Use of Phycobiliproteins from Atacama Cyanobacteria as Food Colorants in a Dairy Beverage Prototype

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
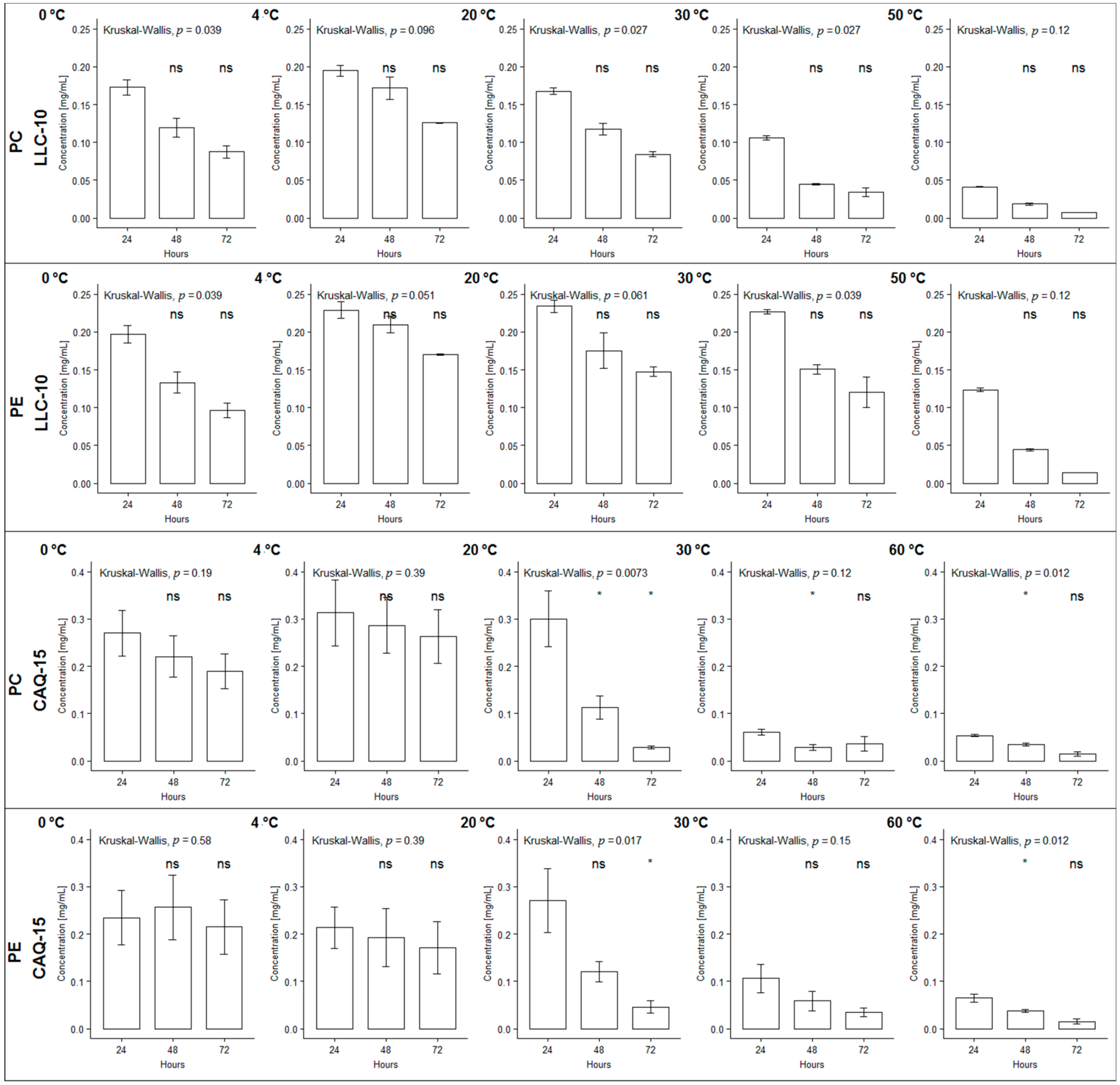
2.1. PBPs Stability under Different Temperature Regimes
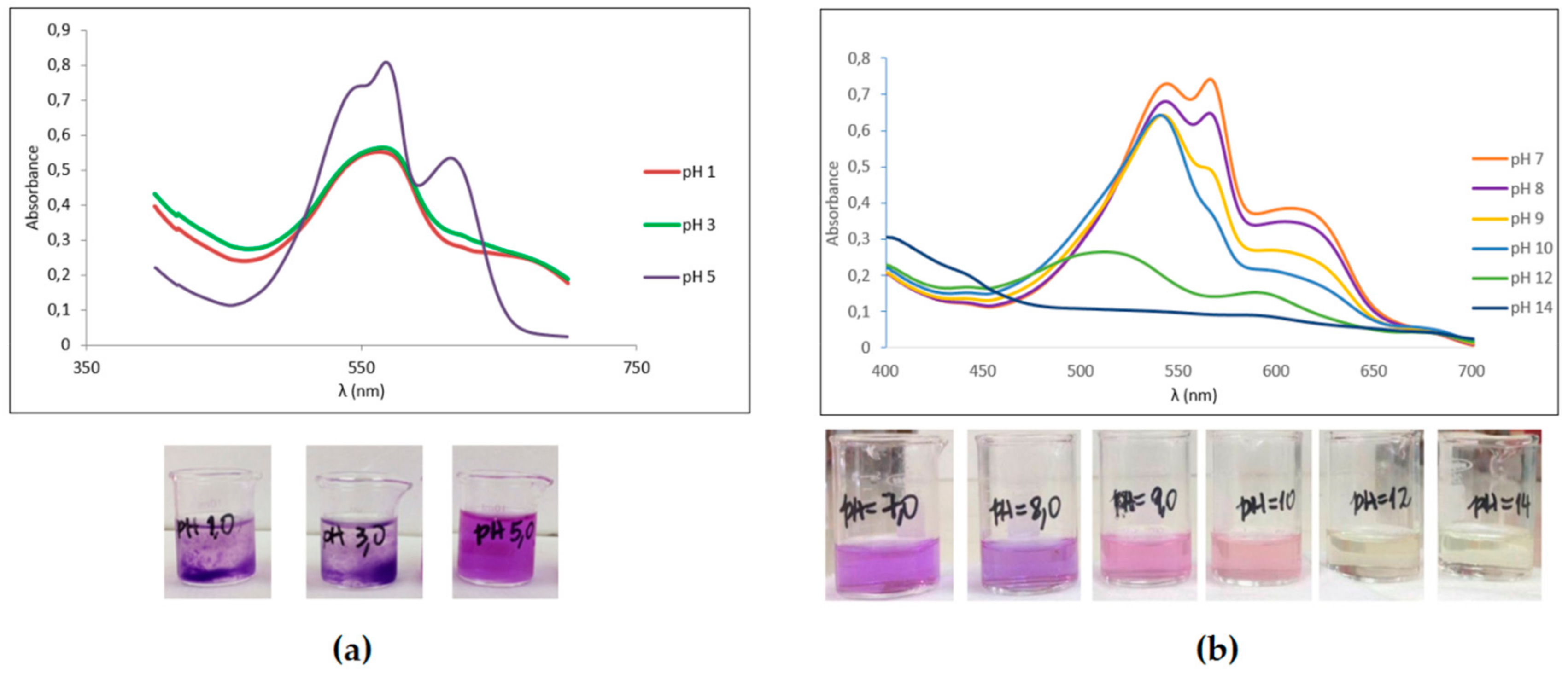
2.2. PBPs Stability under Different pH Regimes
2.3. Antioxidants Activity
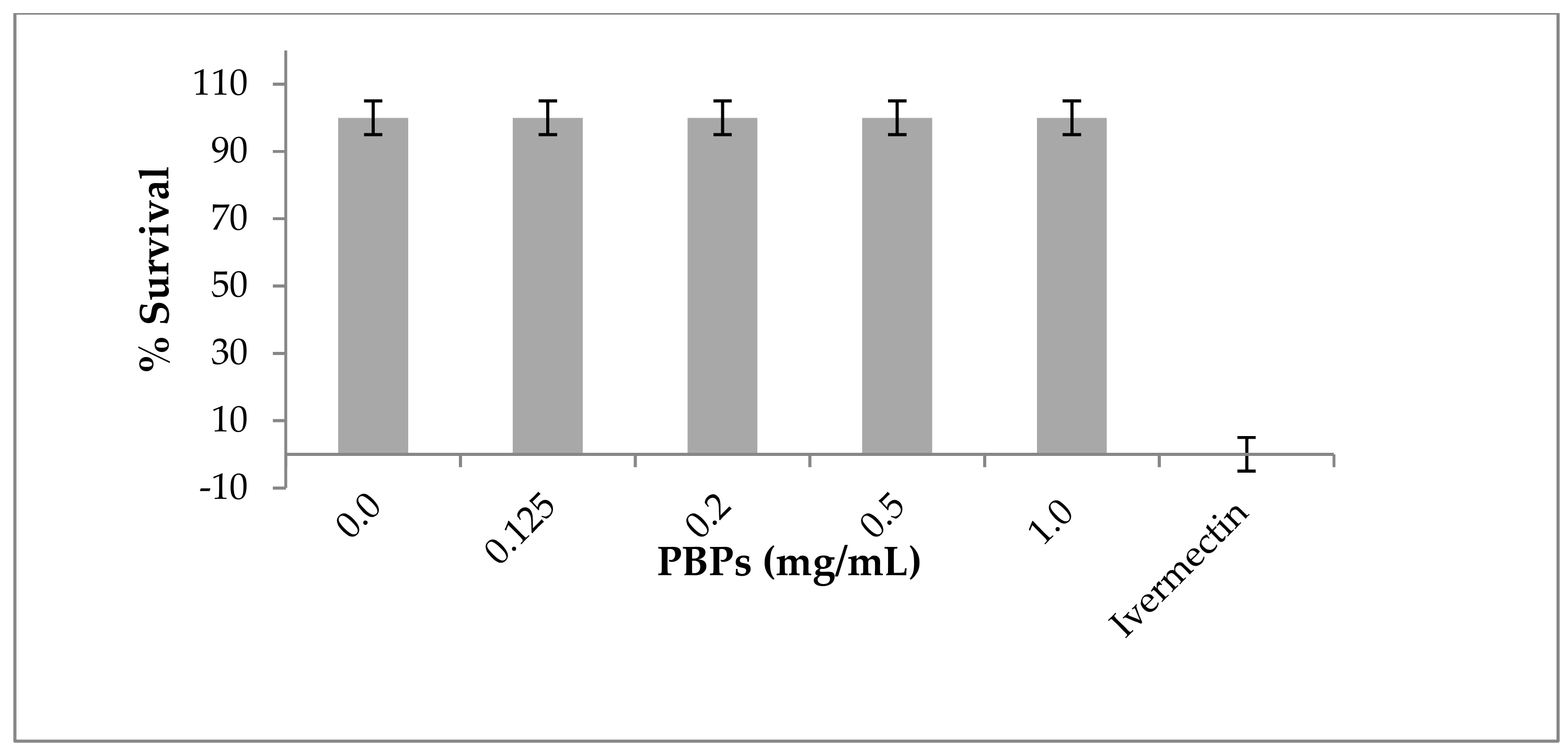
2.4. Toxicity of Phycobiliproteins against C. elegans
2.5. Sensory Test
2.6. Microbiological Assays
3. Materials and Methods
3.1. Strains and Culture Conditions of Cyanobacteria
3.2. Phycobiliprotein Purification
3.3. PBPs Stability
3.4. PBPs Antioxidant Capabilities
3.5. Toxicity Assays
3.6. Preparation and Sensory Analyses of Dairy Prototypes
3.7. Microbiological Analyses
3.8. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Marsac, T.D.N.; Houmard, J. Adaptation of cyanobacteria to environmental stimuli: New steps towards molecular mechanisms. FEMS Microb. 1993, 104, 119–189. [Google Scholar] [CrossRef]
- Whitton, B.; Potts, M. Introduction of cyanobacteria. In The Ecology of Cyanobacteria. Their Diversity in Time and Space; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2012; pp. 1–10. [Google Scholar]
- Abed, R.M.; Dobretsov, S.; Sudesh, K. Applications of cyanobacteria in biotechnology. J. Appl. Microbiol. 2009, 106, 1–12. [Google Scholar] [CrossRef]
- Vijayakumar, S.; Menakha, M. Pharmaceutical applications of cyanobacteria—A review. J. Acute Med. 2015, 5, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Bermejo, R.; Talavera, E.; Alvarez-Pez, J.; Orte, J. Chromatographic purification of biliproteins from Spirulina platensis. High-performance liquid chromatographic separation of their α and β subunits. J. Chromatogr. 1997, 778, 441–450. [Google Scholar] [CrossRef]
- Saini, D.K.; Pabbi, S.; Shukla, P. Cyanobacterial pigments: Perspectives and biotechnological approaches. Food Chem. Toxicol. 2018, 120, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Samsonoff, W.A.; MacColl, R. Biliproteins and phycobilisomes from cyanobacteria and red algae at the extremes of habitat. Arch. Microbiol. 2001, 176, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Marín, J.; Pavón, N.; Llópiz, A.; Fernández, J.; Delgado, L.; Mendoza, Y. Pycocyanobilin promotes PC12 cell survival and modulates immune and inflammatory genes and oxidative stress markers in acute hypoperfusion in rats. Toxicol. Appl. Pharmacol. Rev. 2013, 104, 119–190. [Google Scholar]
- Romay, C.H.; Armesto, J.; Remirez, D.; González, R.; Ledon, N.; García, I. Antioxidant and anti-inflammatory properties of C-phycocyanin from blue-g.reen algae. Inflamm. Res. 1998, 47, 36–41. [Google Scholar] [CrossRef] [PubMed]
- González, R.; Rodríguez, S.; Romay, C.; Ancheta, O.; González, A.; Armesto, J.; Remirez, D.; Merino, N. Anti-inflammatory activity of phycocyanin extract in acetic acid-induced colitis in rats. Pharmacol. Res. 1999, 39, 55–59. [Google Scholar] [CrossRef]
- Pleonsil, P.; Suwanwong, Y. An in vitro study of C-phycocyanin activity on protection of DNA and human erythrocyte membrane from oxidative damage. J. Chem. Pharm. Res. 2012, 5, 332–336. [Google Scholar]
- Liua, Q.; Wanga, Y.; Caoa, M.; Yang, T.; Maoa, H.; Suna, L.; Liu, G. Anti-allergic activity of R-phycocyanin from Porphyra haitanensis in antigen-sensitized mice and mast cells. Int. Immunopharmacol. 2015, 25, 465–473. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, L. Inhibitory effect of phycocyanin from Spirulina platensis on growth of human leukemia K562 cells. J. Appl. Phycol. 2000, 12, 125–130. [Google Scholar] [CrossRef]
- Sonani, R.; Rastogi, R.; Madamwar, D. Antioxidant potential of phycobiliproteins: Role in anti-aging research. Biochem. Anal. Biochem. 2015, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Munir, N.; Sharif, S.; Manzoor, S. Algae: A potent antioxidant source. Sky J. Microbiol. Res. 2013, 1, 22–31. [Google Scholar]
- Griffiths, J.C. Coloring foods and beverages. Food Technol. 2005, 59, 38–44. [Google Scholar]
- Borzelleca, J.F.; Capen, C.C.; Hallagan, J.B. Lifetime toxicity/carcinogenicity study of fd & c red no. 3 (erythrosine) in rats. Food Chem. Toxic. 1987, 10, 723–733. [Google Scholar]
- Mahmoud, N. Toxic effects of the synthetic food dye brilliant blue on liver, kidney and testes functions in rats. J. Egypt. Soc. Toxicol. 2006, 34, 77–84. [Google Scholar]
- Mohammed, M.; Mohammed, Y.; Abo-EL-Sooud, K.; Eleiwa, M.M. Embryotoxic and teratogenic effects of tartrazine in rats. Toxicol. Res. 2019, 35, 75–81. [Google Scholar]
- Shetty, M.J.S.; Geethalekshmi, P.; Mini, C. Natural Pigments as Potential Food Colourants: A Review. Trends Biosci. 2017, 10, 4057–4064. [Google Scholar]
- Romay, C.H.; Remirez, D.; González, R. Actividad Antioxidante de la ficocianina frente a radicales peroxílicos y la peroxidación microsomal. Rev. Cubana Investg. Biomed. 2001, 20, 38–41. [Google Scholar]
- Selvam, R.M.; Athinarayanan, G.; Nanthini, A.U.R.; Singh, A.J.A.; Kalirajan, K.; Selvakumar, P.M. Extraction of natural dyes from Curcuma longa, Trigonella foenum graecum and Nerium oleander, plants and their application in antimicrobial fabric. Ind. Crops Prod. 2015, 70, 84–90. [Google Scholar] [CrossRef]
- Mirjalili, M.; Nazarpoor, K.; Karimi, L. Eco-friendly dyeing of wool using natural dye from weld as co-partner with synthetic dye. J. Clean. Prod. 2011, 19, 1045–1051. [Google Scholar] [CrossRef]
- Frick, D. The coloration of food: Review of Progress in Coloration and Related Topics. Color. Technol. 2003, 33, 15–32. [Google Scholar] [CrossRef]
- Dweck, A.C. Natural ingredients for coloring and styling. Int. J. Cosmet. Sci. 2002, 24, 287–302. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, M.; Shabbir, M.; Mohammad, F. Natural Colorants: Historical, Processing and Sustainable Prospects. Nat. Prod. Bioprospect. 2017, 7, 123–145. [Google Scholar] [CrossRef] [Green Version]
- Chengaiah, B.; Rao, K.M.; Kumar, K.M.; Alagusundaram, M.; Chetty, C.M. Medicinal importance of natural dyes, a review. Int. J. PharmTech Res. 2010, 2, 144–154. [Google Scholar]
- Andriamanantena, M.; Danthu, P.; Cardon, D.; Fawbush, C.D.; Raonizafinimanana, B.; Razafintsalama, V.E.; Rakotonandrasana, S.R.; Ethève, A.; Petit, T.; Caro, Y. Malagasy Dye Plant Species: A Promising Source of Novel Natural Colorants with Potential Applications—A Review. Chem. Biodivers. 2019, 16, e1900442. [Google Scholar] [CrossRef]
- FDA. U.S. Food and Drug. Summary of Color Additives for Use in the United States in Foods, Drugs, Cosmetics and Medical Devices. Available online: https://www.fda.gov/industry/color-additive-inventories/summary-color-additives-use-united-states-foods-drugs-cosmetics-and-medical-devices (accessed on 14 January 2020).
- Olsen, K.; Jespersen, L.; Strømdahl, L. Heat and light stability of three natural blue colorants for use in confectionery and beverages. Eur. Food Res. Technol. 2005, 220, 261–266. [Google Scholar]
- Pandey, V.D.; Pandey, A.; Sharma, V. Biotechnological applications of cyanobacterial phycobiliproteins. Int. J. Curr. Microbiol. App. Sci. 2013, 2, 89–97. [Google Scholar]
- Bermejo, R. Phycocianins. In Cyanobacteria an Economic Perspective; Sharma, N.K., Rai, A.K., Stal, L.J., Eds.; Willey Blackwell, John Willey and Sons Ltd.: Hoboken, NJ, USA, 2014; pp. 209–225. [Google Scholar]
- Galetović, A.; Gómez-Silva, B. Inmovilización de cianobacterias filamentosas en perlas de alginato. Rev. Cienc. Salud. 1999, 3, 28–33. [Google Scholar]
- Seibert, M.; Connolly, J.S. Fluorescence properties of C-phycocyanin isolated from a thermophilic cyanobacterium. Photochem. Photobiol. 1984, 40, 267–271. [Google Scholar] [CrossRef]
- Couteau, C.; Baudry, S.; Roussakis, C.; Coiffard, L.J.M. Study of thermodegradation of phycocyanin from Spirulina platensis. Sci. Aliments 2004, 24, 415–421. [Google Scholar] [CrossRef]
- Patel, A.; Pawar, R.; Mishra, S.; Sonawane, S.; Ghosh, P.K. Kinetic studies on thermal denaturation of C-phycocyanin. Indian J. Biochem. Biophys. 2004, 41, 254–257. [Google Scholar] [PubMed]
- Antelo, F.S.; Costa, J.A.V.; Kalil, S.J. Thermal degradation kinetics of the phycocyanin from Spirulina platensis. Biochem. Eng. J. 2008, 41, 43–47. [Google Scholar] [CrossRef]
- Zhao, J.; Brand, J.J. Specific bleaching of phycobiliproteins from cyanobacteria and red algae at high temperature in vivo. Arch. Microbiol. 1989, 152, 447–452. [Google Scholar] [CrossRef]
- Tripathi, S.N.; Kapoor, S.; Shrivastava, A. Extraction and purification of an unusual phycoerythrin in a terrestrial desiccation tolerant cyanobacterium Lyngbya arboricola. J. Appl. Phycol. 2007, 19, 441–447. [Google Scholar] [CrossRef]
- Kaur, S.; Khattar, J.I.S.; Singh, Y.; Singh, D.P.; Ahluwalia, A.S. Extraction, purification and characterisation of phycocyanin from Anabaena fertilissima PUPCCC 410.5: As a natural and food grade stable pigment. Appl. Phycol. 2019, 31, 1685–1696. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Sonani, R.R.; Madamwar, D. Physico-chemical factors affecting the in vitro stability of phycobiliproteins from Phormidium rubidum A09DM. Bioresour. Technol. 2015, 190, 219–226. [Google Scholar] [CrossRef]
- Munawaroh, H.S.H.; Darojatun, K.; Gumilar, G.G.; Aisyah, S.; Wulandari, A.P. Characterization of phycocyanin from Spirulina fusiformis and its thermal stability. J. Phys. Conf. Ser. 2018, 1013, 012205. [Google Scholar] [CrossRef] [Green Version]
- Pumas, C.; Peerapornpisal, Y.; Vacharapiyasophon, P.; Leelapornpisid, P.; Boonchum, W.; Ishii, M.; Khanongnuch, C. Purification and characterization of a thermostable phycoerythrin from hot spring cyanobacterium Leptolyngbya sp. KC45. Int. J. Agric. Biol. 2012, 14, 121–125. [Google Scholar]
- Leu, J.; Lin, T.; Selvamani, M.J.P.; Chen, H.; Liang, J.; Pan, K. Characterization of a novel thermophilic cyanobacterial strain from taian hot springs in Taiwan for high CO2 mitigation and C-phycocyanin extraction. Process Biochem. 2013, 48, 41–48. [Google Scholar] [CrossRef]
- Liang, Y.; Kaczmarek, M.B.; Kasprzak, A.K.; Tang, J.; Shah, M.M.R.; Jin, P.; Daroch, M. Thermosynechococcaceae as a source of thermostable C-phycocyanins: Properties and molecular insights. Algal Res. 2018, 35, 223–235. [Google Scholar] [CrossRef]
- Rahman, D.Y.; Sarian, F.D.; van Wijk, A.; Martinez-Garcia, M.; van der Maarel, M.J.E.C. Thermostable phycocyanin from the red microalga Cyanidioschyzon merolae, a new natural blue food colorant. J. Appl. Phycol. 2017, 29, 1233–1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.K.; Shrivastav, A.; Mishra, S. Effect of preservatives for food grade C-PC from Spirulina platensis. Process Biochem. 2008, 43, 339–345. [Google Scholar] [CrossRef]
- Mishra, S.K.; Shrivastav, A.; Pancha, I.; Jain, D.; Mishra, S. Effect of preservatives for food grade C-phycoerythrin, isolated from marine cyanobacteria Pseudanabaena sp. Int. J. Biol. Macromol. 2010, 47, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, G.; Xiang, W.; Li, T.; He, H. Stability and antioxidant activity of food-grade phycocyanin isolated from Spirulina platensis. Int. J. Food Prop. 2016, 19, 2349–2362. [Google Scholar] [CrossRef] [Green Version]
- Martelli, G.; Folli, C.; Visai, L.; Daglia, M.; Ferrari, D. Thermal stability improvement of blue colorant C-phycocyanin from Spirulina platensis for food industry applications. Process Biochem. 2014, 49, 154–159. [Google Scholar] [CrossRef]
- Romay, C.H.; González, R.; Ledón, N.; Remirez, D.; Rimbau, V. C-Phycocyanin: A biliprotein with antioxidant, anti-Inflammatory and neuroprotective effects. Curr. Protein Pept. Sci. 2003, 4, 207–216. [Google Scholar] [CrossRef]
- Patel, S.N.; Sonani, R.R.; Jakharia, K.; Bhastana, B.; Patel, H.M.; Chaubey, M.G.; Singh, N.K.; Madamwar, D. Antioxidant activity and associated structural attributes of Halomicronema phycoerythrin. Int. J. Biol. Macromol. 2018, 111, 359–369. [Google Scholar] [CrossRef]
- Soni, B.; Trivedi, U.; Madamwar, D. A novel method of single step hydrophobic interaction chromatography for the purification of phycocyanin from Phormidium fragile and its characterization for antioxidant property. Bioresour. Technol. 2008, 188, 194. [Google Scholar] [CrossRef]
- Sonani, R.R.; Singh, N.K.; Kumar, J.; Thakar, D.; Madamwar, D. Concurrent purification and antioxidant activity of phycobiliproteinsfrom Lyngbya sp. A09DM: An antioxidant and anti-aging potential of phycoerythrin in Caenorhabditis elegans. Process Biochem. 2017, 49, 1757–1766. [Google Scholar] [CrossRef]
- Kuskoski, E.M.; Asuero, A.G.; Ana, M.; Troncoso, A.M.; Mancini-Filho, J.; Fett, R. Aplicación de diversos métodos químicos para determinar actividad antioxidante en pulpa de frutos. Ciênc. Tecnol. Aliment. Camp. 2005, 25, 726–732. [Google Scholar] [CrossRef] [Green Version]
- Leung, M.C.K.; Williams, P.L.; Benedetto, A.; Au, C.; Helmcke, K.J.; Aschner, M.; Meyer, J.N. Caenorhabditis elegans: An emerging model in biomedical and environmental toxicology. Toxicol. Sci. 2008, 106, 5–28. [Google Scholar] [CrossRef] [PubMed]
- Kaletta, T.; Hengartner, M.O. Finding function in novel targets: C. elegans as a model organism. Nat. Rev. Drug Discov. 2006, 5, 387–398. [Google Scholar] [CrossRef]
- Ju, J.; Saul, N.; Kochan, C.; Putschew, A.; Pu, Y.; Yin, L.; Steinberg, C.E. Cyanobacterial xenobiotics as evaluated by a Caenorhabditis elegans. Neurotoxicity screening test. Int. J. Environ. Res. Public Health 2014, 11, 4589–4606. [Google Scholar] [CrossRef] [Green Version]
- Gouveia, L.; Batista, A.; Miranda, A.; Empis, J.; Raymundo, A. Chlorella vulgaris biomass used as coloring source in traditional butter cookies. Innov. Food Sci. Emerg. Technol. 2007, 8, 433–436. [Google Scholar] [CrossRef]
- Hossain, A.K.M.M.; Brennan, M.A.; Mason, S.L.; Guo, X.; Zeng, X.A.; Brennan, C.S. The Effect of astaxanthin-rich microalgae “Haematococcus pluvialis” and wholemeal flours incorporation in improving the physical and functional properties of cookies. Foods 2017, 6, 57. [Google Scholar] [CrossRef] [Green Version]
- Caporgno, M.P.; Mathys, A. Trends in microalgae incorporation into innovative food products with potential health benefits. Front. Nutr. 2018, 5, 58. [Google Scholar] [CrossRef]
- Allen, M.; Arnon, D. Studies on nitrogen-fixing blue-green algae. I. Growth and nitrogen fixation by Anabaena cylindrica. Plant Physiol. 1955, 30, 366–372. [Google Scholar] [CrossRef] [Green Version]
- Ranjitha, K.; Kaushik, B.D. Purification of phycobiliproteins from Nostoc muscorum. J. Sci. Ind. Res. 2005, 64, 372–375. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, J.A.; Fleming, J.T. Basic culture methods. In Caenorhabditis Elegans: Modern Biological Analysis of an Organism; Epstein, H.F., Shakes, D.C., Eds.; Academic Press: San Diego, CA, USA, 1995; pp. 4–27. [Google Scholar]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [PubMed]
- Skantar, A.M.; Agama, K.; Meyer, S.L.F.; Carta, L.K.; Vinyard, B.T. Effects of geldanamycin on hatching and juvenile motility in Caenorhabditis elegans and Heterodera glycines. J. Chem. Ecol. 2005, 31, 2481–2491. [Google Scholar] [CrossRef] [Green Version]
- BBL™ MacConkey II Agar (BD). Available online: http://legacy.bd.com/ds/technicalCenter/inserts/L007388(10).pdf (accessed on 15 January 2020).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain PBP | Percentage of Remaining Phycobiliproteins at Different Interpolated Temperatures (°C) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 0% | 50% | 80% | ||||||||
| 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | ||
| LLC-10 | PC | 84.7 | 54.3 | 52.8 | 42.3 | 27.1 | 26.4 | 16.9 | 10.9 | 10.6 |
| PE | 106.9 | 76.9 | 70.4 | 53.4 | 38.5 | 35.2 | 21.4 | 15.4 | 14.1 | |
| CAQ-15 | PC | 68.1 | 57.8 | 49.5 | 34.0 | 28.9 | 24.7 | 13.6 | 11.6 | 9.9 |
| PE | 82.7 | 62.5 | 52.5 | 41.4 | 31.3 | 26.2 | 16.5 | 12.5 | 10.5 | |
| Sample | ABTS | FRAP |
|---|---|---|
| Methanol extract | µmoles TE/100 g fresh biomass | |
| CAQ-15 | 717 ± 61 | 50.23 ± 1.64 |
| LLC-10 | 641 ± 95 | 12.63 ± 0.73 |
| LLA-10 | 195 ± 38 | 13.14 ± 1.52 |
| Phycobiliprotein | µmoles TE/100 mg phycobiliprotein | |
| PE-CAQ-15 | 198 ± 45 | 0.92 ± 0.15 |
| PC-CAQ-15 | 312 ± 56 | 1.55 ± 0.10 |
| PC-LLA-10 | 205 ± 41 | 2.50 ± 0.15 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galetović, A.; Seura, F.; Gallardo, V.; Graves, R.; Cortés, J.; Valdivia, C.; Núñez, J.; Tapia, C.; Neira, I.; Sanzana, S.; et al. Use of Phycobiliproteins from Atacama Cyanobacteria as Food Colorants in a Dairy Beverage Prototype. Foods 2020, 9, 244. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9020244
Galetović A, Seura F, Gallardo V, Graves R, Cortés J, Valdivia C, Núñez J, Tapia C, Neira I, Sanzana S, et al. Use of Phycobiliproteins from Atacama Cyanobacteria as Food Colorants in a Dairy Beverage Prototype. Foods. 2020; 9(2):244. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9020244
Chicago/Turabian StyleGaletović, Alexandra, Francisca Seura, Valeska Gallardo, Rocío Graves, Juan Cortés, Carolina Valdivia, Javier Núñez, Claudia Tapia, Iván Neira, Sigrid Sanzana, and et al. 2020. "Use of Phycobiliproteins from Atacama Cyanobacteria as Food Colorants in a Dairy Beverage Prototype" Foods 9, no. 2: 244. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9020244