Improving Polyphenolic Compounds: Antioxidant Activity in Chickpea Sprouts through Elicitation with Hydrogen Peroxide

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Chickpea Germination Process
2.3. Germination Percentage
2.4. Preparation of Phytochemical Extracts
2.5. Determination of Total Phenolic Content and Total Flavonoids Content
2.6. Antioxidant Activity
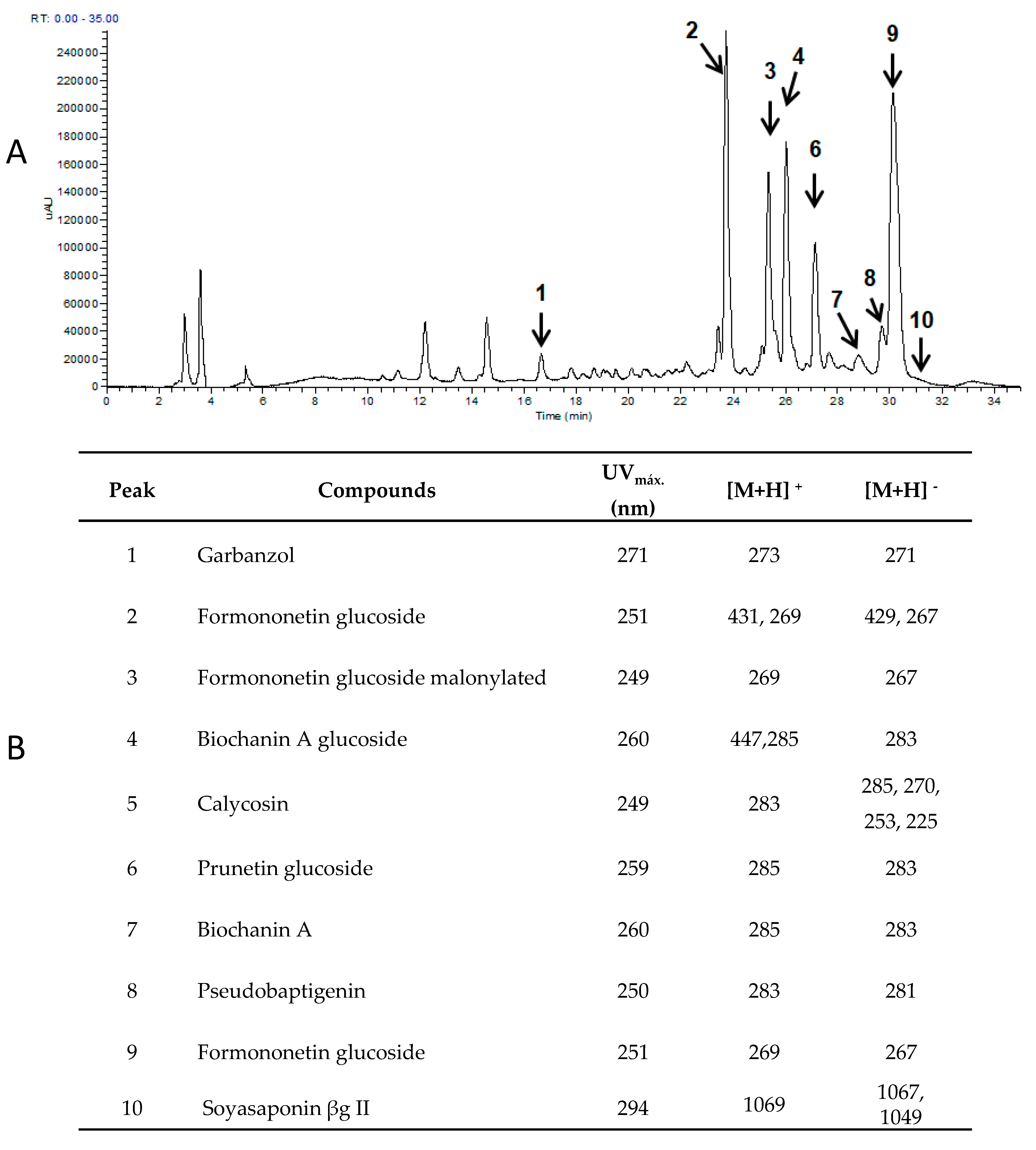
2.7. Identification and Quantification of Flavonoids
2.8. Chemical Composition and Antinutritional Components
2.8.1. Chemical Composition
2.8.2. Phytic Acid
2.8.3. Saponin Content
2.8.4. Trypsin Inhibitors Activity
2.9. Regression Analysis and Optimization
2.10. Statistical Analysis
3. Results and Discussion
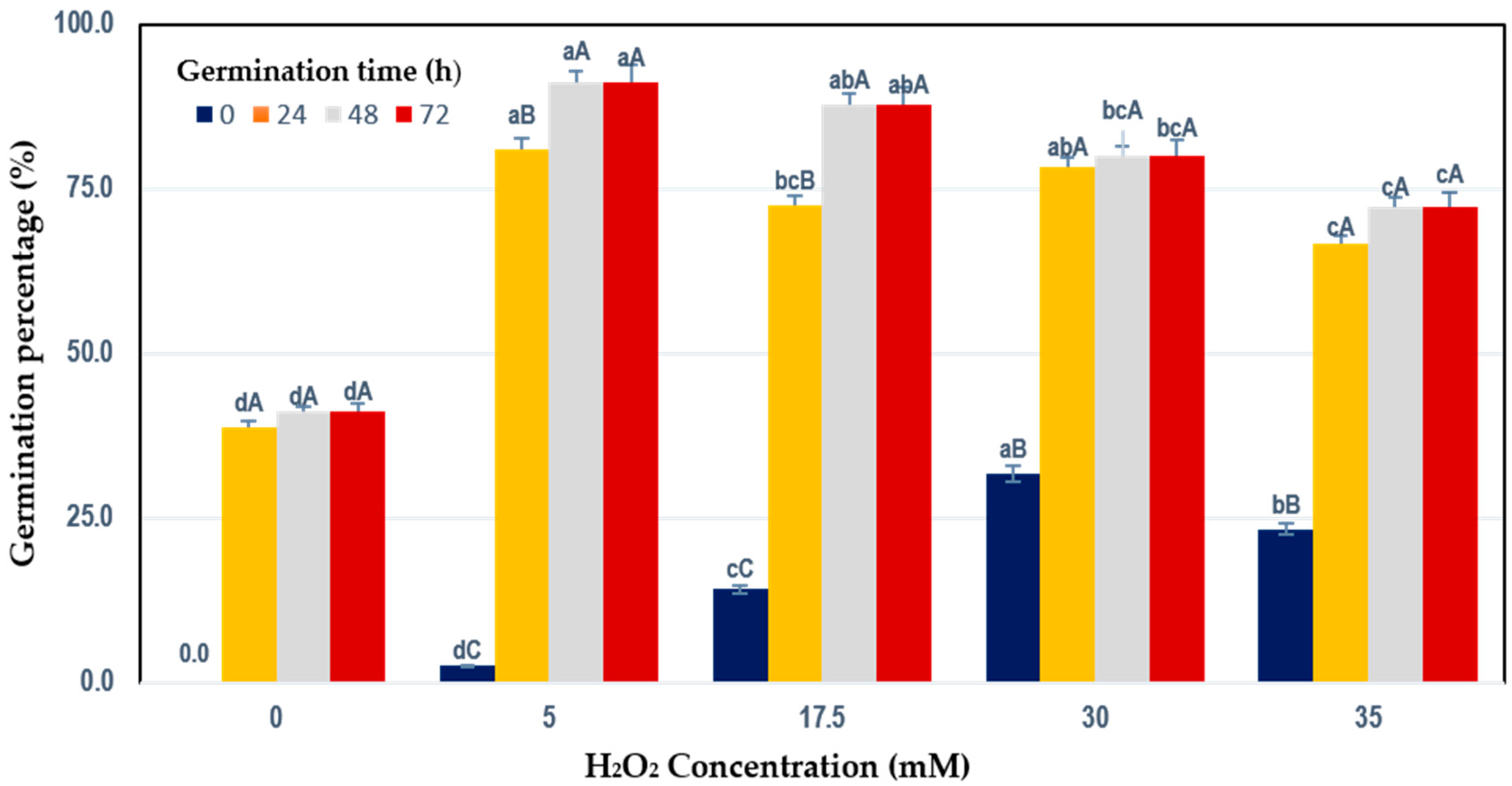
3.1. Percentage of Germination
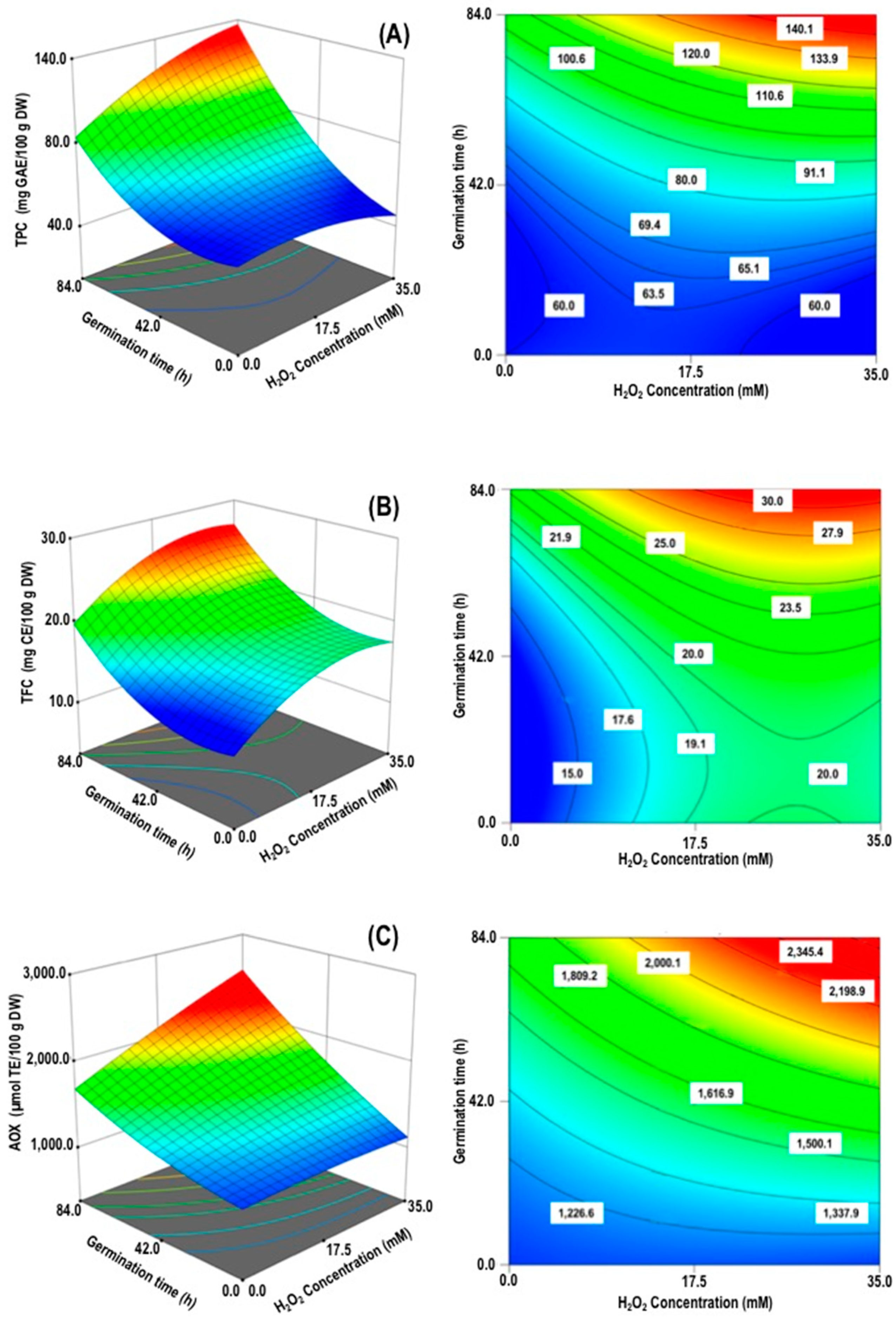
3.2. Appropriate Models by Response Variables
3.2.1. Total Phenolics Content (TPC)
3.2.2. Total Flavonoids Content (TFC)
3.2.3. Antioxidant Activity (AOX)
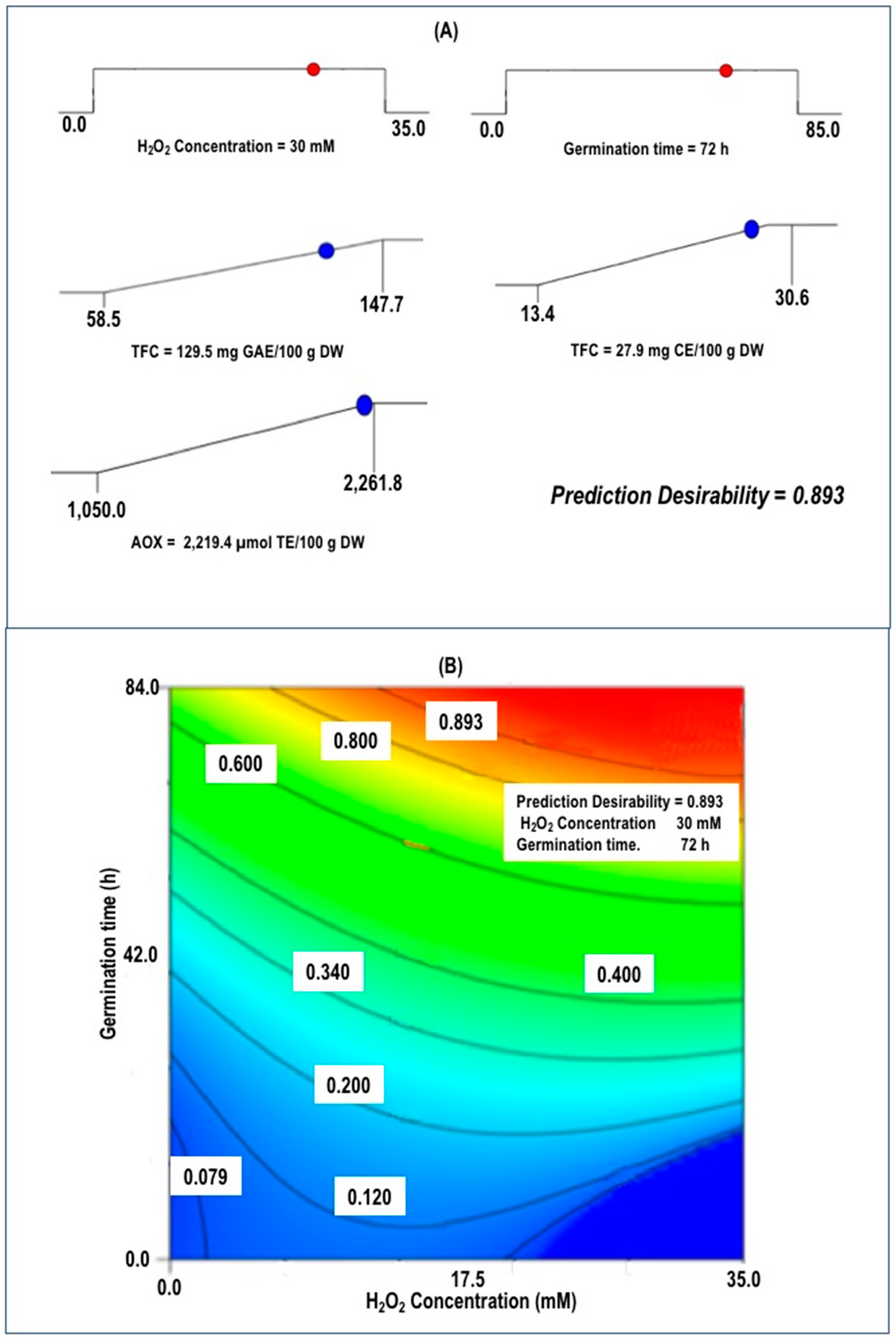
3.3. Optimization and Validation of Germination Conditions
3.4. Chemical Composition, Antinutritional and Nutraceutical Properties of Optimized Germinated Sprouts
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Heiras-Palazuelos, M.J.; Ochoa-Lugo, M.I.; Gutiérrez-Dorado, R.; Lopez-Valenzuela, J.A.; Mora-Rochín, S.; Milán-Carrillo, J.; Garzón-Tiznado, J.A.; Reyes-Moreno, C. Technological properties, antioxidant activity and total phenolic and flavonoid content of pigmented chickpea (Cicer arietinum L.) cultivars. Int. J. Food Sci. Nutr. 2012, 64, 69–76. [Google Scholar] [CrossRef]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic composition and antioxidant potential of grain legume seeds: A review. Food Res. Int. 2017, 101, 1–16. [Google Scholar] [CrossRef]
- Guardado-Félix, D.; Serna-Saldívar, S.O.; Cuevas-Rodríguez, E.O.; Jacobo-Velázquez, D.A.; Gutiérrez-Uribe, J.A. Effect of sodium selenite on isoflavonoid contents and antioxidant capacity of chickpea (Cicer arietinum L.) sprouts. Food Chem. 2017, 226, 69–74. [Google Scholar] [CrossRef]
- Milán-Noris, A.K.; Gutiérrez-Uribe, J.A.; Santacruz, A.; Serna-Saldívar, S.O.; Martínez-Villaluenga, C. Peptides and isoflavones in gastrointestinal digests contribute to the anti-inflammatory potential of cooked or germinated desi and kabuli chickpea (Cicer arietinum L.). Food Chem. 2018, 268, 66–76. [Google Scholar] [CrossRef]
- Du, H.; Huang, Y.; Tang, Y. Genetic and metabolic engineering of isoflavonoid biosynthesis. Appl. Microbiol. Biotechnol. 2010, 86, 1293–1312. [Google Scholar] [CrossRef]
- Gao, Y.; Yao, Y.; Zhu, Y.; Ren, G. Isoflavone Content and Composition in Chickpea (Cicer arietinum L.) Sprouts Germinated under Different Conditions. J. Agric. Food Chem. 2015, 63, 2701–2707. [Google Scholar] [CrossRef]
- Hübner, F.; Arendt, E.K. Germination of Cereal Grains as a Way to Improve the Nutritional Value: A Review. Crit. Rev. Food Sci. Nutr. 2013, 53, 853–861. [Google Scholar] [CrossRef]
- Gan, R.-Y.; Lui, W.-Y.; Wu, K.; Chan, C.-L.; Dai, S.-H.; Sui, Z.-Q.; Corke, H. Bioactive compounds and bioactivities of germinated edible seeds and sprouts: An updated review. Trends Food Sci. Technol. 2017, 59, 1–14. [Google Scholar] [CrossRef]
- Cáceres, P.J.; Martínez-Villaluenga, C.; Amigo, L.; Frias, J. Maximising the phytochemical content and antioxidant activity of Ecuadorian brown rice sprouts through optimal germination conditions. Food Chem. 2014, 152, 407–414. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Rao, J.; Chen, B. Phenolic compounds in germinated cereal and pulse seeds: Classification, transformation, and metabolic process. Crit. Rev. Food Sci. Nutr. 2019, 60, 740–759. [Google Scholar] [CrossRef]
- Sahai, V.; Kumar, V. Anti-diabetic, hepatoprotective and antioxidant potential of Brassica oleracea sprouts. Biocatal. Agric. Biotechnol. 2020, 25, 101623. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Aguilera, Y.; Herrera, T.; Cayuelas, L.T.; Dueñas, M.; Rodríguez-Rodríguez, P.; Ramiro-Cortijo, D.; Arribas, S.M.; Martín-Cabrejas, M.A. Bioavailability of Melatonin from Lentil Sprouts and Its Role in the Plasmatic Antioxidant Status in Rats. Foods 2020, 9, 330. [Google Scholar] [CrossRef] [Green Version]
- Limón, R.I.; Peñas, E.; Martínez-Villaluenga, C.; Frias, J. Role of elicitation on the health-promoting properties of kidney bean sprouts. LWT 2014, 56, 328–334. [Google Scholar] [CrossRef] [Green Version]
- Świeca, M. Production of ready-to-eat lentil sprouts with improved antioxidant capacity: Optimization of elicitation conditions with hydrogen peroxide. Food Chem. 2015, 180, 219–226. [Google Scholar] [CrossRef]
- Barba-Espín, G.; Hernández, J.A.; Diaz-Vivancos, P. Role of H2O2 in pea seed germination. Plant Signal. Behav. 2012, 7, 193–195. [Google Scholar] [CrossRef] [Green Version]
- Baenas, N.; García-Viguera, C.; Moreno, D.A. Elicitation: A Tool for Enriching the Bioactive Composition of Foods. Molecules 2014, 19, 13541–13563. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Yadav, S.; Sibi, G. Seed germination and maturation under the influence of hydrogen peroxide—A review. J. Crit. Rev. 2020, 7, 6–10. [Google Scholar] [CrossRef]
- Mendoza-Sánchez, M.; Guevara-González, R.G.; Castaño-Tostado, E.; Mercado-Silva, E.M.; Acosta-Gallegos, J.A.; Rocha-Guzmán, N.E.; Reynoso-Camacho, R. Effect of chemical stress on germination of cv Dalia bean (Phaseolus vularis L.) as an alternative to increase antioxidant and nutraceutical compounds in sprouts. Food Chem. 2016, 212, 128–137. [Google Scholar] [CrossRef]
- Świeca, M.; Surdyka, M.; Gawlik-Dziki, U.; Złotek, U.; Baraniak, B. Antioxidant potential of fresh and stored lentil sprouts affected by elicitation with temperature stresses. Int. J. Food Sci. Technol. 2014, 49, 1811–1817. [Google Scholar] [CrossRef]
- Peñas, E.; Limón, R.I.; Martínez-Villaluenga, C.; Restani, P.; Pihlanto, A.; Frias, J. Impact of Elicitation on Antioxidant and Potential Antihypertensive Properties of Lentil Sprouts. Plant Foods Hum. Nutr. 2015, 70, 401–407. [Google Scholar] [CrossRef] [Green Version]
- Dueñas, M.; Emartinez-Villaluenga, C.; Limón, R.I.; Peñas, E.; Frias, J. Effect of germination and elicitation on phenolic composition and bioactivity of kidney beans. Food Res. Int. 2015, 70, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Chen, Y.; Hu, T.; Zhang, S.; Zhang, Y.; Zhao, T.; Yu, H.; Kang, Y. The influence of light-emitting diodes on the phenolic compounds and antioxidant activities in pea sprouts. J. Funct. Foods 2016, 25, 459–465. [Google Scholar] [CrossRef]
- Zhang, L.; Mao, X.; Xia, Z. Effects of Sodium Metavanadate and Germination on the Sprouting of Chickpeas and its Content of Vanadium, Formononetin and Biochanin A in the Sprouts. J. Diet. Suppl. 2012, 9, 34–44. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Heimler, D.; Vignolini, P.; Dini, M.G.; Vincieri, F.F.; Romani, A. Antiradical activity and polyphenol composition of local Brassicaceae edible varieties. Food Chem. 2006, 99, 464–469. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free. Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- AOAC. Official Method of Analysis, 18th ed.; Association of Officiating Analytical Chemists: Washington, DC, USA, 2005. [Google Scholar]
- Gao, Y.; Shang, C.; Maroof, M.A.S.; Biyashev, R.M.; Grabau, E.A.; Kwanyuen, P.; Burton, J.W.; Buss, G.R. A Modified Colorimetric Method for Phytic Acid Analysis in Soybean. Crop Sci. 2007, 47, 1797–1803. [Google Scholar] [CrossRef] [Green Version]
- Dini, I.; Tenore, G.C.; Dini, A. Saponins in Ipomoea batatas tubers: Isolation, characterization, quantification and antioxidant properties. Food Chem. 2009, 113, 411–419. [Google Scholar] [CrossRef]
- Domoney, C.; Welham, T. Trypsin inhibitors in Pisum: Variation in amount and pattern of accumulation in developing seed. Seed Sci. Res. 1992, 2, 147–154. [Google Scholar] [CrossRef]
- Khuri, A.A.; Cornell, J.A. Response Surfaces: Designs and Analyses; Marcel Dekker: New York, NY, USA, 1987; Volume 254, pp. 1–17. [Google Scholar]
- Derringer, G.; Suich, R. Simultaneous Optimization of Several Response Variables. J. Qual. Technol. 1980, 12, 214–219. [Google Scholar] [CrossRef]
- Ogawa, K.; Iwabuchi, M. A Mechanism for Promoting the Germination of Zinnia elegans Seeds by Hydrogen Peroxide. Plant Cell Physiol. 2001, 42, 286–291. [Google Scholar] [CrossRef] [Green Version]
- Barba-Espin, G.; Diaz-Vivancos, P.; Clemente-Moreno, M.J.; Albacete, A.; Faize, L.; Faize, M.; Pérez-Alfocea, F.; Hernández, J.A. Interaction between hydrogen peroxide and plant hormones during germination and the early growth of pea seedlings. Plant Cell Environ. 2010, 33, 981–994. [Google Scholar] [CrossRef]
- Sohail, S.A.; Chaurasia, A.; Bara, B.M. Effect of Different Seed Priming Methods on Germination and Vigour of Kabuli Chickpea (Cicer kabulium L.) Seeds. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 1396–1404. [Google Scholar] [CrossRef]
- Candioti, L.V.; De Zan, M.M.; Cámara, M.S.; Goicoechea, H.C. Experimental design and multiple response optimization. Using the desirability function in analytical methods development. Talanta 2014, 124, 123–138. [Google Scholar] [CrossRef]
- Domínguez-Arispuro, D.M.; Cuevas-Rodríguez, E.O.; Milán-Carrillo, J.; León-López, L.; Gutiérrez-Dorado, R.; Reyes-Moreno, C. Optimal germination condition impacts on the antioxidant activity and phenolic acids profile in pigmented desi chickpea (Cicer arietinum L.) seeds. J. Food Sci. Technol. 2017, 55, 638–647. [Google Scholar] [CrossRef]
- Liu, H.; Kang, Y.; Zhao, X.; Liu, Y.; Zhang, X.; Zhang, S. Effects of elicitation on bioactive compounds and biological activities of sprouts. J. Funct. Foods 2019, 53, 136–145. [Google Scholar] [CrossRef]
- Thakur, M.; Bhattacharya, S.; Khosla, P.K.; Puri, S. Improving production of plant secondary metabolites through biotic and abiotic elicitation. J. Appl. Res. Med. Aromat. Plants 2019, 12, 1–12. [Google Scholar] [CrossRef]
- Świeca, M.; Baraniak, B. Influence of elicitation with H2O2 on phenolics content, antioxidant potential and nutritional quality of Lens culinaris sprouts. J. Sci. Food Agric. 2013, 94, 489–496. [Google Scholar] [CrossRef]
- Sá, A.G.A.; Moreno, Y.M.F.; Carciofi, B.A.M. Food processing for the improvement of plant proteins digestibility. Crit. Rev. Food Sci. Nutr. 2019, 1–20. [Google Scholar] [CrossRef]
- Świeca, M. Elicitation with abiotic stresses improves pro-health constituents, antioxidant potential and nutritional quality of lentil sprouts. Saudi J. Biol. Sci. 2015, 22, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Bohn, L.; Meyer, A.S.; Rasmussen, S.K. Phytate: Impact on environment and human nutrition. A challenge for molecular breeding. J. Zhejiang Univ. Sci. B 2008, 9, 165–191. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Sinha, A.K.; Makkar, H.P.; Becker, K. Dietary roles of phytate and phytase in human nutrition: A review. Food Chem. 2010, 120, 945–959. [Google Scholar] [CrossRef]
- Barakat, H.; Reim, V.; Rohn, S. Stability of saponins from chickpea, soy and faba beans in vegetarian, broccoli-based bars subjected to different cooking techniques. Food Res. Int. 2015, 76, 142–149. [Google Scholar] [CrossRef]
- Guajardo-Flores, D.; Serna-Saldívar, S.O.; Gutiérrez-Uribe, J.A. Evaluation of the antioxidant and antiproliferative activities of extracted saponins and flavonols from germinated black beans (Phaseolus vulgaris L.). Food Chem. 2013, 141, 1497–1503. [Google Scholar] [CrossRef]
- Wu, Z.; Song, L.; Feng, S.; Liu, Y.; He, G.; Yioe, Y.; Liu, S.Q.; Huang, D. Germination Dramatically Increases Isoflavonoid Content and Diversity in Chickpea (Cicer arietinum L.) Seeds. J. Agric. Food Chem. 2012, 60, 8606–8615. [Google Scholar] [CrossRef]
- Khang, D.T.; Dung, T.N.; Elzaawely, A.A.; Xuan, T.D. Phenolic Profiles and Antioxidant Activity of Germinated Legumes. Foods 2016, 5, 27. [Google Scholar] [CrossRef]
- Ampofo, J.O.; Ngadi, M. Stimulation of the phenylpropanoid pathway and antioxidant capacities by biotic and abiotic elicitation strategies in common bean (Phaseolus vulgaris) sprouts. Process. Biochem. 2020, 100, 98–106. [Google Scholar] [CrossRef]
- Ampofo, J.O.; Ngadi, M. Ultrasonic assisted phenolic elicitation and antioxidant potential of common bean (Phaseolus vulgaris) sprouts. Ultrason. Sonochem. 2020, 64, 104974. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Germination Conditions 1 | |||||
|---|---|---|---|---|---|
| Assay 2 | Process Variables 3 | Response Variables 4 | |||
| [H2O2] Concentration (mM) (X1) | Germination Time (h) (X2) | Total Phenolic Content (TPC) (mg GAE/100 g DW) YTPC | Total Flavonoid Content (TFC) (mg CE/100 g DW) YFC | Antioxidant Activity (AOX) (µmol TE/100 g DW) YAOX | |
| 1 | 5 (−1) | 12 (−1) | 58.45 | 13.41 | 1265.88 |
| 2 | 30 (+1) | 12 (−1) | 62.61 | 18.71 | 1332.59 |
| 3 | 5 (−1) | 72 (+1) | 89.56 | 21.23 | 1629.44 |
| 4 | 30 (+1) | 72 (+1) | 126.29 | 27.33 | 2117.18 |
| 5 | 0 (−1.414) | 42 (0) | 69.89 | 13.80 | 1369.40 |
| 6 | 35 (+1.414) | 42 (0) | 80.82 | 22.00 | 1772.74 |
| 7 | 17.5 (0) | 0 (−1.414) | 59.55 | 20.39 | 942.77 |
| 8 | 17.5 (0) | 84 (+1.414) | 147.69 | 30.58 | 2334.93 |
| 9 | 17.5 (0) | 42 (0) | 81.94 | 20.32 | 1573.07 |
| 10 | 17.5 (0) | 42 (0) | 76.85 | 21.92 | 1473.01 |
| 11 | 17.5 (0) | 42 (0) | 87.73 | 20.22 | 1651.33 |
| 12 | 17.5 (0) | 42 (0) | 75.54 | 20.77 | 1555.78 |
| 13 | 17.5 (0) | 42 (0) | 83.49 | 19.78 | 1529.90 |
| Regression Parameter Coefficients | |||
|---|---|---|---|
| Responses Variables 1 | |||
| Parameter | TPC (mg GAE/100 DW) | TFC (mg CE/100 DW) | AOX (µmol TE/100 g DW) |
| Coded Values | Coded Values | Coded Values | |
| Intercept | |||
| β0 | 81.1 | +20.6 | +1565.1 |
| Linear | |||
| β1, H2O2 concentration (X1) | +7.04 | +2.87 | +155.4 |
| β2, Germination time (X2) | +27.43 | +3.86 | +360.2 |
| Quadratic | |||
| β11, H2O2 concentration () | NS | −1.73 | NS |
| β22, Germination time () | +9.94 | +2.06 | NS |
| Interactive | |||
| β12, H2O2 concentration x Germination time (X1×2) | +8.14 | NS | +110.2 |
| p-value for model | <0.0001 | <0.0001 | <0.0001 |
| R2 | 0.956 | 0.967 | 0.933 |
| R2ajusted | 0.925 | 0.947 | 0.886 |
| p-value for lack of fit | 0.138 NS | 0.191 NS | 0.726 NS |
| CV (%) | 8.3 | 5.1 | 4.8 |
| Adequate precision (PRESS) | 17.0 | 23.8 | 14.1 |
| Responses | Predicted Models | Predicted Values | Experimental Values 2 |
| TPC | 129.47 | 138.8 ± 2.01 | |
| TFC | 27.86 | 27.8 ± 1.32 | |
| AOX | 2219.4 | 2347.2 ± 133 | |
| Property | UC | Control | OCS |
|---|---|---|---|
| Chemical composition (%DW) | |||
| Protein | 15.98 ± 0.15 b | 19.99 ± 0.59 a | 18.93 ± 0.49 a |
| Lipids | 6.72 ± 0.05 a | 7.73 ± 0.12 b | 7.96 ± 0.03 b |
| Ashes | 3.02 ± 0.02 a | 2.62 ± 0.11 b | 2.77 ± 0.01 b |
| Total carbohydrates | 74.27 ± 0.41 a | 69.66 ± 0.81 b | 71.34 ± 0.92 b |
| Antinutrient compounds | |||
| Phytic acid (mg PAE/100 g DW) | 140.03 ± 6.51 a | 115.71 ± 4.32 b | 97.13 ± 4.11 c |
| Saponins (mg DE/100 g DW) | 426.67 ± 18.1 b | 498.68 ± 16.3 a | 389.94 ± 12.6 c |
| Trypsin inhibitors (TIU/mg DW) | 6.76 ± 0.18 a | 6.24 ± 0.15 b | 6.34 ± 0.12 b |
| Bioactive compounds | |||
| Total phenolic (mg GAE/100 g DW) | 45.5 ± 1.45 c | 128.3 ± 1.01 b | 137.8 ± 2.01 a |
| Total flavonoids (mg CE/100 g DW) | 1 0.5 ± 0.81 b | 27.6 ± 1.01 a | 27.8 ± 1.32 a |
| Total isoflavones (µg FE/100 g DW) | 8.5 ± 1.6 c | 221.3 ± 9.2 b | 257.9 ± 8.1 a |
| Antioxidant activity (µ mol TE)/100 DW) | 953.9 ± 197 c | 2045.1 ± 101 b | 2347.2 ± 133 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
León-López, L.; Escobar-Zúñiga, Y.; Salazar-Salas, N.Y.; Mora Rochín, S.; Cuevas-Rodríguez, E.O.; Reyes-Moreno, C.; Milán-Carrillo, J. Improving Polyphenolic Compounds: Antioxidant Activity in Chickpea Sprouts through Elicitation with Hydrogen Peroxide. Foods 2020, 9, 1791. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9121791
León-López L, Escobar-Zúñiga Y, Salazar-Salas NY, Mora Rochín S, Cuevas-Rodríguez EO, Reyes-Moreno C, Milán-Carrillo J. Improving Polyphenolic Compounds: Antioxidant Activity in Chickpea Sprouts through Elicitation with Hydrogen Peroxide. Foods. 2020; 9(12):1791. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9121791
Chicago/Turabian StyleLeón-López, Liliana, Yudith Escobar-Zúñiga, Nancy Yareli Salazar-Salas, Saraid Mora Rochín, Edith Oliva Cuevas-Rodríguez, Cuauhtémoc Reyes-Moreno, and Jorge Milán-Carrillo. 2020. "Improving Polyphenolic Compounds: Antioxidant Activity in Chickpea Sprouts through Elicitation with Hydrogen Peroxide" Foods 9, no. 12: 1791. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9121791