.J.; Jeon, C.O. Microbial succession and metabolite changes during fermentation of saeu-jeot: Traditional Korean salted seafood. Food Microbiol. 2013, 34, 360–368. [Google Scholar] [CrossRef]
- Bowman, J. The genus Psychrobacter. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 920–930. [Google Scholar]
- Betts, G. Other spoilage bacteria. In Food Spoilage Microorganisms; Blackburn, C.d.W., Ed.; CRC Press: Boca Raton, FL, USA, 2006; pp. 668–693. [Google Scholar]
- Romanenko, L.A.; Lysenko, A.M.; Rohde, M.; Mikhailov, V.V.; Stackebrandt, E. Psychrobacter maritimus sp. nov. and Psychrobacter arenosus sp. nov., isolated from coastal sea ice and sediments of the Sea of Japan. Int. J. Syst. Evol. Microbiol. 2004, 54, 1741–1745. [Google Scholar] [CrossRef]
- Denner, E.B.M.; Mark, B.; Busse, H.J.; Turkiewicz, M.; Lubitz, W. Psychrobacter proteolyticus sp nov., a psychrotrophic, halotolerant bacterium isolated from the antarctic krill Euphausia superba Dana, excreting a cold-adapted metalloprotease. Syst. Appl. Microbiol. 2001, 24, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Heuchert, A.; Glöckner, F.O.; Amann, R.; Fischer, U. Psychrobacter nivimaris sp. nov., a heterotrophic bacterium attached to organic particles isolated from the South Atlantic (Antarctica). Syst. Appl. Microbiol. 2004, 27, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Bozal, N.; Montes, M.J.; Tudela, E.; Guinea, J. Characterization of several Psychrobacter strains isolated from Antarctic environments and description of Psychrobacter luti sp nov and Psychrobacter fozii sp nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 1093–1100. [Google Scholar] [CrossRef] [PubMed]
- Bjørkevoll, I.; Olsen, R.L.; Skjerdal, O.T. Origin and spoilage potential of the microbiota dominating genus Psychrobacter in sterile rehydrated salt-cured and dried salt-cured cod (Gadus morhua). Int. J. Food Microbiol. 2003, 84, 175–187. [Google Scholar] [CrossRef]
- Song, W.Z.; Lin, X.Z.; Huang, X.H. Characterization and expression analysis of three cold shock protein (CSP) genes under different stress conditions in the Antarctic bacterium Psychrobacter sp. G. Polar Biol. 2012, 35, 1515–1524. [Google Scholar] [CrossRef]
- Marceau, A.; Zagorec, M.; Champomier-Verges, M.C. Positive effects of growth at suboptimal temperature and high salt concentration on long-term survival of Lactobacillus sakei. Res. Microbiol. 2003, 154, 37–42. [Google Scholar] [CrossRef]
- Zheng, B.; Liu, Y.; He, X.; Hu, S.; Li, S.; Chen, M.; Jiang, W. Quality improvement on half-fin anchovy (Setipinna taty) fish sauce by Psychrobacter sp. SP-1 fermentation. J. Sci. Food Agric. 2017, 97, 4484–4493. [Google Scholar] [CrossRef]
- Baati, L.; Fabre-Gea, C.; Auriol, D.; Blanc, P.J. Study of the cryotolerance of Lactobacillus acidophilus: Effect of culture and freezing conditions on the viability and cellular protein levels. Int. J. Food Microbiol. 2000, 59, 241–247. [Google Scholar] [CrossRef]
- Cangelosi, G.A.; Meschke, J.S. Dead or alive: Molecular assessment of microbial viability. Appl. Environ. Microbiol. 2014, 80, 5884–5891. [Google Scholar] [CrossRef]
- Fogel, G.B.; Collins, C.R.; Li, J.; Brunk, C.F. Prokaryotic genome size and SSU rDNA copy number: Estimation of microbial relative abundance from a mixed population. Microb. Ecol. 1999, 38, 93–113. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Saito, R.; Miya, S.; Tanaka, Y.; Miyamura, N.; Kuda, T.; Kimura, B. Development of quantitative real-time PCR for detection and enumeration of Enterobacteriaceae. Int. J. Food Microbiol. 2017, 246, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Fouhy, F.; Deane, J.; Rea, M.C.; O’Sullivan, Ó.; Ross, R.P.; O’Callaghan, G.; Plant, B.J.; Stanton, C. The effects of freezing on faecal microbiota as determined using MiSeq sequencing and culture-based investigations. PLoS ONE 2015, 10, e0119355. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
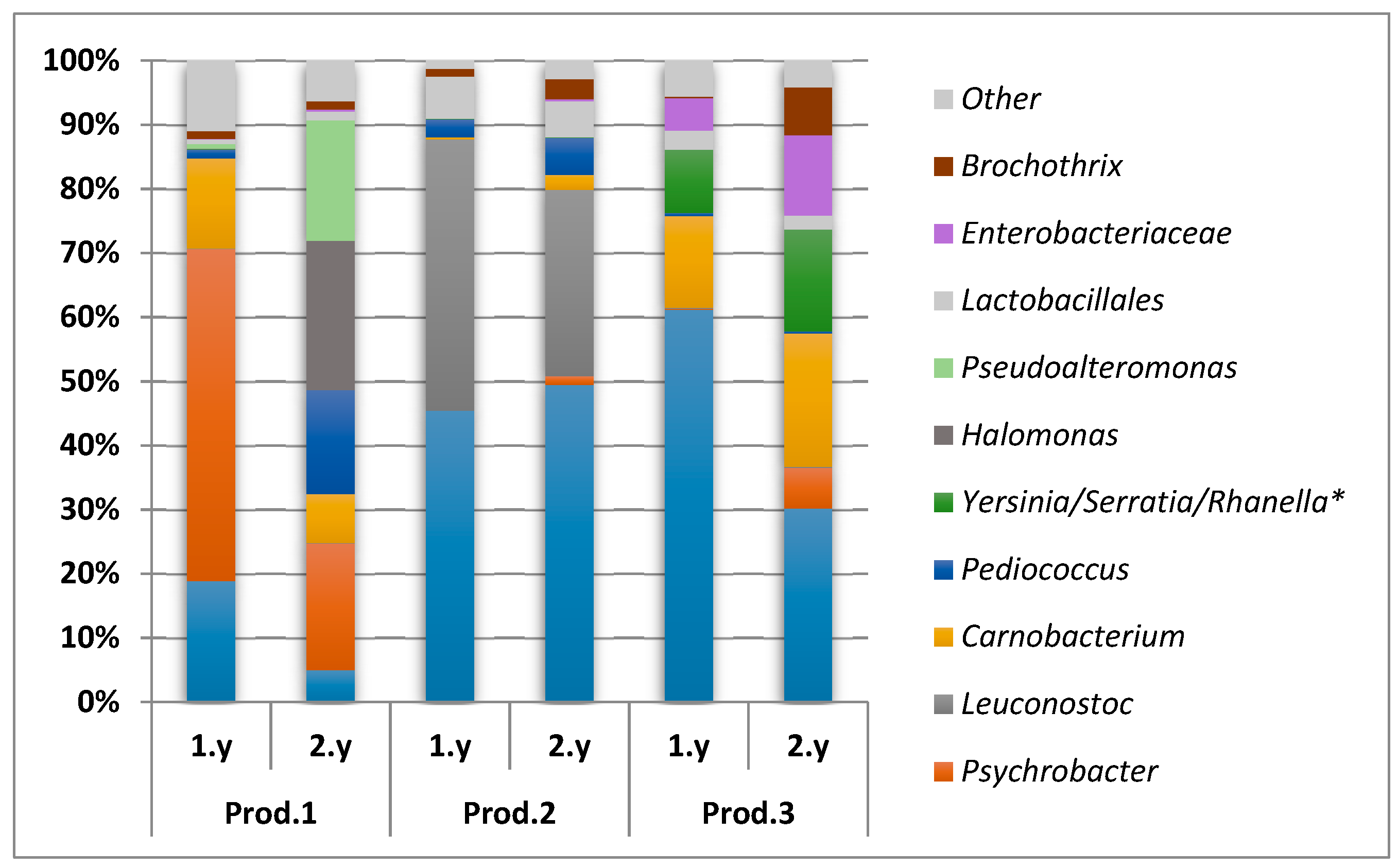{kind=link}
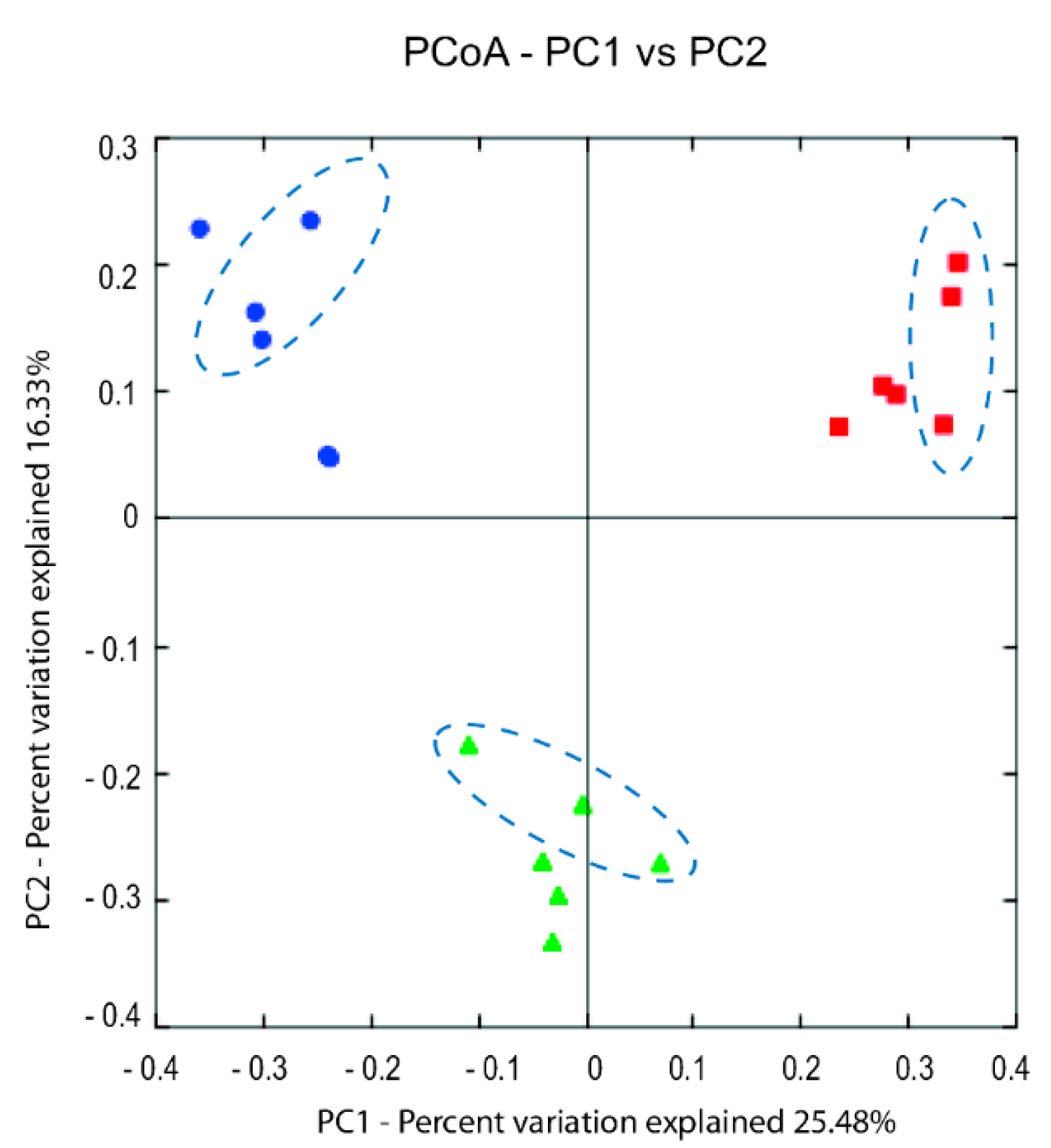{kind=link}
| Producer ID | Fish Species/Origin b | Temperature °C | NaCl % (w/v) d | Sugar Addition | Starter Culture Addition g | Salting Method i | Sampling k |
|---|---|---|---|---|---|---|---|
| 1 a | Char/A | 3.5/4.5 c | 5.8/5.7 | No | No | Dry | Complete |
| 2 | Char/B | 5.0 | 3.5/4.0 | Yes e | No h | Brine | Complete |
| 3 | Trout/C | 7.0 | 4.9/4.2 | No | No | Dry | Complete |
| 4 | Trout/C | 7.0 | 4.8/5.1 | Yes f | Yes | Dry | Year 2+ |
| 5 | Trout/D | 7.5 | 4.9/3.9 | No | No | Dry | Year 2+ |
| 6 | Trout/E | 6.0 | 6.9/7.4 | No | No | Dry | Year 2+ m |
| Category | Explained Variance (%) | p-Value |
|---|---|---|
| Producer a | 45.43 | <0.0001 |
| Year b | 6.98 | 0.0001 |
| Producer and year c | 16.99 | <0.0001 |
| Error | 27.42 |
| Producers | Psychrobacter | Lactobacillus | ||||
|---|---|---|---|---|---|---|
| 0–7 d | 63–91 d | p-Value | 0–7 d | 63–91 d | p-Value | |
| 1 | 30.7 ± 20.6 | 50.5 ± 31.5 | 0.2 | 5.4 ± 2.9 | 5.7 ± 8.6 | 0.9 |
| 2 | 27.1 ± 18.3 | 0.5 ± 0.9 | 0.02 | 0.1 ± 0.2 | 80.5 ± 9.10 | <0.00001 a |
| 3 | 2.7 ± 5.3 | 7. 6 ± 6.8 | 0.1 | 16.5 ± 11.1 | 38.3 ± 24.3 | 0.04 a |
| 4 | 5.9 ± 3.2 | 1.2 ± 1.0 | 0.02 | 12.4 ± 7.5 | 79.6 ± 11.2 | <0.000001 a |
| 5 | 9.2 ± 18.5 | 25.8 ± 20.7 | 0.2 | 18.7 ± 11.1 | 23.4 ± 34.7 | 0.8 |
| 6 | 25.5 ± 16.8 | 41.6 ± 14.2 | 0.09 | 10.6 ± 13.6 | 7.7 ± 9.1 | 0.7 |
| 1 b | ND | 10.0 ± 4.1 | 0.05 | ND | 72.1 ± 17.2 | <0.00001 a,c |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bjerke, G.A.; Rudi, K.; Avershina, E.; Moen, B.; Blom, H.; Axelsson, L. Exploring the Brine Microbiota of a Traditional Norwegian Fermented Fish Product (Rakfisk) from Six Different Producers during Two Consecutive Seasonal Productions. Foods 2019, 8, 72. https://0-doi-org.brum.beds.ac.uk/10.3390/foods8020072
Bjerke GA, Rudi K, Avershina E, Moen B, Blom H, Axelsson L. Exploring the Brine Microbiota of a Traditional Norwegian Fermented Fish Product (Rakfisk) from Six Different Producers during Two Consecutive Seasonal Productions. Foods. 2019; 8(2):72. https://0-doi-org.brum.beds.ac.uk/10.3390/foods8020072
Chicago/Turabian StyleBjerke, Guro Alette, Knut Rudi, Ekaterina Avershina, Birgitte Moen, Hans Blom, and Lars Axelsson. 2019. "Exploring the Brine Microbiota of a Traditional Norwegian Fermented Fish Product (Rakfisk) from Six Different Producers during Two Consecutive Seasonal Productions" Foods 8, no. 2: 72. https://0-doi-org.brum.beds.ac.uk/10.3390/foods8020072