Sensory Improvement of a Pea Protein-Based Product Using Microbial Co-Cultures of Lactic Acid Bacteria and Yeasts

Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Materials, Ingredients, and Strains
2.2. Fermentation of Pea Protein Isolates
2.2.1. Inoculum Preparation
2.2.2. Preparation of Fermented Pea Protein Isolate
2.3. Fermentation Monitoring
2.3.1. Acidification Activity Measurement
2.3.2. Microbial analyses
2.3.3. Biochemical Analysis
Analyses Using HPLC–MS to Determine Sugar Content
Analyses Using HPLC to Determine Ethanol and Lactic Acid Concentrations
2.4. Sensory Evaluation
2.5. Aroma Compound Analysis
2.6. Statistical Analysis
3. Results and Discussion
3.1. The Impact of the Addition of Yeasts on Pea Fermentation by LAB
3.2. Modification of the Sensory Perception in the Presence of Yeasts
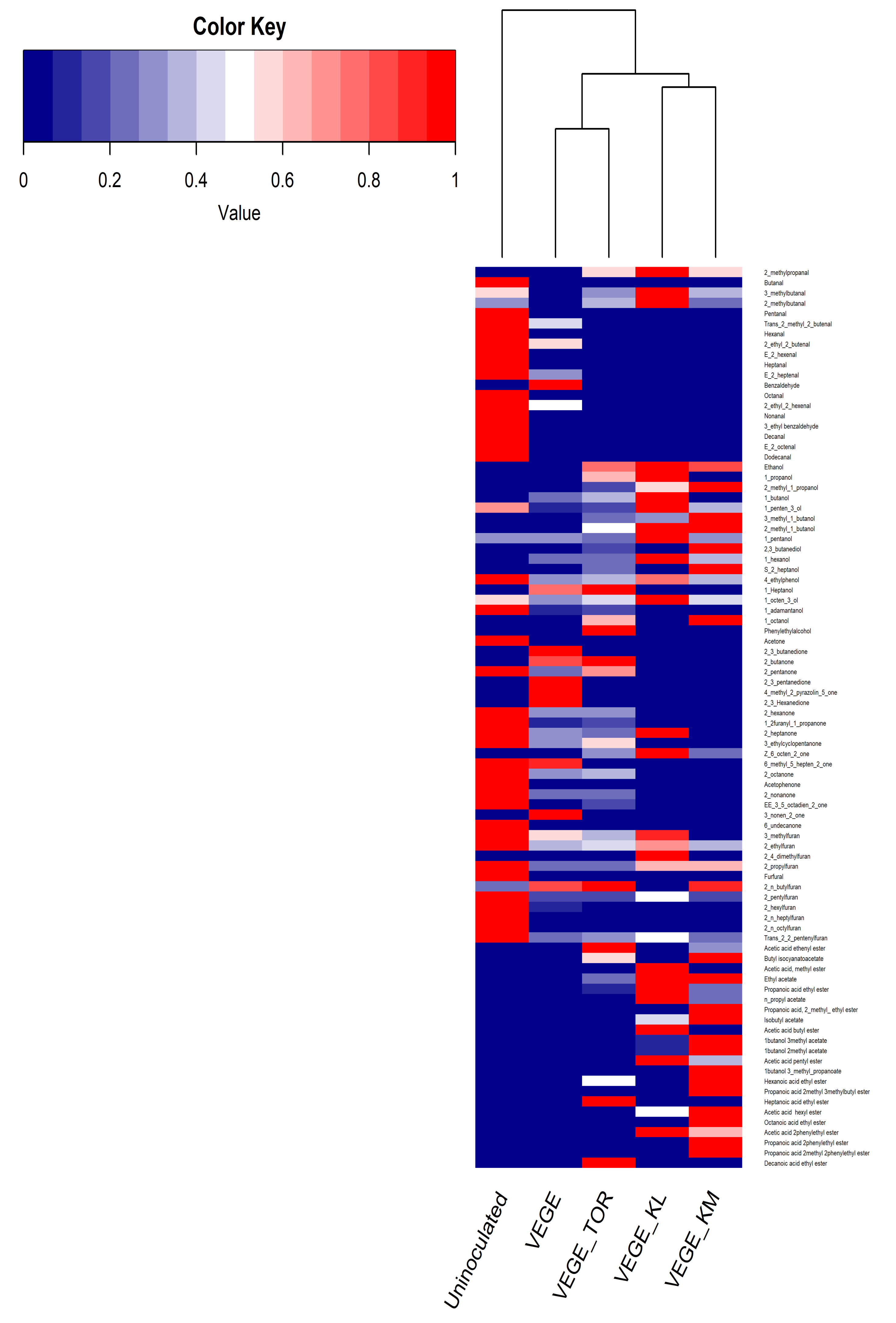
3.3. Characterization of Volatile Compounds Identified Using GC–MS Analysis
3.3.1. Volatile profiles of Uninoculated and Fermented Samples
3.3.2. Degradation of Off-Flavor Molecules in the Fermented Samples
3.4. Understanding the Sensory Perception Generated by Yeasts
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Graça, J.; Godinho, C.A.; Truninger, M. Reducing meat consumption and following plant-based diets: Current evidence and future directions to inform integrated transitions. Trends Food Sci. Technol. 2019, 91, 380–390. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.R.A.; Silva, M.M.N.; Ribeiro, B.D. Health Issues and Technological Aspects of Plant-based Alternative Milk. Food Res. Int. 2020, 108972. [Google Scholar] [CrossRef]
- McClements, D.J.; Newman, E.; McClements, I.F. Plant-based Milks: A Review of the Science Underpinning Their Design, Fabrication, and Performance. Compr. Rev. Food Sci. Food Saf. 2019, 18, 2047–2067. [Google Scholar] [CrossRef] [Green Version]
- Singh, N. Pulses: An overview. J. Food Sci. Technol. 2017, 54, 853–857. [Google Scholar] [CrossRef] [Green Version]
- Roland, W.S.; Pouvreau, L.; Curran, J.; van de Velde, F.; De Kok, P. Flavor Aspects of Pulse Ingredients. Cereal Chem. 2017, 94, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Dahl, W.J.; Foster, L.M.; Tyler, R.T. Review of the health benefits of peas (Pisum sativum L.). Br. J. Nutr. 2012, 108, S3–S10. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, A.; Khalil, I.A.; Ateeq, N.; Sayyar Khan, M. Nutritional quality of important food legumes. Food Chem. 2006, 97, 331–335. [Google Scholar] [CrossRef]
- Lu, Z.X.; He, J.F.; Zhang, Y.C.; Bing, D.J. Composition, physicochemical properties of pea protein and its application in functional foods. Crit. Rev. Food Sci. Nutr. 2019, 0, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Murat, C.; Bard, M.-H.; Dhalleine, C.; Cayot, N. Characterisation of odour active compounds along extraction process from pea flour to pea protein extract. Food Res. Int. 2013, 53, 31–41. [Google Scholar] [CrossRef]
- Schindler, S.; Zelena, K.; Krings, U.; Bez, J.; Eisner, P.; Berger, R.G. Improvement of the Aroma of Pea (Pisum sativum) Protein Extracts by Lactic Acid Fermentation. Food Biotechnol. 2012, 26, 58–74. [Google Scholar] [CrossRef]
- Bott, L.; Chambers, E. Sensory Characteristics of Combinations of Chemicals Potentially Associated with Beany Aroma in Foods. J. Sens. Stud. 2006, 21, 308–321. [Google Scholar] [CrossRef]
- Tangyu, M.; Muller, J.; Bolten, C.J.; Wittmann, C. Fermentation of plant-based milk alternatives for improved flavour and nutritional value. Appl. Microbiol. Biotechnol. 2019, 103, 9263–9275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blandino, A.; Al-Aseeri, M.E.; Pandiella, S.S.; Cantero, D.; Webb, C. Cereal-based fermented foods and beverages. Food Res. Int. 2003, 36, 527–543. [Google Scholar] [CrossRef]
- Spitaels, F.; Wieme, A.D.; Janssens, M.; Aerts, M.; Daniel, H.-M.; Van Landschoot, A.; De Vuyst, L.; Vandamme, P. The Microbial Diversity of Traditional Spontaneously Fermented Lambic Beer. PLoS ONE 2014, 9, e95384. [Google Scholar] [CrossRef]
- Aidoo, K.E.; Nout, M.J.R.; Sarkar, P.K. Occurrence and function of yeasts in Asian indigenous fermented foods. FEMS Yeast Res. 2006, 6, 30–39. [Google Scholar] [CrossRef]
- Park, Y.K.; Lee, J.H.; Mah, J.-H. Occurrence and reduction of biogenic amines in traditional Asian fermented soybean foods: A review. Food Chem. 2019, 278, 1–9. [Google Scholar] [CrossRef]
- Onda, T.; Yanagida, F.; Uchimura, T.; Tsuji, M.; Ogino, S.; Shinohara, T.; Yokotsuka, K. Analysis of Lactic Acid Bacterial Flora during Miso Fermentation. Food Sci. Technol. Res. 2003, 9, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.M.; Ostenfeld Larsen, T.; Schnürer, J. Production of volatile compounds by Rhizopus oligosporus during soybean and barley tempeh fermentation. Int. J. Food Microbiol. 2007, 113, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Wang, P.; Zhang, X.; Zhang, J.; Sun, Y.; Gao, L.; Wang, W. High-throughput sequencing approach to characterize dynamic changes of the fungal and bacterial communities during the production of sufu, a traditional Chinese fermented soybean food. Food Microbiol. 2020, 86, 103340. [Google Scholar] [CrossRef]
- Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; Cesare, A.D.; Hilbert, F.; Lindqvist, R.; et al. Update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA 10: Suitability of taxonomic units notified to EFSA until March 2019. EFSA J. 2019, 17, e05753. [Google Scholar] [CrossRef] [Green Version]
- Sieuwerts, S.; de Bok, F.A.M.; Hugenholtz, J.; van Hylckama Vlieg, J.E.T. Unraveling Microbial Interactions in Food Fermentations: From Classical to Genomics Approaches. Appl. Environ. Microbiol. 2008, 74, 4997–5007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindler, S.; Wittig, M.; Zelena, K.; Krings, U.; Bez, J.; Eisner, P.; Berger, R.G. Lactic fermentation to improve the aroma of protein extracts of sweet lupin (Lupinus angustifolius). Food Chem. 2011, 128, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Yousseef, M.; Lafarge, C.; Valentin, D.; Lubbers, S.; Husson, F. Fermentation of cow milk and/or pea milk mixtures by different starter cultures: Physico-chemical and sensorial properties. LWT Food Sci. Technol. 2016, 69, 430–437. [Google Scholar] [CrossRef] [Green Version]
- Ben-Harb, S.; Saint-Eve, A.; Panouillé, M.; Souchon, I.; Bonnarme, P.; Dugat-Bony, E.; Irlinger, F. Design of microbial consortia for the fermentation of pea-protein-enriched emulsions. Int. J. Food Microbiol. 2019, 293, 124–136. [Google Scholar] [CrossRef]
- Chiba, H.; Takahashi, N.; Sasaki, R. Enzymatic Improvement of Food Flavor II. Removal of Beany Flavor from Soybean Products by Aldehyde Dehydrogenase. Agric. Biol. Chem. 1979, 43, 1883–1889. [Google Scholar] [CrossRef]
- Hirst, M.B.; Richter, C.L. Review of Aroma Formation through Metabolic Pathways of Saccharomyces cerevisiae in Beverage Fermentations. Am. J. Enol. Vitic. 2016, 67, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Picque, D.; Corrieu, G. Characterization and classification of lactic acid bacteria based on their acidification kinetics. Food Sci. Technol. Lebensm. Wiss. Technol. 1992, 25, 181–186. [Google Scholar]
- Le Boucher, C.; Courant, F.; Jeanson, S.; Chereau, S.; Maillard, M.-B.; Royer, A.-L.; Thierry, A.; Dervilly-Pinel, G.; Le Bizec, B.; Lortal, S. First mass spectrometry metabolic fingerprinting of bacterial metabolism in a model cheese. Food Chem. 2013, 141, 1032–1040. [Google Scholar] [CrossRef]
- Merico, A.; Galafassi, S.; PiÅ¡kur, J.; Compagno, C. The oxygen level determines the fermentation pattern in Kluyveromyces lactis. FEMS Yeast Res. 2009, 9, 749–756. [Google Scholar] [CrossRef] [Green Version]
- Lane, M.M.; Morrissey, J.P. Kluyveromyces marxianus: A yeast emerging from its sister’s shadow. Fungal Biol. Rev. 2010, 24, 17–26. [Google Scholar] [CrossRef]
- Hu, Y.; Piao, C.; Chen, Y.; Zhou, Y.; Wang, D.; Yu, H.; Xu, B. Soybean residue (okara) fermentation with the yeast Kluyveromyces marxianus. Food Biosci. 2019, 31, 100439. [Google Scholar] [CrossRef]
- McMahon, K.M.; Culver, C.; Castura, J.C.; Ross, C.F. Perception of carbonation in sparkling wines using descriptive analysis (DA) and temporal check-all-that-apply (TCATA). Food Qual. Prefer. 2017, 59, 14–26. [Google Scholar] [CrossRef]
- Fonseca, G.G.; Heinzle, E.; Wittmann, C.; Gombert, A.K. The yeast Kluyveromyces marxianus and its biotechnological potential. Appl. Microbiol. Biotechnol. 2008, 79, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Lederer, C.L.; Bodyfelt, F.W.; McDaniel, M.R. The Effect of Carbonation Level on the Sensory Properties of Flavored Milk Beverages. J. Dairy Sci. 1991, 74, 2100–2108. [Google Scholar] [CrossRef]
- Verstrepen, K.J.; Derdelinckx, G.; Dufour, J.-P.; Winderickx, J.; Thevelein, J.M.; Pretorius, I.S.; Delvaux, F.R. Flavor-active esters: Adding fruitiness to beer. J. Biosci. Bioeng. 2003, 96, 110–118. [Google Scholar] [CrossRef]
- Azarnia, S.; Boye, J.I.; Warkentin, T.; Malcolmson, L. Changes in volatile flavour compounds in field pea cultivars as affected by storage conditions. Int. J. Food Sci. Technol. 2011, 46, 2408–2419. [Google Scholar] [CrossRef]
- Murat Chloe Gourrat, K.; Jerosch, H.; Cayot, N. Analytical comparison and sensory representativity of SAFE, SPME, and Purge and Trap extracts of volatile compounds from pea flour. Food Chem. 2012, 135, 913–920. [Google Scholar] [CrossRef]
- Buttery, R.G.; Orts, W.J.; Takeoka, G.R.; Nam, Y. Volatile Flavor Components of Rice Cakes. J. Agric. Food Chem. 1999, 47, 4353–4356. [Google Scholar] [CrossRef]
- Heng, L.; Vincken, J.-P.; Koningsveld G van Legger, A.; Gruppen, H.; Boekel T van Roozen, J.; Voragen, F. Bitterness of saponins and their content in dry peas. J. Sci. Food Agric. 2006, 86, 1225–1231. [Google Scholar] [CrossRef]
- Achouri, A.; Boye, J.; Zamani, Y. Identification of volatile compounds in soymilk using solid-phase microextraction-gas chromatography. Food Chem. 2006, 99, 759–766. [Google Scholar] [CrossRef]
- Blagden, T.D.; Gilliland, S.E. Reduction of Levels of Volatile Components Associated with the “Beany” Flavor in Soymilk by Lactobacilli and Streptococci. J. Food Sci. 2005, 70, M186–M189. [Google Scholar] [CrossRef]
- Ben-Harb, S.; Irlinger, F.; Saint-Eve, A.; Panouillé, M.; Souchon, I.; Bonnarme, P. Versatility of microbial consortia and sensory properties induced by the composition of different milk and pea protein-based gels. LWT 2020, 118, 108720. [Google Scholar] [CrossRef]
- Smit, B.A.; Engels, W.J.M.; Smit, G. Branched chain aldehydes: Production and breakdown pathways and relevance for flavour in foods. Appl. Microbiol. Biotechnol. 2009, 81, 987–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vong, W.C.; Liu, S.-Q. Bioconversion of green volatiles in okara (soybean residue) into esters by coupling enzyme catalysis and yeast (Lindnera saturnus) fermentation. Appl. Microbiol. Biotechnol. 2018, 102, 10017–10026. [Google Scholar] [CrossRef]
- Peinado, R.A.; Moreno, J.; Bueno, J.E.; Moreno, J.A.; Mauricio, J.C. Comparative study of aromatic compounds in two young white wines subjected to pre-fermentative cryomaceration. Food Chem. 2004, 84, 585–590. [Google Scholar] [CrossRef]
- Engan, S. Organoleptic threshold values of some alcohols and esters in beer. J. Inst. Brew. 1972, 78, 33–36. [Google Scholar] [CrossRef]
- Dennis, E.G.; Keyzers, R.A.; Kalua, C.M.; Maffei, S.M.; Nicholson, E.L.; Boss, P.K. Grape Contribution to Wine Aroma: Production of Hexyl Acetate, Octyl Acetate, and Benzyl Acetate during Yeast Fermentation Is Dependent upon Precursors in the Must. J. Agric. Food Chem. 2012, 60, 2638–2646. [Google Scholar] [CrossRef]
- Pires, E.J.; Teixeira, J.A.; Brányik, T.; Vicente, A.A. Yeast: The soul of beer’s aroma—A review of flavour-active esters and higher alcohols produced by the brewing yeast. Appl. Microbiol. Biotechnol. 2014, 98, 1937–1949. [Google Scholar] [CrossRef] [Green Version]
- Meilgaard, M.C.; Dalgliesh, C.E.; Clapperton, J.F. Beer flavour terminology1. J. Inst. Brew. 1979, 85, 38–42. [Google Scholar] [CrossRef]
- Plata, C.; Millán, C.; Mauricio, J.C.; Ortega, J.M. Formation of ethyl acetate and isoamyl acetate by various species of wine yeasts. Food Microbiol. 2003, 20, 217–224. [Google Scholar] [CrossRef]
- Lyumugabe, F.; Bajyana Songa, E.; Wathelet, J.P.; Thonart, P. Volatile compounds of the traditional sorghum beers “ikigage” brewed with Vernonia amygdalina “umubirizi”. Cerevisia 2013, 37, 89–96. [Google Scholar] [CrossRef]
- Hazelwood, L.A.; Daran, J.-M.; van Maris, A.J.A.; Pronk, J.T.; Dickinson, J.R. The Ehrlich Pathway for Fusel Alcohol Production: A Century of Research on Saccharomyces cerevisiae Metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.-Q.; Holland, R.; Crow, V.L. Esters and their biosynthesis in fermented dairy products: A review. Int. Dairy J. 2004, 14, 923–945. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
|
Bacteria Biomass (× 108 CFU/mL) |
Yeast Biomass (× 107 CFU/mL) | Kinetic Parameters (g/L) | |||||
|---|---|---|---|---|---|---|---|
| at tf | at t0 | at tf * | tf (h) | Total Residual Sugar at tf | Lactic Acid at tf | Ethanol at tf | |
| VEGE047 | 2.9 ± 0.2 b | - | - | 13.1 ± 0.5 ab | 25.4 ± 0.3 a | 3.5 ± 0.03 b | - |
| VEGE047 + K. marxianus | 4.2 ± 0.6 ab | 1.4 ± 0.1 b | 4.4 ± 0.5 a | 14.9 ± 0.9 a | 13.2 ± 0.1 d | 3.0 ± 0.05 d | 4.8 ± 0.007 a |
| VEGE047 + K. lactis | 6.0 ± 1.6 a | 3.4 ± 0.6 a | 4.1 ± 0.7 a | 13.1 ± 0.4 ab | 20.8 ± 0.6 b | 3.6 ± 0.03 a | 1.7 ± 0.004 c |
| VEGE047 + T. delbrueckii | 5.4 ± 0.3 a | 0.4 ± 0.1 b | 0.7 ± 0.1 a | 12.3 ± 0.01 b | 16 ± 0.3 c | 3.3 ± 0.02 c | 4.2 ± 0.02 b |
| VEGE | VEGE + K. marxianus | VEGE + K. lactis | VEGE + T. delbrueckii | |
|---|---|---|---|---|
| Global intensity | 4.660 b | 7.033 a | 6.390 a | 6.900 a |
| Sour | 2.257 b | 5.000 a | 4.250 a | 5.400 a |
| Bitter | 1.837 a | 3.200 a | 2.840 a | 2.867 a |
| Astringent | 3.730 a | 4.733 a | 4.243 a | 4.933 a |
| Tangy | 0.417 c | 5.267 a | 3.500 b | 5.200 a |
| Sparkling | 0.050 d | 7.467 b | 4.817 c | 9.367 a |
| Green flavor/Vegetal | 2.637 a | 0.733 b | 1.533 ab | 0.867 b |
| Leguminous plant | 3.723 a | 1.147 b | 1.500 b | 0.800 b |
| Citrus fruit | 0.400 a | 0.300 a | 0.850 a | 0.200 a |
| Nut | 0.267 a | 0.167 a | 0.333 a | 0.200 a |
| Beer/yeast | 0.183 c | 5.000 a | 3.150 b | 2.800 b |
| Sourdough | 0.243 b | 1.260 ab | 1.380 ab | 1.800 a |
| Cultured apple cider | 0.200 a | 1.340 a | 1.067 a | 1.067 a |
| Volatile Compounds | Descriptors | Uninoculated | VEGE | VEGE + K. marxianus | VEGE + K. lactis | VEGE + T. delbrueckii | Detection Threshold a |
|---|---|---|---|---|---|---|---|
| 2-pentylfuran | Musty/earthy, mushroom, floral, buttery, rancid, green | 304.3 | 46.4 | 56.7 | 148 | 55 | 6 |
| Hexanal | Green, grass | 181.3 | <DL | <DL | <DL | <DL | 4.5 |
| 2-ethylfuran | Beany, earthy, malty, sweet | 77.7 | 28.1 | 29.7 | 56.7 | 32.8 | |
| Butanal | Pungent, green, malty, chocolate, cocoa | 54.9 | <DL | <DL | <DL | <DL | |
| 1-penten-3-ol | Green, vegetable, fruity | 19.6 | 3.7 | 11 | 28.1 | 5.6 | |
| (E)-2-octenal | Green, cucumber, musty/earthy, waxy, fatty, grass, banana, sweet | 10.3 | <DL | <DL | <DL | <DL | 3 |
| Nonanal | Aldehydic, fatty, green, geranium, floral, soapy, citrus, waxy | 8.1 | <DL | <DL | <DL | <DL | 1 |
| Heptanal | Green, fresh, fatty | 5.7 | <DL | <DL | <DL | <DL | 3 |
| 2-nonanone | Green, earthy, grassy, fruity, sweet, | 5.0 | <DL | <DL | <DL | 1.2 | |
| 2-methylpropanal | Aldehylic, grass, green, floral | 4.0 | 0.7 | 40.9 | 70 | 39.6 | 6 |
| 1-octen-3-ol | Mushroom, earthy, burnt, green, vegetable, stale | 3.9 | 2.2 | 2.9 | 6.7 | 2.9 | 1 |
| (E)-2-heptenal | Pungent green, fatty | 2.7 | 0.8 | <DL | <DL | <DL | 13 |
| 2-octanone | Green, floral, soapy, fruity, fatty | 2.5 | 0.7 | <DL | 0.9 | 0.8 | |
| Octanal | Aldehylic, green, soapy, citrus-like, sweet, waxy, fruity | 2.5 | <DL | <DL | <DL | <DL | 0.7 |
| (E)-2-Hexenal | Tea-like, green grass, almond, cherry, juicy, rancid | 2.4 | <DL | <DL | <DL | <DL | 17 |
| Decanal | Fresh, marine, aldehydic, iodized, soapy, grapefruit, bitter, sweet | 1.04 | <DL | <DL | <DL | <DL | 0.1 |
| 6-methyl-5-hepten-2-one | Nutty, moldy, green, vegetable, citrus | 0.5 | 0.5 | <DL | <DL | <DL | 2 |
| Trans-2-methyl-2-butenal | Strong green-type odor and a fruity flavor | 0.3 | 0.1 | <DL | <DL | <DL | |
| 1-hexanol | Green, musty/earthy, peanut hull, chemical-like, fruity, grassy | <DL | 126.5 | 175.4 | 500.1 | 116.7 | 500 |
| 1-octanol | Moss, mushroom, green, vegetable, fatty, waxy, citrus, floral | <DL | <DL | 4.4 | <DL | 2.9 |
| Esters Compounds | Odor Descriptor * | Threshold Level a | Threshold Level in Beer b | VEGE + K. marxianus | VEGE + K. lactis | VEGE + T. delbrueckii |
|---|---|---|---|---|---|---|
| Ethyl acetate | Ethereal, fruity, sweet, grape and rum-like | 12,000 | 25,000 | 2095 | 3040 | 633 |
| Propyl acetate | Solvent-like pungency, lifting, fusel, amyl alcohol, sweet and fruity | 4700 | 9.2 | 39.1 | >DL | |
| Hexyl acetate | Green, fruity, sweet, fatty, fresh, apple and pear | 670 | 73 | 5.7 | >DL | |
| Isobutyl acetate | Sweet, fruity, ethereal with an apple banana nuance | 1600 | 500 | 14 | 6.5 | >DL |
| 2-methyl butyl acetate | Sweet, banana, fruity, ripe, estery and tropical with a juicy, fruit-like note | 78 | 0.84 | >DL | ||
| 3-methyl butyl acetate | Sweet, banana, fruity with a ripe estery nuance | 160 | 2000 | 40 | 4.9 | 6.8 |
| 2-phenylethyl acetate | Sweet, honey, floral rosy, with a slight yeasty honey note with a cocoa and balsamic nuance | 1800 | 200 | 354 | 372 | 2.7 |
| Ethyl propanoate | Sweet, ethereal, rummy, grape, winey and fermented with an eggnog nuance | 1800 | 51 | 240 | 30 | |
| Ethyl isobutyrate | Citrus, fruity, sweet | 45 | 16 | 1 | ||
| Ethyl hexanoate | Sweet, fruity, pineapple, waxy, fatty and estery with a green banana nuance | 80 | 200 | 5 | >DL | 10 |
| Ethyl octanoate | Waxy, sweet, musty, pineapple and fruity with a creamy, dairy nuance | 580 | 1000 | 2 | >DL | 3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Youssef, C.; Bonnarme, P.; Fraud, S.; Péron, A.-C.; Helinck, S.; Landaud, S. Sensory Improvement of a Pea Protein-Based Product Using Microbial Co-Cultures of Lactic Acid Bacteria and Yeasts. Foods 2020, 9, 349. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9030349
El Youssef C, Bonnarme P, Fraud S, Péron A-C, Helinck S, Landaud S. Sensory Improvement of a Pea Protein-Based Product Using Microbial Co-Cultures of Lactic Acid Bacteria and Yeasts. Foods. 2020; 9(3):349. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9030349
Chicago/Turabian StyleEl Youssef, Cynthia, Pascal Bonnarme, Sébastien Fraud, Anne-Claire Péron, Sandra Helinck, and Sophie Landaud. 2020. "Sensory Improvement of a Pea Protein-Based Product Using Microbial Co-Cultures of Lactic Acid Bacteria and Yeasts" Foods 9, no. 3: 349. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9030349