From by-Product to Unconventional Vegetable: Preliminary Evaluation of Fresh Fava Hulls Highlights Richness in L-Dopa and Low Content of Anti-Nutritional Factor

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cropping Details and Sampling Procedures
2.2. Physical Analysis
2.3. Total Titratable Acidity, pH and Crude Protein Content
2.4. Ascorbic Acid Content and Total Phenols
2.5. Chlorophylls and Carotenoid Content
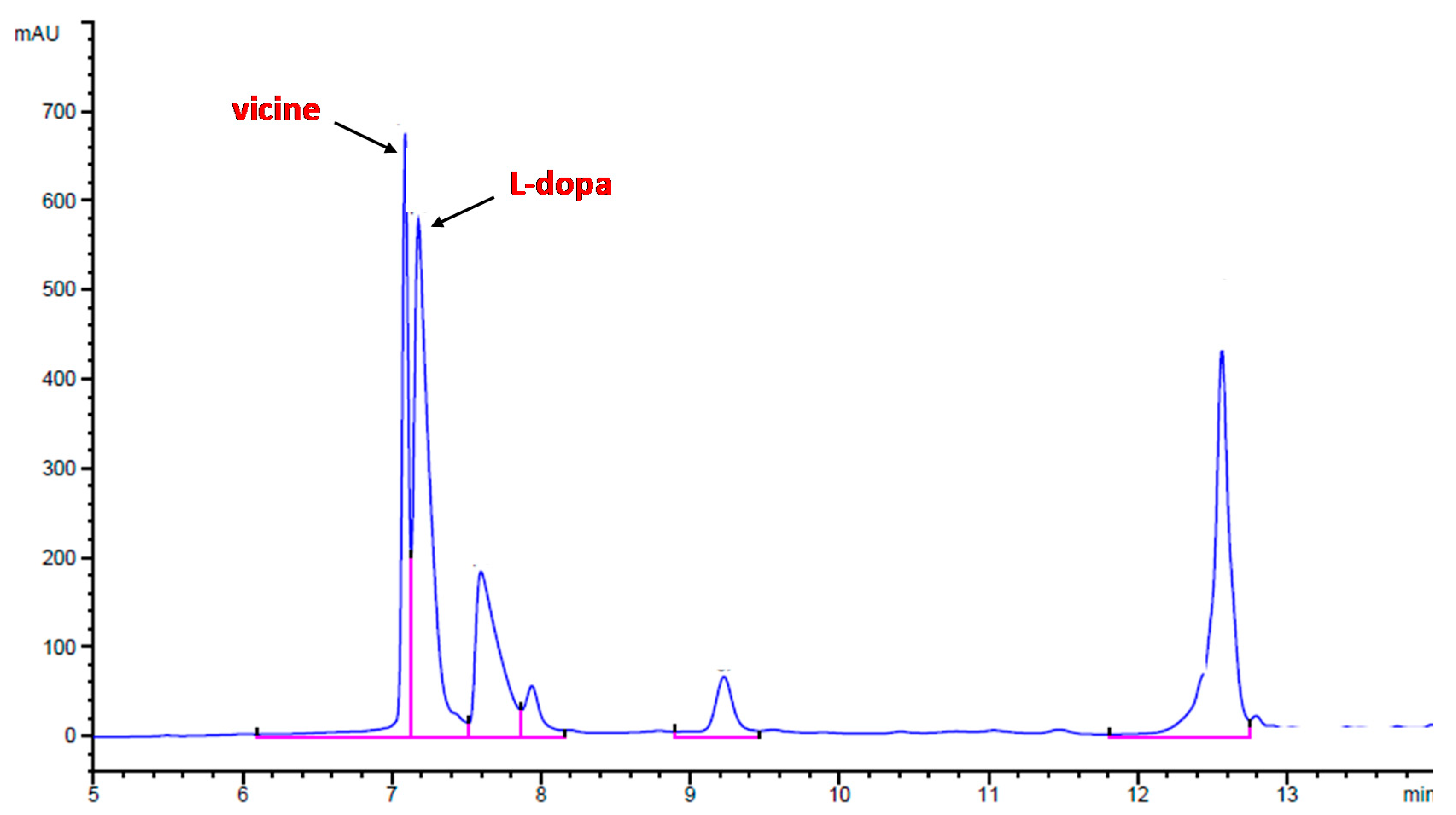
2.6. Content of Vicine and L-dopa
2.7. Statistical Analysis
3. Results
3.1. Yield and Colour Traits
3.2. Chemical Traits
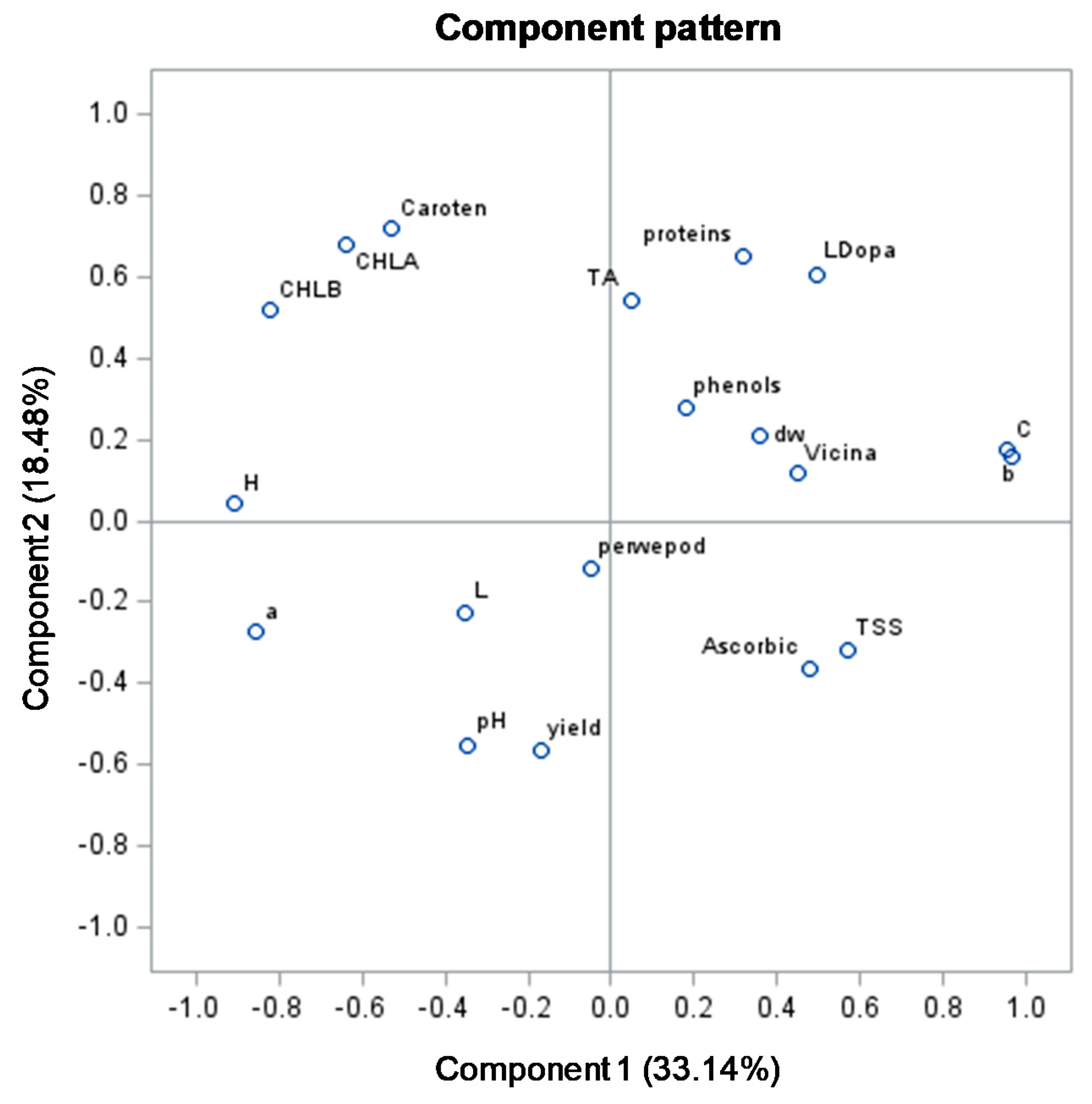
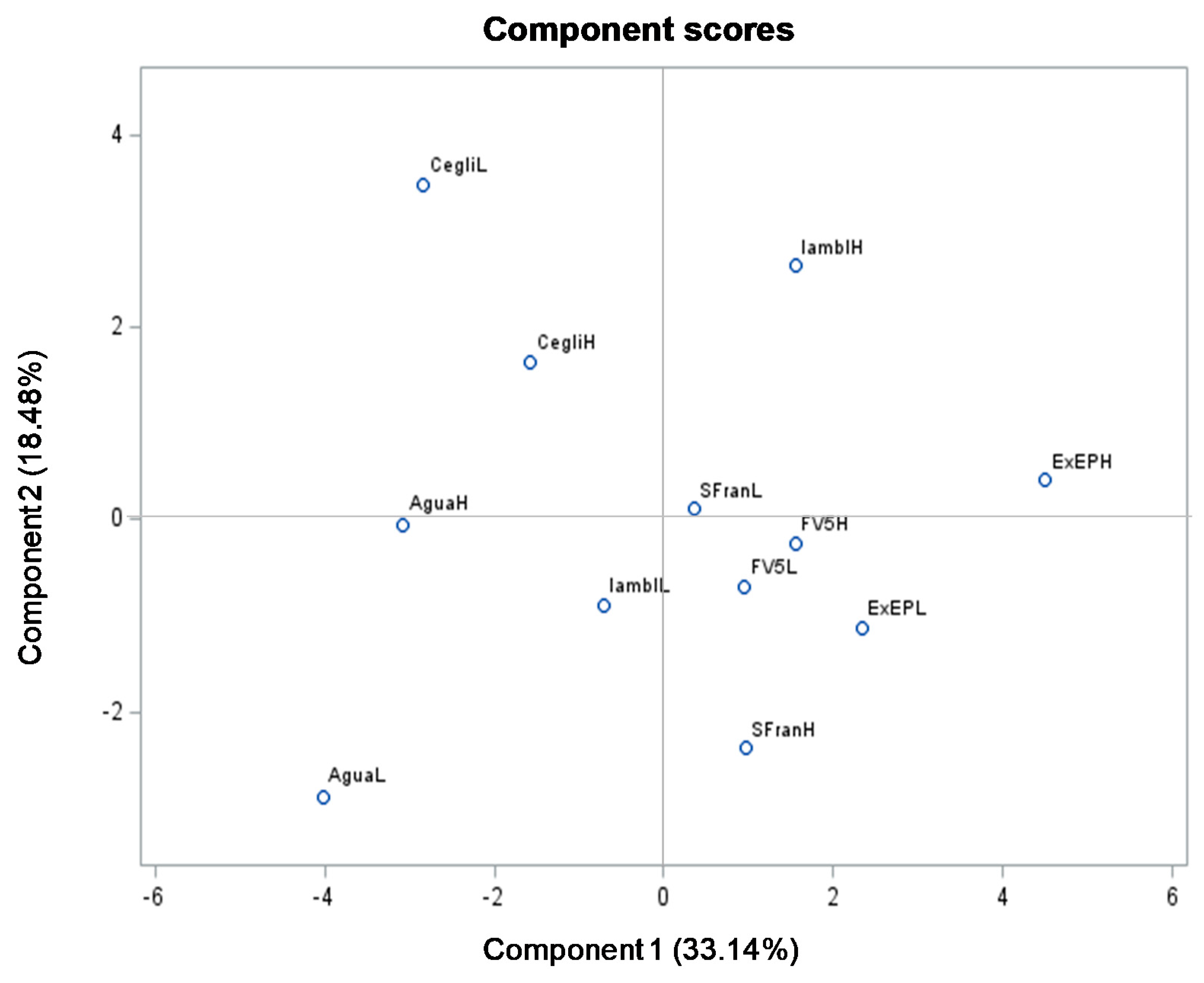
3.3. Principal Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wolosiak, R.; Worobiej, E.; Piecyk, M.; Druzynska, B.; Nowak, D.; Lewicki, P.P. Activities of amine and phenolic antioxidants and their changes in broad beans (Vicia faba) after freezing and steam cooking. Int. J. Food Sci. Technol. 2009, 45, 29–37. [Google Scholar] [CrossRef]
- dos Santos Rosa, E.A.; Savvas, D.; Barros, A.; Balliu, A.; Karapanos, I.; Weiss, J.; Gutiérrez-Cortines, M.E.; Ntatsi, G. The quality of leguminous vegetables as influenced by preharvest factors. Sci. Hortic. (Amst.) 2018, 232, 191–205. [Google Scholar] [CrossRef]
- De Cillis, F.; Leoni, B.; Massaro, M.; Renna, M.; Santamaria, P. Yield and quality of faba bean (Vicia faba L. var. major) genotypes as a vegetable for fresh consumption: A comparison between Italian landraces and commercial varieties. Agriculture 2019, 9, 253. [Google Scholar] [CrossRef] [Green Version]
- Korczyn, A.D.; Graff, E.; Rabey, J.M.; Vered, Y.; Shabtai, H. Improvement of parkinsonian features correlate with high plasma levodopa values after broad bean (Vicia faba) consumption. J. Neurol. Neurosurg. Psychiatry 2008, 55, 725–727. [Google Scholar] [CrossRef] [Green Version]
- Cappellini, M.; Fiorelli, G. Glucose-6-phosphate dehydrogenase deficiency. Lancet 2008, 371, 64–74. [Google Scholar] [CrossRef]
- Tesco Frozen Broad Beans. Available online: https://www.tesco.com/groceries/en-GB/products/282412496 (accessed on 6 December 2019).
- Bonduelle Canned Broad Beans. Available online: https://hollandshopper.nl/Bonduelle-tuinboontjes (accessed on 6 December 2019).
- Renna, M.; Signore, A.; Paradiso, V.M.; Santamaria, P. Faba greens, globe artichoke’s offshoots, crenate broomrape and summer squash greens: Unconventional vegetables of Puglia (Southern Italy) with good quality traits. Front. Plant Sci. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianco, V.V.; Renna, M.; Santamaria, P. Ortaggi Liberati L’ebook. Dieci Prodotti Straordinari Della Biodiversità Pugliese… e Qualcosa in Più.; Università degli Studi di Bari Aldo Moro: Bari, Italy, 2018; ISBN 978-88-6629-030-8. [Google Scholar]
- Signore, A.; Renna, M.; Santamaria, P. Agrobiodiversity of vegetable crops: Aspect, needs, and future perspectives. Annu. Plant Rev. Online 2019, 2, 1–24. [Google Scholar] [CrossRef]
- AOAC Official Methods of Analysis of AOAC International, 16th ed.; AOAC International: Arlington, VA, USA, 1995.
- Renna, M.; Gonnella, M.; Giannino, D.; Santamaria, P. Quality evaluation of cook-chilled chicory stems (Cichorium intybus L., Catalogna group) by conventional and sous vide cooking methods. J. Sci. Food Agric. 2014, 94, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Albanese, D.; Russo, L.; Cinquanta, L.; Brasiello, A.; Di Matteo, M. Physical and chemical changes in minimally processed green asparagus during cold-storage. Food Chem. 2007, 101, 274–280. [Google Scholar] [CrossRef]
- USDA Broadbeans, Immature Seeds, Raw. Available online: https://fdc.nal.usda.gov/fdc-app.html#/food-details/170377/nutrients (accessed on 15 October 2019).
- USDA Beans, Snap, Green, Raw. Available online: https://fdc.nal.usda.gov/fdc-app.html#/food-details/169961/nutrients (accessed on 26 November 2019).
- USDA Beans, Snap, Yellow, Raw. Available online: https://fdc.nal.usda.gov/fdc-app.html#/food-details/169320/nutrients (accessed on 27 November 2019).
- Harris, J.R. Ascorbic Acid: Biochemistry and Biomedical Cell Biology; Plenum Press: New York, NY, USA, 1996; ISBN 1461303257. [Google Scholar]
- European Food Safety Authority. Tolerable Upper Intake Levels for Vitamins and Minerals; European Food Safety Authority: Parma, Italy, 2006; ISBN 9291990140.
- Giusti, F.; Caprioli, G.; Ricciutelli, M.; Vittori, S.; Sagratini, G. Determination of fourteen polyphenols in pulses by high performance liquid chromatography-diode array detection (HPLC-DAD) and correlation study with antioxidant activity and colour. Food Chem. 2017, 221, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Vinson, J.A.; Hao, Y.; Su, X.; Zubik, L. Phenol antioxidant quantity and quality in foods: Vegetables. J. Agric. Food Chem. 1998, 46, 3630–3634. [Google Scholar] [CrossRef]
- Al Muzrakchi, A.; Gehani, A.A. G6PD deficiency. Qatar Med. J. 2004, 13, 46. [Google Scholar] [CrossRef]
- Goyoaga, C.; Burbano, C.; Cuadrado, C.; Varela, A.; Guillamón, E.; Pedrosa, M.M.; Muzquiz, M. Content and distribution of vicine, convicine and L-dopa during germination and seedling growth of two Vicia faba L. varieties. Eur. Food Res. Technol. 2008, 227, 1537–1542. [Google Scholar] [CrossRef]
- Pulkkinen, M.; Gautam, M.; Lampi, A.-M.; Ollilainen, V.; Stoddard, F.; Sontag-Strohm, T.; Salovaara, H.; Piironen, V. Determination of vicine and convicine from faba bean with an optimized high-performance liquid chromatographic method. Food Res. Int. 2015, 76, 168–177. [Google Scholar] [CrossRef]
- Gardiner, E.E.; Marquardt, R.R.; Kemp, G. Variation in vicine and convicine concentration of faba bean genotypes. Can. J. Plant Sci. 1982, 62, 589–592. [Google Scholar] [CrossRef]
- Ramírez-Moreno, J.M.; Salguero Bodes, I.; Romaskevych, O.; Duran-Herrera, M.C. Consumo de habas (Vicia faba) y enfermedad de Parkinson: Una fuente natural de L-dopa a tener en cuenta. Neurología 2015, 30, 375–376. [Google Scholar] [CrossRef] [PubMed]
- Hulya, A.; Sibel, E.; Sibel, O. Broad bean (Vicia Faba)—A natural source of L-dopa—Prolongs “on” periods in patients with Parkinson’s disease who have “on–off” fluctuations. Mov. Disord. 2000, 15, 164–166. [Google Scholar]
- Kempster, P.A.; Bogetic, Z.; Secombei, J.W.; Martin, H.D.; Balazs, N.D.; Wahlqvist, M.L. Motor effects of broad beans (Vicia faba) in Parkinson’s disease: Single dose studies. Asia Pac. J. Clin. Nutr. 1993, 2, 85–89. [Google Scholar] [PubMed]
- Rabey, J.M.; Vered, Y.; Shabtai, H.; Graff, E.; Korczyn, A.D. Improvement of parkinsonian features correlate with high plasma levodopa values after broad bean (Vicia faba) consumption. J. Neurol. Neurosurg. Psychiatry 1992, 55, 725–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vered, J.; Rabey, D.; Palevitch, I.; Grosskopf, A.; Harsat, A.; Yanowski, H.; Shabtai, E.; Graff, Y. Bioavailability of levodopa after consumption of Vicia faba seedlings by Parkinsonian patients and control subjects. Clin. Neuropharmacol. 1994, 17, 138–146. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cumulative Whole Pods Yield | Mean Pod Weight | Hulls | ||
|---|---|---|---|---|
| (Number m−2) | (g m−2) | (g) | (%) | |
| Density (Plants m−2) | ||||
| 4.16 | 86.3 ± 16.0 | 1794 ± 373 | 20.7 ± 1.7 | 74.7 ± 4.6 |
| 2.08 | 66.5 ± 16.5 | 1376 ± 403 | 20.6 ± 2.4 | 74.5 ± 4.6 |
| Genotype | ||||
| Aguadulce supersimonia | 78.6 ± 23.0 | 1839 ± 510 | 23.5 ± 2.1 a | 74.0 ± 4.5 ab |
| Extra-early purple | 86.4 ± 22.1 | 1737 ± 502 | 20.0 ± 1.0 b | 73.1 ± 5.9 b |
| Cegliese | 70.1 ± 23.7 | 1275 ± 487 | 18.0 ± 1.1 c | 73.7 ± 4.7 ab |
| FV5 | 73.0 ± 15.3 | 1529 ± 352 | 20.9 ± 0.7 b | 76.8 ± 4.0 a |
| Iambola | 73.0 ± 11.4 | 1502 ± 309 | 20.4 ± 1.4 b | 74.5 ± 4.2 ab |
| San Francesco | 77.4 ± 18.8 | 1627 ± 372 | 21.1 ± 0.8 b | 75.6 ± 3.8 ab |
| Significance | ||||
| Density | * | * | ns | ns |
| Genotype | ns | ns | *** | * |
| Density*Genotype | ns | ns | ns | ns |
| L | a* | b* | h° | C | |
|---|---|---|---|---|---|
| Density (Plants m−2) | |||||
| 4.16 | 52.9 ± 2.1 | −17.7 ± 0.8 | 34.8 ± 1.4 | 117.0 ± 0.6 | 39.0 ± 1.6 |
| 2.08 | 52.6 ± 2.2 | −17.4 ± 0.7 | 34.2 ± 1.2 | 117.0 ± 0.7 | 38.4 ± 1.3 |
| Genotype | |||||
| Aguadulce supersimonia | 53.4 ± 2.8 | −17.2 ± 0.8 | 33.7 ± 1.3 | 117.1 ± 0.5 ab | 37.8 ± 1.5 |
| Extra-early purple | 53.9 ± 2.5 | −17.3 ± 0.8 | 35.0 ± 1.3 | 116.4 ± 0.7 c | 39.0 ± 1.5 |
| Cegliese | 52.1 ± 2.3 | −17.9 ± 1.1 | 34.3 ± 1.6 | 117.5 ± 0.6 a | 38.7 ± 1.9 |
| FV5 | 51.9 ± 1.2 | −17.8 ± 0.6 | 34.7 ± 1.2 | 117.2 ± 0.5 ab | 39.1 ± 1.3 |
| Iambola | 52.6 ± 1.8 | −17.4 ± 0.6 | 34.7 ± 1.3 | 116.7 ± 0.4 bc | 38.8 ± 1.4 |
| San Francesco | 52.5 ± 1.8 | −17.6 ± 0.5 | 34.6 ± 1.2 | 116.9 ± 0.4 b | 38.9 ± 1.2 |
| Significance | |||||
| Density | ns | ns | ns | ns | ns |
| Genotype | ns | ns | ns | ** | ns |
| Density*Genotype | ns | ns | ns | ns | ns |
| pH | TSS | TA | Dry Weight | Protein | |
|---|---|---|---|---|---|
| (°Brix) | (g 100 g−1 FW) | ||||
| Density (Plants m−2) | |||||
| 4.16 | 5.5 ± 0.1 | 7.5 ± 1.1 | 0.25 ± 0.05 | 13.5 ± 1.0 | 2.63 ± 0.30 |
| 2.08 | 5.5 ± 0.1 | 7.3 ± 0.9 | 0.26 ± 0.06 | 14.0 ± 1.8 | 2.66 ± 0.34 |
| Genotype | |||||
| Aguadulce Supersimonia | 5.7 ± 0.1 a | 6.9 ± 0.6 | 0.24 ± 0.06 | 13.1 ± 0.7 | 2.46 ± 0.20 |
| Extra-early purple | 5.4 ± 0.1 b | 7.9 ± 0.9 | 0.25 ± 0.05 | 14.9 ± 1.5 | 2.80 ± 0.42 |
| Cegliese | 5.4 ± 0.2 b | 6.8 ± 1.4 | 0.26 ± 0.08 | 13.9 ± 0.7 | 2.79 ± 0.32 |
| FV5 | 5.5 ± 0.1 ab | 7.3 ± 0.7 | 0.22 ± 0.04 | 13.8 ± 2.4 | 2.64 ± 0.37 |
| Iambola | 5.4 ± 0.2 b | 7.7 ± 1.0 | 0.29 ± 0.02 | 14.1 ± 0.8 | 2.75 ± 0.15 |
| San Francesco | 5.5 ± 0.1 ab | 7.9 ± 0.7 | 0.26 ± 0.06 | 12.8 ± 1.3 | 2.44 ± 0.27 |
| Significance | |||||
| Density | ns | ns | ns | ns | ns |
| Genotype | ** | ns | ns | ns | ns |
| Density*Genotype | ns | ns | ns | ns | ns |
| Chl a | Chl b | TC | Ascorbic Acid | Total Phenols | Vicine | L-Dopa | |
|---|---|---|---|---|---|---|---|
| (µg g−1 FW) | (mg 100 g−1 FW) | ||||||
| Density (Plants m−2) | |||||||
| 4.16 | 17.0 ± 5.4 | 6.7 ± 2.7 | 7.2 ± 1.8 | 42 ± 18 | 563 ± 129 | 14.5 ± 5.3 | 178.4 ± 32.2 |
| 2.08 | 16.2 ± 4.9 | 7.2 ± 1.9 | 7.0 ± 1.9 | 37 ± 10 | 602 ± 114 | 10.4 ± 3.6 | 163.1 ± 27.3 |
| Variety | |||||||
| Aguadulce Supersimonia | 18.1 ± 5.2 ab | 8.3 ± 1.8 ab | 7.5 ± 1.5 ab | 32 ± 3 b | 443 ± 70 b | 13.1 ± 7.8 | 158.5 ± 20.2 |
| Extra-early purple | 12.8 ± 3.1 b | 4.5 ± 2.6 c | 6.0 ± 1.4 b | 53 ± 7 a | 546 ± 113 ab | 14.6 ± 6.8 | 185.5 ± 33.7 |
| Cegliese | 22.8 ± 5.4 a | 9.6 ± 2.0 a | 9.1 ± 2.3 a | 33 ± 2 b | 638 ± 50 a | 10.4 ± 3.4 | 176.3 ± 21.6 |
| FV5 | 13.3 ± 3.0 b | 6.2 ± 0.9 bc | 5.9 ± 1.2 b | 30 ± 3 b | 570 ± 120 ab | 13.7 ± 4.5 | 182.8 ± 28.5 |
| Iambola | 16.3 ± 4.2 ab | 6.9 ± 1.1 bc | 7.1 ± 1.3 ab | 30 ± 3 b | 649 ± 77 a | 10.3 ± 3.4 | 165.7 ± 44.7 |
| San Francesco | 16.2 ± 3.7 ab | 6.3 ± 1.7 bc | 7.2 ± 1.7 ab | 58 ± 20 a | 651151 a | 12.6 ± 1.7 | 155.7 ± 26.4 |
| Significance | |||||||
| Density | ns | ns | ns | ns | ns | ns | ns |
| Genotype | ** | *** | ** | *** | ** | ns | ns |
| Density*Genotype | ns | ns | ns | ns | ns | ns | ns |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Renna, M.; De Cillis, F.; Leoni, B.; Acciardi, E.; Santamaria, P. From by-Product to Unconventional Vegetable: Preliminary Evaluation of Fresh Fava Hulls Highlights Richness in L-Dopa and Low Content of Anti-Nutritional Factor. Foods 2020, 9, 159. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9020159
Renna M, De Cillis F, Leoni B, Acciardi E, Santamaria P. From by-Product to Unconventional Vegetable: Preliminary Evaluation of Fresh Fava Hulls Highlights Richness in L-Dopa and Low Content of Anti-Nutritional Factor. Foods. 2020; 9(2):159. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9020159
Chicago/Turabian StyleRenna, Massimiliano, Francesca De Cillis, Beniamino Leoni, Ermelinda Acciardi, and Pietro Santamaria. 2020. "From by-Product to Unconventional Vegetable: Preliminary Evaluation of Fresh Fava Hulls Highlights Richness in L-Dopa and Low Content of Anti-Nutritional Factor" Foods 9, no. 2: 159. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9020159