Chemical Composition of Wild Fallow Deer (Dama Dama) Meat from South Africa: A Preliminary Evaluation †

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Harvesting and Slaughtering
2.2. Muscle Removal and Sample Preparation
2.3. Proximate Analysis
2.4. Minerals
2.5. Statistical Analysis
2.6. Comparative Data for Deer, Antelope, and Livestock
3. Results
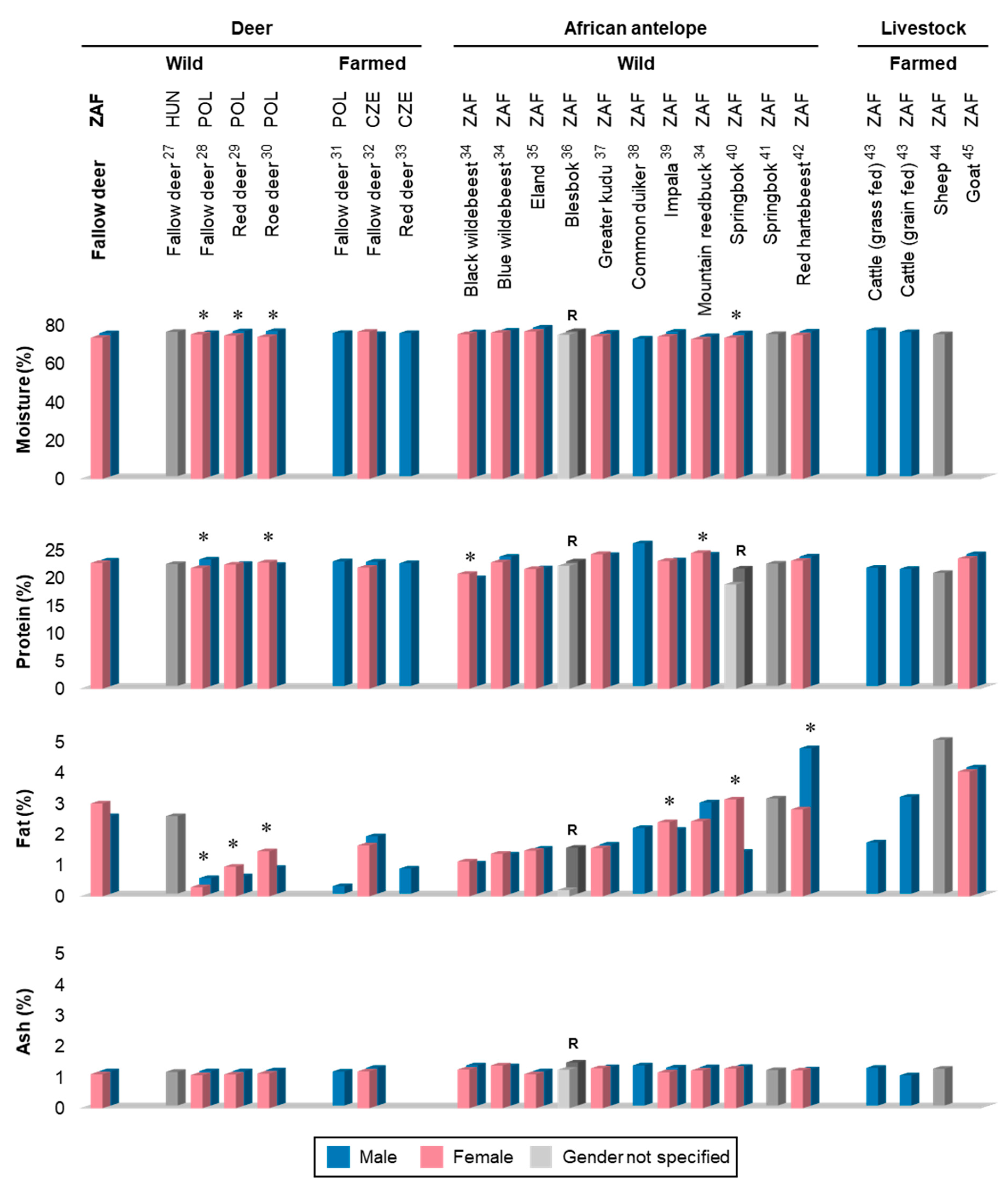
3.1. Proximate Composition
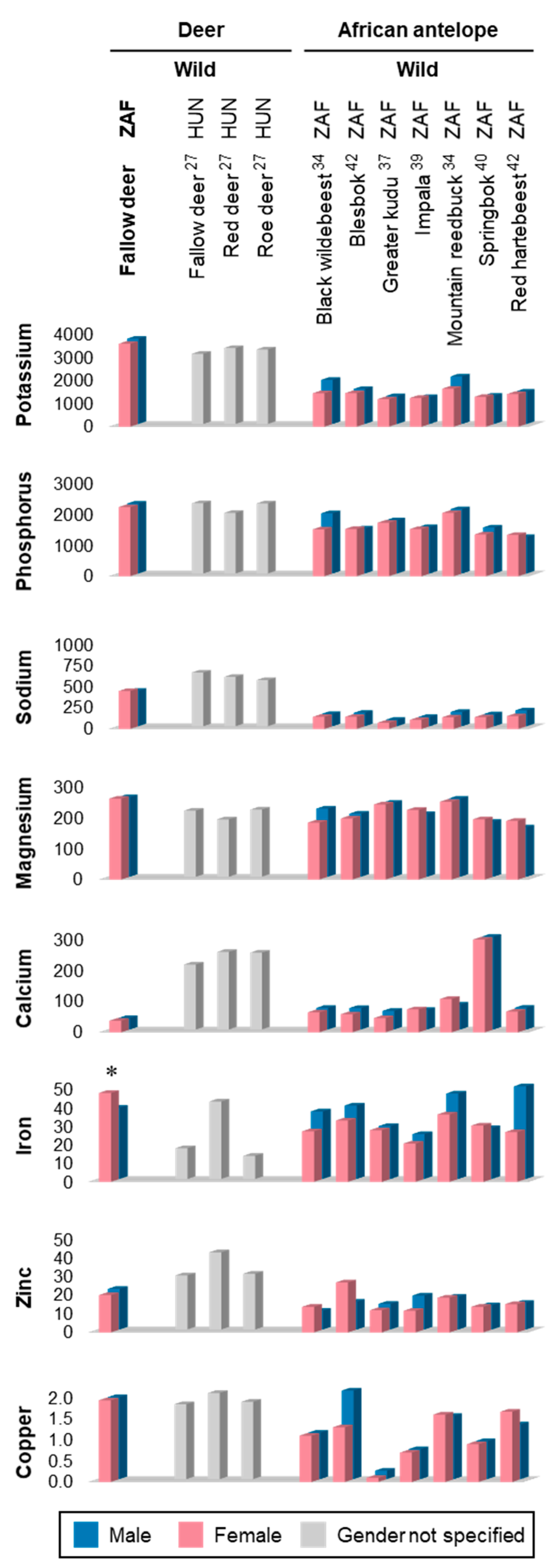
3.2. Mineral Composition
4. Discussion
4.1. Proximate Composition
4.2. Mineral Composition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Thornton, P.K. Livestock production: Recent trends, future prospects. Philos. Trans. R. Soc. B: Biol. Sci. 2010, 365, 2853–2867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esterhuizen, D. The South African Meat Market; GAIN Report; USDA Foreign Agricultural Service: Washington, DC, USA, 2015. Available online: www.fas.usda.gov/data/south-africa-south-african-meat-market (accessed on 7 September 2019).
- Goldblatt, A. Agriculture: Facts & Trends, South Africa; World Wide Fund for Nature: Cape Town, South Africa, 2016. [Google Scholar]
- Von Bormann, T. Agri-Food Systems: Facts and Futures. How South Africa Can Produce 50% More by 2050; World Wide Fund for Nature: Cape Town, South Africa, 2019. [Google Scholar]
- Cooperative Governance and Traditional Affairs. Integrated Urban Development Framework; COGTA: Pretoria, South Africa, 2016.
- Schreiner, B.G.; Mungatana, E.D.; Baleta, H. Impacts of drought induced water shortages in South Africa: Economic Analysis (WRC Report No. 2604/1/18); Water Research Commission: Pretoria, South Africa, 2018. [Google Scholar]
- Shisana, O.; Labadarios, D.; Rehle, T.; Simbayi, L.; Zuma, K.; Dhansay, A.; Reddy, P.; Parker, W.; Hoosain, E.; Naidoo, P.; et al. South African National Health and Nutrition Examination Survey (SANHANES-1); HSRC Press: Cape Town, South Africa, 2013. [Google Scholar]
- Cawthorn, D.M.; Hoffman, L.C. The role of traditional and non-traditional meat animals in feeding a growing and evolving world. Anim. Front. 2014, 4, 6–12. [Google Scholar] [CrossRef] [Green Version]
- Cooper, S.M.; Van der Merwe, M. Game ranching for meat production in marginal African agricultural lands. J. Arid Land Stud. 2014, 24, 249–252. [Google Scholar]
- Van Hoven, W. Private game reserves in southern Africa. In Institutional Arrangements for Conservation, Development and Tourism in Eastern and Southern Africa: A Dynamic Perspective; van der Duim, R., Lamers, M., van Wijk, J., Eds.; Springer: Dordrecht, Germany, 2015; pp. 101–118. [Google Scholar]
- Department of Environmental Affairs. National Biodiversity Economy Strategy; Department of Environmental Affairs: Pretoria, South Africa, 2016. Available online: www.environment.gov.za/sites/default/files/reports/nationalbiodiversityeconomystrategy.pdf (accessed on 16 August 2019).
- Chapman, N.G.; Chapman, D.I. The distribution of fallow deer: A worldwide review. Mamm. Rev. 1980, 10, 61–138. [Google Scholar] [CrossRef]
- Bothma, J.P. The fallow deer: Dama dama. Game Hunt 2014, 20, 14–17. [Google Scholar]
- Hudson, R.J. Livestock diversification: Issues and trends. In Management of Agricultural, Forestry and Fisheries Enterprises; EOLSS Publications: Oxford, UK, 2009; pp. 361–374. [Google Scholar]
- Kudrnáčová, E.; Bartoň, L.; Bureš, D.; Hoffman, L.C. Carcass and meat characteristics from farm-raised and wild fallow deer (Dama dama) and red deer (Cervus elaphus): A review. Meat Sci. 2018, 141, 9–27. [Google Scholar] [CrossRef]
- Sebranek, J.G. Raw material composition analysis. In Encyclopedia of Meat Sciences; Devine, C., Dikeman, M., Eds.; Elsevier: Oxford, UK, 2014; pp. 321–328. [Google Scholar]
- Hocquette, J.F.; Gondret, F.; Baéza, E.; Médale, F.; Jurie, C.; Pethick, D.W. Intramuscular fat content in meat-producing animals: Development, genetic and nutritional control, and identification of putative markers. Animal 2010, 4, 303–319. [Google Scholar] [CrossRef] [Green Version]
- Ledger, H.P.; Sachs, R.; Smith, N.S. Wildlife and food production. World Rev. Anim. Prod. 1967, 3, 13–36. [Google Scholar]
- Parker, K.L.; Barboza, P.S.; Gillingham, M.P. Nutrition integrates environmental responses of ungulates. Funct. Ecol. 2009, 23, 57–69. [Google Scholar] [CrossRef]
- Hoffman, L.C. The yield and carcass chemical composition of impala (Aepyceros melampus), a southern African antelope species. J. Sci. Food. Agric. 2000, 80, 752–756. [Google Scholar] [CrossRef]
- Garel, M.; Forsyth, D.M.; Loison, A.; Dubray, D.; Jullien, J.M.; Tustin, K.G.; Maillard, J.; Gaillard, J.M. Age-related male reproductive effort in two mountain ungulates of contrasting sexual size dimorphism. Can. J. Zool. 2011, 89, 929–937. [Google Scholar] [CrossRef]
- Pearson, A.M.; Young, R.B. Muscle and Meat Biochemistry; Elsevier: Oxford, UK, 2012; pp. 235–261. [Google Scholar]
- Cawthorn, D.M.; Fitzhenry, L.B.; Muchenje, V.; Bureš, D.; Kotrba, R.; Hoffman, L.C. Physical quality attributes of male and female wild fallow deer (Dama dama) muscles. Meat Sci. 2018, 137, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Dharmani, A. Dama dama; Animal Diversity Web: Ann Arbor, MI, USA, 2000; Available online: https://animaldiversity.org/accounts/Dama_dama/ (accessed on 15 December 2019).
- Association of Official Analytical Chemists. Official Method of Analysis, 17th ed.; AOAC International: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Lee, C.M.; Trevino, B.; Chaiyawat, M. A simple and rapid solvent extraction method for determining total lipids in fish tissue. J. AOAC Int. 1995, 79, 487–492. [Google Scholar] [CrossRef] [Green Version]
- Zomborszky, Z.; Szentmihalyi, G.; Sarudi, I.; Horn, P.; Szabo, C.S. Nutrient composition of muscles in deer and boar. J. Food Sci. 1996, 61, 625–627. [Google Scholar] [CrossRef]
- Piaskowska, N.; Daszkiewicz, T.; Kubiak, D.; Janiszewski, P. The effect of gender on meat (Longissimus lumborum muscle) quality characteristics in the fallow deer Dama dama L. Ital. J. Anim. Sci. 2015, 14, 3845. [Google Scholar] [CrossRef]
- Daszkiewicz, T.; Janiszewski, P.; Wajda, S. Quality characteristics of meat from wild red deer (Cervus elaphus L.) hinds and stags. J. Muscle Foods 2009, 20, 428–448. [Google Scholar] [CrossRef]
- Daszkiewicz, T.; Kubiak, D.; Winarski, R.; Koba-Kowalczyk, M. The effect of gender on the quality of roe deer (Capreolus capreolus L.) meat. Small Rumin. Res. 2012, 103, 169–175. [Google Scholar] [CrossRef]
- Daszkiewicz, T.; Hnatyk, N.; Dąbrowski, D.; Janiszewski, P.; Gugołek, A.; Kubiak, D.; Śmiecińskaa, K.; Winarskia, R.; Koba-Kowalczyka, M. A comparison of the quality of the Longissimus lumborum muscle from wild and farm-raised fallow deer (Dama dama). Small Rumin. Res. 2015, 129, 77–83. [Google Scholar] [CrossRef]
- Švrčula, V.; Košinová, K.; Okrouhlá, M.; Chodová, D.; Hart, V. The effect of sex on meat quality of fallow deer (Dama dama) from the farm located in the Middle Bohemia. Ital. J. Anim. Sci. 2019, 18, 498–504. [Google Scholar] [CrossRef] [Green Version]
- Bureš, D.; Bartoň, L.; Kotrba, R.; Hakl, J. Quality attributes and composition of meat from red deer (Cervus elaphus), fallow deer (Dama dama) and Aberdeen Angus and Holstein cattle (Bos taurus). J. Sci. Food. Agric. 2015, 95, 2299–2306. [Google Scholar] [CrossRef]
- Van Schalkwyk, S. Meat Quality Characteristics of Three South African Game Species: Black Wildebeest (Connochaetes gnou), Blue Wildebeest (Connochaetes taurinus), Mountain Reedbuck (Redunca fulvorufula). Master’s Thesis, Stellenbosch University, Stellenbosch, South Africa, December 2004. [Google Scholar]
- Needham, T.; Laubser, J.G.; Kotrba, R.; Bureš, D.; Hoffman, L.C. Sex influence on muscle yield and physiochemical characteristics of common eland (Taurotragus oryx) meat. Meat Sci. 2019, 152, 41–48. [Google Scholar] [CrossRef]
- Hoffman, L.C.; Smit, K.; Muller, N. Chemical characteristics of blesbok (Damaliscus dorcas phillipsi) meat. J. Food Compos. Anal. 2008, 21, 315–319. [Google Scholar] [CrossRef]
- Mostert, R.; Hoffman, L.C. Effect of gender on the meat quality characteristics and chemical composition of kudu (Tragelaphus strepsiceros), an African antelope species. Food Chem. 2007, 104, 565–570. [Google Scholar] [CrossRef]
- Hoffman, L.C.; Ferreira, A.V. Chemical composition of two muscles of the common duiker (Sylvicapra grimmia). J. Sci. Food Agric. 2004, 84, 1541–1544. [Google Scholar] [CrossRef]
- Hoffman, L.C.; Mostert, A.C.; Kidd, M.; Laubscher, L.L. Meat quality of kudu (Tragelaphus strepsiceros) and impala (Aepyceros melampus): Carcass yield, physical quality and chemical composition of kudu and impala Longissimus dorsi muscle as affected by gender and age. Meat Sci. 2009, 83, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, L.C.; Kroucamp, M.; Manley, M. Meat quality characteristics of springbok (Antidorcas marsupialis). 2: Chemical composition of springbok meat as influenced by age, gender and production region. Meat Sci. 2007, 76, 762–767. [Google Scholar] [CrossRef]
- Neethling, J.; Muller, M.; van der Rijst, M.; Hoffman, L.C. Sensory quality and fatty acid content of springbok (Antidorcas marsupialis) meat: Influence of farm location and sex. J. Sci. Food Agric. 2018, 98, 2548–2556. [Google Scholar] [CrossRef] [PubMed]
- Smit, K. Meat Quality Characteristics of Blesbok (Damaliscus dorcas phillipsi) and Red Hartebeest (Alcelaphus buselaphus caama) Meat. Master’s Thesis, Stellenbosch University, Stellenbosch, South Africa, December 2004. [Google Scholar]
- Moholisa, E.; Strydom, P.E.; Hugo, A. The effect of beef production system on proximate composition and fatty acid profile of three beef muscles. S. Afr. J. Anim. Sci. 2018, 48, 295–306. [Google Scholar] [CrossRef] [Green Version]
- Sainsbury, J. Nutrient Content and Carcass Composition of South African Mutton with a Focus on Bioavailability of Selected Nutrients. Master’s Thesis, University of Pretoria, Pretoria, South Africa, December 2009. [Google Scholar]
- Simela, L. Meat Characteristics and Acceptability of Chevon from South African Indigenous Goats. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, June 2005. [Google Scholar]
- Keeton, J.T.; Ellerbeck, S.M.; Núñez de González, M.T. Chemical composition. In Encyclopedia of Meat Sciences; Devine, C., Dikeman, M., Eds.; Elsevier: Oxford, UK, 2014; pp. 235–243. [Google Scholar]
- Listrat, A.; Lebret, B.; Louveau, I.; Astruc, T.; Bonnet, M.; Lefaucheur, L.; Picard, B.; Bugeon, J. How muscle structure and composition influence meat and flesh quality. Sci. World J. 2016, 2016, 3182746. [Google Scholar] [CrossRef] [Green Version]
- Neethling, J.; Hoffman, L.C.; Britz, T.J. Impact of season on the chemical composition of male and female blesbok (Damaliscus pygargus phillipsi) muscles. J. Sci. Food Agric. 2014, 94, 424–431. [Google Scholar] [CrossRef]
- Van Heerden, A.M. Profiling the Meat Quality of Blue Wildebeest (Connochaetes taurinus). Master’s Thesis, University of Stellenbosch, Stellenbosch, South Africa, December 2018. [Google Scholar]
- Food and Agriculture Organization/World Health Organization. Protein and Amino Acid Requirements in Human Nutrition; FAO: Rome, Italy, 2007. [Google Scholar]
- Żochowska-Kujawska, J.; Kotowicz, M.; Sobczak, M.; Lachowicz, K.; Wójcik, J. Age-related changes in the carcass composition and meat quality of fallow deer (Dama dama L.). Meat Sci. 2019, 147, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Stanisz, M.; Skorupski, M.; Ślósarz, P.; Bykowska-Maciejewska, M.; Składanowska-Baryza, J.; Stańczak, Ł.; Krokowska-Paluszak, M.; Ludwiczak, A. The seasonal variation in the quality of venison from wild fallow deer (Dama dama)—A pilot study. Meat Sci. 2019, 150, 56–64. [Google Scholar] [CrossRef]
- Ortega-Barrales, P.; Fernández-de Córdova, M.L. Meat. In Handbook of Mineral Elements in Food; de la Guardia, M., Garrigues, S., Eds.; John Wiley & Sons: New York, NY, USA, 2015; pp. 599–619. [Google Scholar]
- Doornenbal, H.; Murray, A.C. Effects of age, breed, sex and muscle on certain mineral concentrations in cattle. J. Food Sci. 1982, 47, 55–58. [Google Scholar] [CrossRef]
- Lin, K.C.; Cross, H.R.; Johnson, H.K.; Breidenstein, B.C.; Randecker, V.; Field, R.A. Mineral composition of lamb carcasses from the United States and New Zealand. Meat Sci. 1988, 24, 47–59. [Google Scholar] [CrossRef]
- Otten, J.J.; Hellwig, J.P.; Meyers, L.D. Dietary Reference Intakes: The Essential Guide to Nutrient Requirements; National Academies Press: Washington, DC, USA, 2006; pp. 534–535. [Google Scholar]
- Taggart, M.A.; Reglero, M.M.; Camarero, P.R.; Mateo, R. Should legislation regarding maximum Pb and Cd levels in human food also cover large game meat? Environ. Int. 2011, 37, 18–25. [Google Scholar] [CrossRef]
- Department of Health. Regulations relating to maximum levels of metals in foodstuffs (R588/2018). Govern. Gazette 2018, 41704, 28–37. [Google Scholar]
- European Food Safety Authority. Scientific Opinion of the Panel on Food Additives, Flavourings, Processing Aids and Food Contact Materials on a request from European Commission on Safety of aluminium from dietary intake. EFSA J. 2008, 754, 1–34. [Google Scholar]



{kind=link}
{kind=link}
| Parameter (g/100 g) | Muscle | Total Group † | p-Value | Sex | p-Value | |
|---|---|---|---|---|---|---|
| (n = 12) | Muscle | Male (n = 6) | Female (n = 6) | Sex | ||
| Moisture | LTL | 73.8 a ± 0.22 | <0.0001 | 74.2 ± 0.36 | 73.4 ± 0.18 | 0.1187 |
| BF | 74.5 b ± 0.20 | 74.8 ± 0.29 | 74.2 ± 0.23 | |||
| SM | 73.3 a ± 0.21 | 73.4 ± 0.25 | 73.3 ± 0.36 | |||
| ST | 75.8 c ± 0.28 | 75.5 ± 0.36 | 76.0 ± 0.44 | |||
| IS | 76.1 c ± 0.16 | 76.2 ± 0.19 | 75.9 ± 0.25 | |||
| SS | 76.2 c ± 0.16 | 76.6 ± 0.11 | 75.9 ± 0.22 | |||
| Protein | LTL | 22.7 a ± 0.20 | <0.0001 | 22.6 ± 0.35 | 22.7 ± 0.23 | 0.9636 |
| BF | 21.9 b ± 0.24 | 21.8 ± 0.27 | 22.0 ± 0.43 | |||
| SM | 23.1 a ± 0.23 | 23.1 ± 0.30 | 23.0 ± 0.39 | |||
| ST | 20.9 cd ± 0.33 | 21.1 ± 0.50 | 20.6 ± 0.45 | |||
| IS | 21.3 c ± 0.15 | 21.3 ± 0.10 | 21.4 ± 0.30 | |||
| SS | 20.4 d ± 0.17 | 20.2 ± 0.24 | 20.6 ± 0.24 | |||
| Lipid | LTL | 2.8 a ± 0.16 | <0.0001 | 2.5 ± 0.09 | 3.0 ± 0.26 | 0.2967 |
| BF | 3.2 b ± 0.20 | 3.4 ± 0.36 | 3.1 ± 0.19 | |||
| SM | 2.9 ab ± 0.10 | 3.0 ± 0.19 | 2.9 ± 0.11 | |||
| ST | 2.7 a ± 0.12 | 2.4 ± 0.11 | 2.9 ± 0.17 | |||
| IS | 2.2 c ± 0.08 | 2.1 ± 0.06 | 2.3 ± 0.14 | |||
| SS | 2.8 a ± 0.15 | 2.8 ± 0.22 | 2.9 ± 0.22 | |||
| Ash | LTL | 1.1 a ± 0.02 | 0.0003 | 1.1 ± 0.02 | 1.1 ± 0.03 | 0.7267 |
| BF | 1.2 a ± 0.03 | 1.2 ± 0.05 | 1.2 ± 0.03 | |||
| SM | 1.4 bc ± 0.08 | 1.3 ± 0.06 | 1.5 ± 0.13 | |||
| ST | 1.5 c ± 0.07 | 1.4 ± 0.07 | 1.5 ± 0.12 | |||
| IS | 1.3 ab ± 0.05 | 1.3 ± 0.07 | 1.2 ± 0.08 | |||
| SS | 1.4 bc ± 0.06 | 1.4 ± 0.10 | 1.4 ± 0.09 | |||
| Mineral (mg/kg Meat) | Total Group † | Sex # | ||||||
|---|---|---|---|---|---|---|---|---|
| (n = 12) | p-Value | Male (n = 6) | Female (n = 6) | p-Value | ||||
| LTL | BF | Muscle | LTL | BF | LTL | BF | Sex | |
| Macro-minerals | ||||||||
| Potassium (K) | 3622.478 ± 33.775 * | 3743.060 ± 50.492 | 0.0004 | 3670.861 a ± 34.042 | 3852.473 b ± 45.258 | 3574.096 a ± 54.076 | 3633.647 a ± 66.186 | 0.0446 |
| Phosphorus (P) | 2245.846 ± 18.126 | 2301.909 ± 20.857 | 0.0208 | 2256.688 ± 26.579 | 2313.660 ± 26.152 | 2235.003 ± 26.308 | 2290.158 ± 34.278 | 0.5307 |
| Sodium (Na) | 435.319 ± 8.105 | 432.910 ± 9.621 | 0.7496 | 419.832 ± 11.758 | 426.394 ± 16.299 | 450.806 ± 7.408 | 439.427 ± 11.165 | 0.1841 |
| Magnesium (Mg) | 259.402 ± 1.353 | 272.992 ± 2.072 | 0.0001 | 256.985 ± 1.758 | 271.213 ± 2.261 | 261.818 ± 1.621 | 274.771 ± 3.536 | 0.1609 |
| Calcium (Ca) | 37.143 ± 0.720 | 34.810 ± 0.656 | 0.0168 | 36.809 ± 1.190 | 34.793 ± 1.109 | 37.478 ± 0.905 | 34.826 ± 0.817 | 0.7733 |
| Micro-minerals | ||||||||
| Iron (Fe) | 43.196 ± 1.954 | 38.294 ± 1.352 | 0.0023 | 38.414 ab ± 1.811 | 34.788 a ± 1.045 | 47.978 c ± 2.093 | 41.800 b ± 1.426 | 0.0019 |
| Zinc (Zn) | 20.844 ± 0.630 | 14.955 ± 0.499 | 0.0000 | 21.700 ± 0.843 | 14.521 ± 0.481 | 19.988 ± 0.862 | 15.390 ± 0.888 | 0.6644 |
| Silicon (Si) | 6.233 ± 0.316 | 5.506 ± 0.345 | 0.1599 | 6.522 ± 0.507 | 5.686 ± 0.669 | 5.944 ± 0.388 | 5.325 ± 0.254 | 0.3500 |
| Copper (Cu) | 1.942 ± 0.071 | 2.014 ± 0.055 | 0.1181 | 1.944 ± 0.132 | 2.008 ± 0.089 | 1.939 ± 0.069 | 2.020 ± 0.074 | 0.9762 |
| Manganese (Mn) | 0.201 ± 0.008 | 0.218 ± 0.008 | 0.0618 | 0.202 ± 0.012 | 0.203 ± 0.008 | 0.200 ± 0.013 | 0.233 ± 0.010 | 0.3038 |
| Selenium (Se) | 0.146 ± 0.007 | 0.139 ± 0.007 | 0.3232 | 0.139 ab ± 0.011 | 0.122 a ± 0.004 | 0.153 b ± 0.007 | 0.155 b ± 0.008 | 0.0208 |
| Chromium (Cr) | 0.058 ± 0.019 | 0.071 ± 0.028 | 0.7038 | 0.037 ± 0.015 | 0.104 ± 0.053 | 0.078 ± 0.034 | 0.037 ± 0.016 | 0.7043 |
| Cobalt (Co) | 0.003 ± 0.001 | 0.002 ± 0.000 | 0.3357 | 0.004 ± 0.002 | 0.001 ± 0.001 | 0.002 ± 0.000 | 0.002 ± 0.001 | 0.4963 |
| Molybdenum (Mo) | --- | --- | --- | --- | --- | --- | ||
| Undefined functions or environmental contaminants | ||||||||
| Aluminium (Al) | 5.763 ± 0.379 | 3.170 ± 0.55 | 0.0033 | 5.516 ± 0.585 | 2.696 ± 0.913 | 6.010 ± 0.514 | 3.644 ± 0.623 | 0.3111 |
| Lead (Pb) | 0.044 ± 0.029 | 0.009 ± 0.006 | 0.3958 | 0.011 ± 0.004 | 0.014 ± 0.011 | 0.020 ± 0.011 | 0.003 ± 0.003 | 0.3729 |
| Strontium (Sr) | 0.019 ± 0.003 | 0.014 ± 0.002 | 0.2003 | 0.024 ± 0.005 | 0.017 ± 0.004 | 0.014 ± 0.003 | 0.013 ± 0.002 | 0.0586 |
| Barium (Ba) | 0.014 ± 0.002 | 0.010 ± 0.001 | 0.0727 | 0.018 ± 0.003 | 0.010 ± 0.002 | 0.011 ± 0.002 | 0.009 ± 0.001 | 0.1048 |
| Antimony (Sb) | --- | --- | --- | --- | --- | --- | ||
| Arsenic (As) | --- | --- | --- | --- | --- | --- | ||
| Boron (B) | --- | --- | --- | --- | --- | --- | ||
| Cadmium (Cd) | --- | --- | --- | --- | --- | --- | ||
| Mercury (Hg) | --- | --- | --- | --- | --- | --- | ||
| Nickel (Ni) | --- | --- | --- | --- | --- | --- | ||
| Titanium (Ti) | --- | --- | --- | --- | --- | --- | ||
| Vanadium (V) | --- | --- | --- | --- | --- | --- | ||
| Element | LTL (mg/kg) | BF (mg/kg) | RDA/AI 56 (mg/day) | RDA/AI met by 100 g LTL | RDA/AI met by 100 g BF | Maximum Level |
|---|---|---|---|---|---|---|
| Essential Macro- and Micro-Minerals | ||||||
| Calcium (Ca) | 37.143 | 34.810 | 1000–1200 | 0.31–0.37% | 0.29–0.35% | --- |
| Copper (Cu) | 1.942 | 2.014 | 0.9 | 21.6% | 22.4% | --- |
| Iron (Fe) | 43.196 | 38.294 | 8–18 | 24–54% | 21.3–47.9% | --- |
| Magnesium (Mg) | 259.402 | 272.992 | 310–400 | 6.5–8.4% | 6.8–8.8% | --- |
| Manganese (Mn) | 0.201 | 0.218 | 1.8–2.3 | 0.9–1.1% | 0.9–1.1% | --- |
| Potassium (K) | 3622.478 | 3743.060 | 4700 | 7.7% | 8.0% | --- |
| Phosphorus (P) | 2245.846 | 2301.909 | 700 | 32.1% | 32.9% | --- |
| Sodium (Na) | 435.319 | 432.910 | 1500 | 2.9% | 2.9% | --- |
| Zinc (Zn) | 20.844 | 14.955 | 8–11 | 18.9–26.1% | 13.6–18.7% | --- |
| Elements with undefined functions or environmental contaminants | ||||||
| Lead (Pb) | 0.044 | 0.009 | --- | --- | --- | 0.1 mg/kg (meat) 58 |
| Aluminium (Al) | 5.763 | 3.170 | --- | --- | --- | 1 mg/kg bw/week 59 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cawthorn, D.-M.; Fitzhenry, L.B.; Kotrba, R.; Bureš, D.; Hoffman, L.C. Chemical Composition of Wild Fallow Deer (Dama Dama) Meat from South Africa: A Preliminary Evaluation. Foods 2020, 9, 598. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9050598
Cawthorn D-M, Fitzhenry LB, Kotrba R, Bureš D, Hoffman LC. Chemical Composition of Wild Fallow Deer (Dama Dama) Meat from South Africa: A Preliminary Evaluation. Foods. 2020; 9(5):598. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9050598
Chicago/Turabian StyleCawthorn, Donna-Mareè, Leon Brett Fitzhenry, Radim Kotrba, Daniel Bureš, and Louwrens C. Hoffman. 2020. "Chemical Composition of Wild Fallow Deer (Dama Dama) Meat from South Africa: A Preliminary Evaluation" Foods 9, no. 5: 598. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9050598