Carcass Yields and Physiochemical Meat Quality of Semi-extensive and Intensively Farmed Impala (Aepyceros melampus)

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Locations, Production Systems, Animals and Design
2.2. Culling, Carcass Processing and Yields
2.3. Physiochemical Meat Quality Evaluation
2.4. Statistical Analysis
3. Results
3.1. Carcass and Offal Yields
3.2. Physiochemical Meat Quality
4. Discussion
4.1. Carcass and Offal Yields
4.2. Physiochemical Meat Quality
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Taylor, A.; Lindsey, P.; Davies-Mostert, H. An Assessment of the Economic, Social and Conservation Value of the Wildlife Ranching Industry and Its Potential to Support the Green Economy in South Africa; The Endangered Wildlife Trust: Johannesburg, South Africa, 2016; Available online: http://www.sagreenfund.org.za/wordpress/wp-content/uploads/2016/04/EWT-RESEARCH-REPORT.pdf (accessed on 25 February 2020).
- Child, B.A.; Musengezi, J.; Parent, G.D.; Child, G.F.T. The economics and institutional economics of wildlife on private land in Africa. Pastoralism 2012, 2, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Oberem, P.; Oberem, P. The New Game Rancher; Briza Publications: Pretoria, South Africa, 2016. [Google Scholar]
- Snijders, D. Wild property and its boundaries—On wildlife policy and rural consequences in South Africa. J. Peasant Stud. 2012, 39, 503–520. [Google Scholar] [CrossRef] [Green Version]
- Berry, M.P.S. A comparison of different wildlife production enterprises in the Northern Cape Province, South Africa. S. Afr. J. Wildl. Res. 1986, 16, 124–128. Available online: https://journals.co.za/content/wild/16/4/AJA03794369_3579 (accessed on 25 February 2020).
- Hoffman, L.C. The yield and carcass chemical composition of impala (Aepyceros melampus), a southern African antelope species. J. Sci. 2000, 80, 752–756. [Google Scholar] [CrossRef]
- Van Zyl, L.; Ferreira, A.V. Physical and chemical carcass composition of springbok (Antidorcas marsupialis), blesbok (Damaliscus dorcas phillipsi) and impala (Aepyceros melampus). Small Rumin. Res. 2004, 53, 103–109. [Google Scholar] [CrossRef]
- Von La Chevallerie, M.; Van Zyl, J.H.M. Some effects of shooting on losses of meat and meat quality in springbok and impala. S. Afr. J. Anim. Sci. 1971, 1, 113–116. Available online: https://www.sasas.co.za/journals/some-effects-of-shooting-on-losses-of-meat-and-meat-quality-in-springbok-and-impala/ (accessed on 25 February 2020).
- Cooper, S.M.; Van der Merwe, M. Game ranching for meat production in marginal African agricultural lands. J. Arid Land Stud. 2014, 24, 249–252. Available online: http://nodaiweb.university.jp/desert/pdf11/249-252%20Cooper_r.pdf (accessed on 25 February 2020).
- Huntley, B.J. Carcass composition of mature male blesbok and kudu. S. Afr. J. Anim. Sci. 1971, 1, 125–128. Available online: https://www.ajol.info/index.php/sajas/article/viewFile/140499/130244 (accessed on 25 February 2020).
- McCrindle, C.M.E.; Siegmund-Schultze, M.; Heeb, A.W.; Zárate, A.V.; Ramrajh, S. Improving food security and safety through use of edible by-products from wild game. J. Arid Land Stud. 2013, 15, 1245–1257. [Google Scholar] [CrossRef] [Green Version]
- Fairall, N. Production parameters of the impala, Aepyceros melampus. S. Afr. J. Anim. Sci. 1983, 13, 176–179. Available online: https://www.sasas.co.za/journals/production-parameters-of-the-impala-aepyceros-melampus/ (accessed on 25 February 2020).
- Hoffman, L.C.; Kritzinger, B.; Ferreira, A.V. The effects of region and gender on the fatty acid, amino acid, mineral, myoglobin and collagen contents of impala (Aepyceros melampus) meat. Meat Sci. 2005, 69, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Selier, S.A.J.; Hoffman, L.C.; Castley, G. A conservation assessment of Aepyceros melampus melampus. In The Red List of Mammals of South Africa, Swaziland and Lesotho; Child, M.F., Roxburgh, L., Do Linh San, E., Raimondo, D., Davies-Mostert, H.T., Eds.; South African National Biodiversity Institute and Endangered Wildlife Trust: Midrand, South Africa, 2016. [Google Scholar]
- Skinner, J.D.; Monro, R.H.; Zimmermann, I. Comparative food intake and growth of cattle and impala on mixed tree savanna. S. Afr. J. Wildl. Res. 1984, 14, 1–9. Available online: https://hdl.handle.net/10520/AJA03794369_1699 (accessed on 25 February 2020).
- Du Plessis, I.; Van Der Waal, C.C.; Van Wyk, R.R.; Webb, E.C.; Kritzinger, B.; Van den Berg, J. Regional differences in growth parameters between two impala populations. S. Afr. J. Anim. Sci. 2006, 36, 90–94. Available online: https://www.sasas.co.za/journals/regional-differences-in-growth-parameters-between-two-impala-populations/ (accessed on 25 February 2020).
- Kohn, T.A.; Kritzinger, B.; Hoffman, L.C.; Myburgh, K.H. Characteristics of impala (Aepyceros melampus) skeletal muscles. Meat Sci. 2005, 69, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, L.C.; Mostert, A.C.; Kidd, M.; Laubscher, L.L. Meat quality of kudu (Tragelaphus strepsiceros) and impala (Aepyceros melampus): Carcass yield, physical quality and chemical composition of kudu and impala Longissimus dorsi muscle as affected by gender and age. Meat Sci. 2009, 83, 788–795. [Google Scholar] [CrossRef]
- Hoffman, L.C.; Mostert, A.C.; Laubscher, L.L. Meat quality of kudu (Tragelaphus strepsiceros) and impala (Aepyceros melampus): The effect of gender and age on the fatty acid profile, cholesterol content and sensory characteristics of kudu and impala meat. Meat Sci. 2009, 83, 737–743. [Google Scholar] [CrossRef]
- Mucina, L.; Rutherford, M.C. The Vegetation of South Africa, Lesotho and Swaziland; South African National Biodiversity Institute: Pretoria, South Africa, 2006. [Google Scholar]
- Association of Official Analytical Chemist International. Official Method of Analysis, 17th ed.; Association of Official Analytical Chemists Inc.: Arlington, TX, USA, 2002. [Google Scholar]
- Furstenburg, D. Impala (Aepyceros melampus). In The New Game Rancher; Oberem, P., Oberem, P., Eds.; Briza Publications: Pretoria, South Africa, 2016; pp. 217–225. [Google Scholar]
- Van Schalkwyk, D.L.; Hoffman, L.C. Guidelines for the Harvesting and Processing of Wild Game in Namibia 2016; Ministry of Environment & Tourism: Windhoek, Namibia, 2016.
- Commission Internationale de l’Eclairage. Recommendations on Uniform Color Spaces-Color Difference Equations, Psychometric Color Terms; CIE: Paris, France, 1976. [Google Scholar]
- Honikel, K.O. Reference methods for the assessment of physical characteristics of meat. Meat Sci. 1998, 49, 447–457. [Google Scholar] [CrossRef]
- Needham, T.; Bureš, D.; Kotrba, R.; Hoffman, L.C. Influence of sex on the muscle yield and physiochemical characteristics of fresh meat harvested from common eland (Taurotragus oryx). Meat Sci. 2019, 152, 41–48. [Google Scholar] [CrossRef]
- Lee, C.M.; Trevino, B.; Chaiyawat, M. A simple and rapid solvent extraction method for determining total lipids in fish tissue. J. AOAC Int. 1996, 79, 487–492. [Google Scholar] [CrossRef] [Green Version]
- Fairall, N.; Braack, H.H. Growth and development of the impala Aepyceros melampus. Koedoe 1979, 19, 83–88. [Google Scholar] [CrossRef] [Green Version]
- Furstenburg, D. The impala. In Intensive Wildlife Production in Southern Africa; Bothma, J., Du, P., Van Rooyen, N., Eds.; Van Schaik Publishers: Pretoria, South Africa, 2005; pp. 187–202. [Google Scholar]
- Anderson, I.G. Mass and body measurements of impala, Aepyceros melampus, from a game ranch. S. Afr. J. Wildl. Res. 1982, 12, 76–78. Available online: https://hdl.handle.net/10520/AJA03794369_3603 (accessed on 25 February 2020).
- Hoffman, L.C.; Kritzinger, B.; Ferreira, A.V. The effects of sex and region on the carcass yield and m longissimus lumborum proximate composition of impala. J. Sci. Food Agric. 2005, 85, 391–398. [Google Scholar] [CrossRef]
- Muchenje, V.; Dzama, K.; Chimonyo, M.; Raats, J.G.; Strydom, P.E. Tick susceptibility and its effects on growth performance and carcass characteristics of Nguni, Bonsmara and Angus steers raised on natural pasture. Animal 2008, 2, 298–304. [Google Scholar] [CrossRef] [Green Version]
- Cloete, J.J.E.; Hoffman, L.C.; Cloete, S.W.P.; Fourie, J.E. A comparison between the body composition, carcass characteristics and retail cuts of South African Mutton Merino and Dormer sheep. S. Afr. J. Anim. Sci. 2004, 34, 44–51. [Google Scholar] [CrossRef]
- Hoffman, L.C. Meat quality attributes of night-cropped impala (Aepyceros melampus). S. Afr. J. Anim. Sci. 2000, 30, 133–137. [Google Scholar] [CrossRef]
- Lewis, A.R.; Pinchin, A.M.; Kestin, S.C. Welfare implications of the night shooting of wild impala (Aepyceros melampus). Anim. Welf. 1997, 6, 123–131. [Google Scholar]
- Honikel, K.O. Conversion of muscle to meat. In Encyclopedia of Meat Sciences; Academic Press: Oxford, UK, 2004; pp. 314–318. [Google Scholar]
- Immonen, K.; Ruusunen, M.; Puolanne, E. Some effects of residual glycogen concentration on the physical and sensory quality of normal pH beef. Meat Sci. 2000, 55, 33–38. [Google Scholar] [CrossRef]
- Viljoen, H.F.; De Kock, H.L.; Webb, E.C. Consumer acceptability of dark, firm and dry (DFD) and normal pH beef steaks. Meat Sci. 2002, 61, 181–185. [Google Scholar] [CrossRef]
- Troy, D.J.; Kerry, J.P. Consumer perception and the role of science in the meat industry. Meat Sci. 2010, 86, 214–226. [Google Scholar] [CrossRef]
- Kritzinger, B.; Hoffman, L.C.; Ferreira, A.V. A comparison between the effects of two cropping methods on the meat quality of impala (Aepyceros melampus). S. Afr. J. Anim. Sci. 2003, 33, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Macdougall, D.B.; Shaw, B.G.; Nute, G.R.; Rhodes, D.N. Effect of pre-slaughter handling on the quality and microbiology of venison from farmed young red deer. J. Sci. Food Agric. 1979, 30, 1160–1167. [Google Scholar] [CrossRef]
- Neethling, N.E.; Sigge, G.O.; Hoffman, L.C.; Suman, S.P. Color stability of fallow deer (Dama dama) Infraspinatus, Longissimus thoracis et lumborum, and Biceps femoris muscles during refrigerated storage. Meat Muscle Biol. 2018, 2, 147–161. [Google Scholar] [CrossRef] [Green Version]
- Shange, N.; Gouws, P.; Hoffman, L.C. Changes in pH, color and the microbiology of black wildebeest (Connochaetes gnou) longissimus thoracis et lumborum (LTL) muscle with normal and high (DFD) muscle pH. Meat Sci. 2019, 147, 13–19. [Google Scholar] [CrossRef]
- Sebsibe, A. Sheep and goat meat characteristics and quality. In Meat Production and Quality, Sheep and Goat Production Handbook for Ethiopia; Yami, A., Markel, R.C., Eds.; Brana Publishing House: Addis Ababa, Ethiopia, 2008; pp. 325–329. [Google Scholar]
- Hoffman, L.C.; Van Schalkwyk, S.; Muller, N.M. Physical and chemical properties of male and female mountain reedbuck (Redunca fulvorufula) meat. S. Afr. J. Wildl. Res. 2008, 38, 11–16. [Google Scholar] [CrossRef]
- Hoffman, L.C.; Kroucamp, M.; Manley, M. Meat quality characteristics of springbok (Antidorcas marsupialis). 2: Chemical composition of springbok meat as influenced by age, gender and production region. Meat Sci. 2007, 76, 762–767. [Google Scholar] [CrossRef] [PubMed]
- Destefanis, G.; Brugiapaglia, A.; Barge, M.T.; Dal Molin, E. Relationship between beef consumer tenderness perception and Warner–Bratzler shear force. Meat Sci. 2008, 78, 153–156. [Google Scholar] [CrossRef]
- Huff-Lonergan, E.; Lonergan, S.M. Mechanisms of water-holding capacity of meat: The role of post-mortem biochemical and structural changes. Meat Sci. 2005, 71, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Listrat, A.; Lebret, B.; Louveau, I.; Astruc, T.; Bonnet, M.; Lefaucheur, L.; Picard, B.; Bugeon, J. How muscle structure and composition influence meat and flesh quality. Sci. World J. 2016, 1–14. [Google Scholar] [CrossRef] [Green Version]
- North, M.K.; Hoffman, L.C. Changes in springbok (Antidorcas marsupialis) Longissimus thoracis et lumborum muscle during conditioning as assessed by a trained sensory panel. Meat Sci. 2015, 108, 1–8. [Google Scholar] [CrossRef]
- Neethling, J.; Hoffman, L.C.; Britz, T.J. Impact of season on the chemical composition of male and female blesbok (Damaliscus pygargus phillipsi) muscles. J. Sci. Food Agric. 2014, 94, 424–431. [Google Scholar] [CrossRef]
- Schack, W.; Bergh, T.; Du Toit, J.G. Meat production. In Game Ranch Management, 6th ed.; Bothma, J., Du, P., Du Toit, J.G., Eds.; Van Schaik Publishers: Pretoria, South Africa, 2016; pp. 759–788. [Google Scholar]
- Geldenhuys, G.; Hoffman, L.C.; Muller, M. Sensory profiling of Egyptian goose (Alopochen aegyptiaca) meat. Food Res. Int. 2014, 64, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Purslow, P.P. Intramuscular connective tissue and its role in meat quality. Meat Sci. 2005, 70, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Tshabalala, P.A.; Strydom, P.E.; Webb, E.C.; De Kock, H.L. Meat quality of designated South African indigenous goat and sheep breeds. Meat Sci. 2003, 65, 563–570. [Google Scholar] [CrossRef]


{kind=link}
| Parameters | Sex | Production System | ||||
|---|---|---|---|---|---|---|
| Female (n = 11) | Male (n = 12) | p | Intensive (n = 12) | Semi-Extensive (n = 12) | p | |
| Undressed carcass (kg) | 37.8 ± 1.30 | 36.4 ± 1.30 | 0.451 | 37.9 ± 0.92 | 36.4 ± 0.96 | 0.287 |
| Warm dressed carcass (kg) | 21.0 ± 0.82 | 21.6 ± 0.82 | 0.639 | 21.9 ± 0.65 | 21.3 ± 0.68 | 0.500 |
| Warm dressing percentage | 55.6 ± 0.76 | 59.1 ± 0.76 | 0.004 | 57.9 ± 0.51 | 58.4 ± 0.54 | 0.505 |
| Cold dressed carcass (kg) | 20.5 ± 0.82 | 20.9 ± 0.82 | 0.727 | 20.9 ± 0.65 | 20.7 ± 0.67 | 0.789 |
| Total offal (%) | 41.6 ± 0.84 | 39.2 ± 0.84 | 0.05 | 39.4 ± 0.42 | 40.4 ± 0.43 | 0.130 |
| Total external offal (%) | 12.5 ± 0.28 | 14.8 ± 0.28 | <0.001 | 14.6 ± 0.24 | 14.7 ± 0.25 | 0.638 |
| Total internal offal (%) | 29.1 ± 0.88 | 24.3 ± 0.88 | <0.001 | 24.9 ± 0.40 | 25.6 ± 0.39 | 0.156 |
| Head (%) | 5.1 ± 0.12 | 6.5 ± 0.12 | <0.001 | 6.64 ± 0.07 | 6.56 ± 0.08 | 0.506 |
| Feet (%) | 2.8 ± 0.08 | 3.3 ± 0.08 | <0.001 | 3.3 ± 0.12 | 3.2 ± 0.13 | 0.817 |
| Skin (%) | 4.6 ± 0.20 | 5.0 ± 0.20 | 0.192 | 4.6 ± 0.18 | 4.9 ± 0.19 | 0.294 |
| Heart (%) | 0.7 ± 0.01 | 0.7 ± 0.01 | 0.139 | 0.7 ± 0.02 | 0.7 ± 0.02 | 0.111 |
| Lungs & trachea (%) | 1.9 ± 0.15 | 2.1 ± 0.15 | 0.312 | 1.8 ± 0.11 | 2.1 ± 0.12 | 0.05 |
| Liver (%) | 1.6 ± 0.05 | 1.6 ± 0.05 | 0.451 | 1.4 ± 0.05 | 1.6 ± 0.05 | 0.059 |
| Kidneys (%) | 0.3 ± 0.01 | 0.3 ± 0.01 | 0.072 | 0.31 ± 0.01 | 0.27± 0.01 | 0.009 |
| Spleen (%) | 0.4 ± 0.04 | 0.5 ± 0.04 | 0.018 | 0.47 ± 0.02 | 0.53 ± 0.02 | 0.106 |
| GIT (%) | 24.3 ± 0.82 | 18.9 ± 0.82 | <0.001 | 19.9 ± 0.47 | 20.5 ± 0.49 | 0.416 |
| Muscle | Sex | p | |
|---|---|---|---|
| Female (n = 11) | Male (n = 12) | ||
| LTL | 855.4 ± 34.41 | 850.1 ± 34.41 | 0.916 |
| BF | 610.8 ± 27.38 | 602.2 ± 27.38 | 0.826 |
| SM | 638.9 ± 30.20 | 644.0 ± 31.68 | 0.910 |
| ST | 182.1 ± 7.28 | 183.0 ± 7.28 | 0.937 |
| IS | 164.6 ± 7.73 | 178.4 ± 7.73 | 0.222 |
| SS | 139.8 ± 6.78 | 154.0 ± 6.78 | 0.155 |
| Parameter | Sex | Muscle | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Female (n = 11) | Male (n = 12) | p | LTL | BF | SM | ST | IS | SS | p | |
| pHu | 5.6 ± 0.05 | 5.8 ± 0.05 | 0.021 | 5.6 ± 0.03 c | 5.7 ± 0.03 c | 5.6 ± 0.03 c | 5.7 ± 0.03 c | 5.8 ± 0.03 b | 5.9 ± 0.03 a | <0.001 |
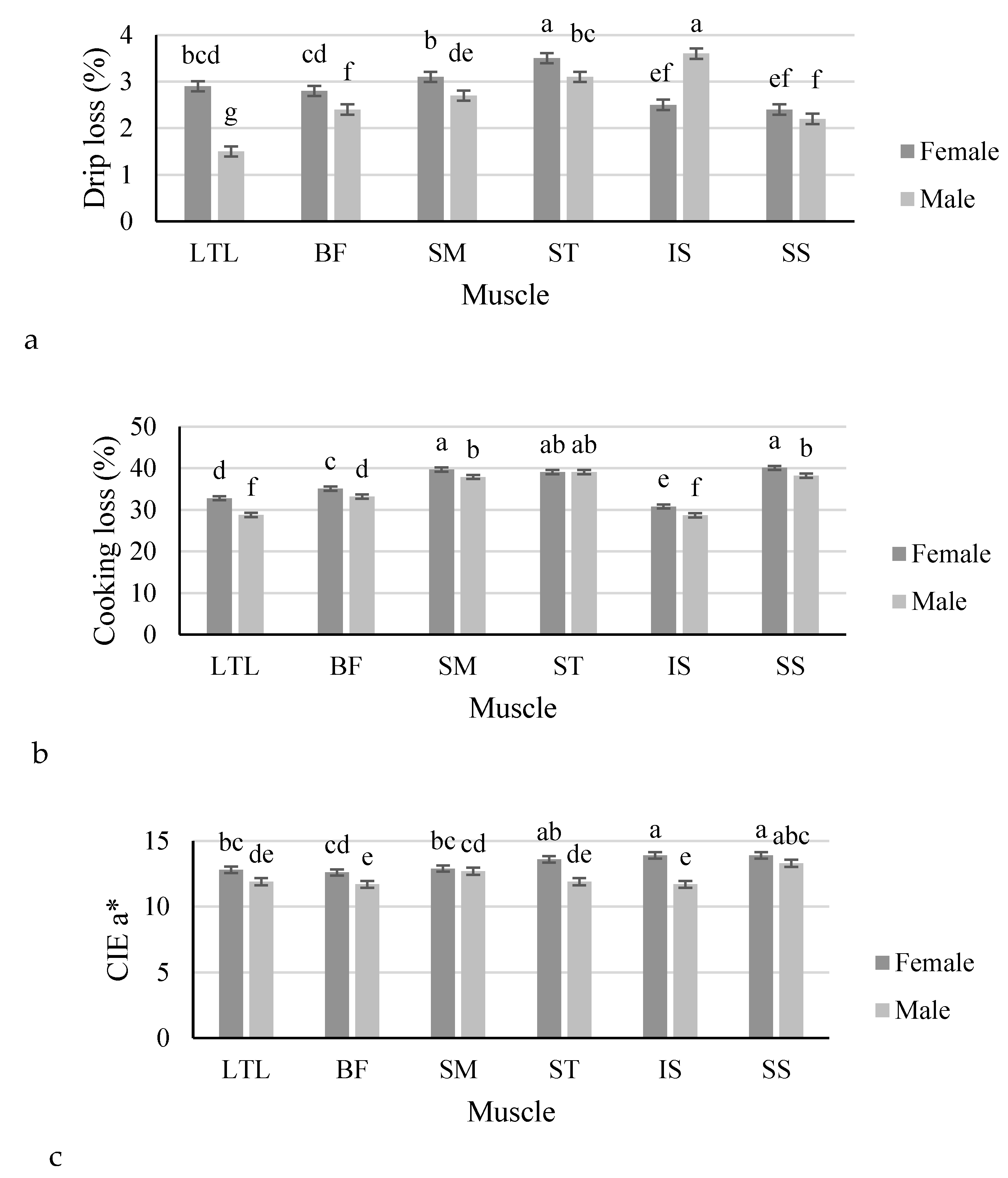
| Drip loss (%) | 2.9 ± 0.08 | 2.6 ± 0.08 | 0.013 | 2.2 ± 0.08 d | 2.6 ± 0.08 c | 2.9 ± 0.08 b | 3.3 ± 0.09 a | 3.0 ± 0.08 b | 2.3 ± 0.08 d | <0.001 |
| Cooking loss (%) | 36.3 ± 0.57 | 34.3 ± 0.59 | 0.028 | 30.8 ± 0.36 c | 34.1 ± 0.35 b | 38.8 ± 0.35 a | 39.1 ± 0.36 a | 29.8 ± 0.37 d | 39.2 ± 0.35 a | <0.001 |
| Shear force (N) | 28.8 ± 1.08 | 23.2 ± 1.07 | 0.002 | 25.5 ± 1.10 b | 30.1 ± 1.10 a | 31.7 ± 1.13 a | 25.7 ± 1.10 b | 19.2 ± 1.10 c | 23.6 ± 1.10 b | <0.001 |
| Color: | ||||||||||
| L* | 33.5 ± 0.77 | 34.8 ± 0.77 | 0.247 | 31.4 ± 0.41 d | 34.1 ± 0.41 b | 32.8 ± 0.41 c | 36.8 ± 0.41 a | 36.1 ± 0.41 a | 33.9 ± 0.41 bc | <0.001 |
| a* | 13.3 ± 0.27 | 12.2 ± 0.27 | 0.009 | 12.3 ± 0.21 bc | 12.1 ± 0.20 c | 12.8 ± 0.20 b | 12.7 ± 0.20 b | 12.8 ± 0.20 b | 13.6 ± 0.20 a | <0.001 |
| b* | 8.4 ± 0.47 | 7.7 ± 0.47 | 0.280 | 7.3 ± 0.28 d | 7.6 ± 0.27 d | 8.1 ± 0.27 bc | 8.8 ± 0.27 a | 8.6 ± 0.27 ab | 8.1 ± 0.27 abc | 0.001 |
| Chroma | 15.8 ± 0.45 | 14.5 ± 0.45 | 0.05 | 14.5 ± 0.29 bc | 14.4 ± 0.28 c | 15.2 ± 0.28 ab | 15.6 ± 0.28 a | 15.5 ± 0.28 a | 15.9 ± 0.28 a | 0.001 |
| Hue-angle | 32.1 ± 1.09 | 32.0 ± 1.10 | 0.965 | 29.6 ± 0.77 b | 31.6 ± 0.75 b | 31.7 ± 0.75 b | 34.6 ± 0.75 a | 33.8 ± 0.75 a | 30.9 ± 0.75 b | <0.001 |
| Parameter | Production System | p | |
|---|---|---|---|
| Intensive (n = 12) | Semi-Extensive (n = 12) | ||
| pHu | 5.8 ± 0.04 | 5.6 ± 0.05 | 0.012 |
| Drip loss (%) | 2.2 ± 0.13 | 1.5 ± 0.14 | 0.006 |
| Cooking loss (%) | 36.8 ± 0.53 | 29.5 ± 0.57 | <0.001 |
| Shear force (N) | 39.3 ± 1.78 | 22.4 ± 1.85 | <0.001 |
| Color | |||
| L* | 30.9 ± 0.77 | 32.2 ± 0.81 | 0.272 |
| a* | 11.4 ± 0.37 | 12.2 ± 0.38 | 0.145 |
| b* | 6.0 ± 0.57 | 7.1 ± 0.59 | 0.186 |
| Chroma | 13.1 ± 0.52 | 14.2 ± 0.54 | 0.121 |
| Hue-angle | 27.8 ± 1.73 | 29.4 ± 1.81 | 0.542 |
| Proximate composition (g/100 g) | |||
| Moisture | 75.3 ± 0.13 | 75.7 ± 0.13 | 0.191 |
| Protein | 22.7 ± 0.12 | 22.0 ± 0.12 | 0.0004 |
| Fat | 2.0 ± 0.06 | 1.8 ± 0.06 | 0.019 |
| Ash | 1.21 ± 0.01 | 1.19 ± 0.01 | 0.117 |
| Parameter (g/100 g) | Muscle | Sex | p | |
|---|---|---|---|---|
| Female (n = 11) | Male (n = 12) | |||
| Moisture | LTL | 76.0 ± 0.15 cd | 76.0 ± 0.13 c | 0.002 |
| BF | 75.2 ± 0.13 fg | 75.6 ± 0.13 de | ||
| SM | 74.8 ± 0.13 g | 75.3 ± 0.13 ef | ||
| ST | 76.1 ± 0.13 c | 76.0 ± 0.13 c | ||
| IS | 76.3 ± 0.13 bc | 76.5 ± 0.13 b | ||
| SS | 76.1c ± 0.13 | 77.0 ± 0.13 a | ||
| Protein | LTL | 22.6 ± 0.19 bc | 21.6 ± 0.18 ef | <0.001 |
| BF | 23.1 ± 0.18 ab | 22.7 ± 0.18 bc | ||
| SM | 23.5 ± 0.18 a | 22.3 ± 0.18 cd | ||
| ST | 22.6 ± 0.18 cd | 22.8 ± 0.18 bc | ||
| IS | 21.6 ± 0.18 ef | 21.4 ± 0.18 f | ||
| SS | 21.5 ± 0.12 c | 20.7 ± 0.19 g | ||
| Intramuscular fat | LTL | 1.2 ± 0.08 f | 1.9 ± 0.08 b | <0.001 |
| BF | 1.6 ± 0.08 c | 1.3 ± 0.08 def | ||
| SM | 1.4 ± 0.08 cd | 2.2 ± 0.08 a | ||
| ST | 1.1 ± 0.08 f | 1.4 ± 0.08 cde | ||
| IS | 1.9 ± 0.08 b | 2.1 ± 0.09 ab | ||
| SS | 1.2 ± 0.08 ef | 1.8 ± 0.09 b | ||
| Ash | LTL | 1.2 ± 0.02 b | 1.4 ± 0.03 a | 0.004 |
| BF | 1.2 ± 0.02 b | 1.2 ± 0.02 bc | ||
| SM | 1.2 ± 0.02 bcd | 1.2 ± 0.02 bcd | ||
| ST | 1.2 ± 0.02 b | 1.2 ± 0.02 cde | ||
| IS | 1.1 ± 0.02 e | 1.1 ± 0.02 de | ||
| SS | 1.1 ± 0.02 cde | 1.1 ± 0.02 cde | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Needham, T.; Engels, R.A.; Bureš, D.; Kotrba, R.; van Rensburg, B.J.; Hoffman, L.C. Carcass Yields and Physiochemical Meat Quality of Semi-extensive and Intensively Farmed Impala (Aepyceros melampus). Foods 2020, 9, 418. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9040418
Needham T, Engels RA, Bureš D, Kotrba R, van Rensburg BJ, Hoffman LC. Carcass Yields and Physiochemical Meat Quality of Semi-extensive and Intensively Farmed Impala (Aepyceros melampus). Foods. 2020; 9(4):418. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9040418
Chicago/Turabian StyleNeedham, Tersia, Retha A. Engels, Daniel Bureš, Radim Kotrba, Berndt J. van Rensburg, and Louwrens C. Hoffman. 2020. "Carcass Yields and Physiochemical Meat Quality of Semi-extensive and Intensively Farmed Impala (Aepyceros melampus)" Foods 9, no. 4: 418. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9040418