Optimization of Short-Term Hot-Water Treatment of Apples for Fruit Salad Production by Non-Invasive Chlorophyll-Fluorescence Imaging

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Short-Term Hot-Water Treatment
2.2. Spectral Analyses
2.3. Chlorophyll Fluorescence Imaging
2.4. Internal Quality Parameters
2.5. Statistical Analysis
3. Results
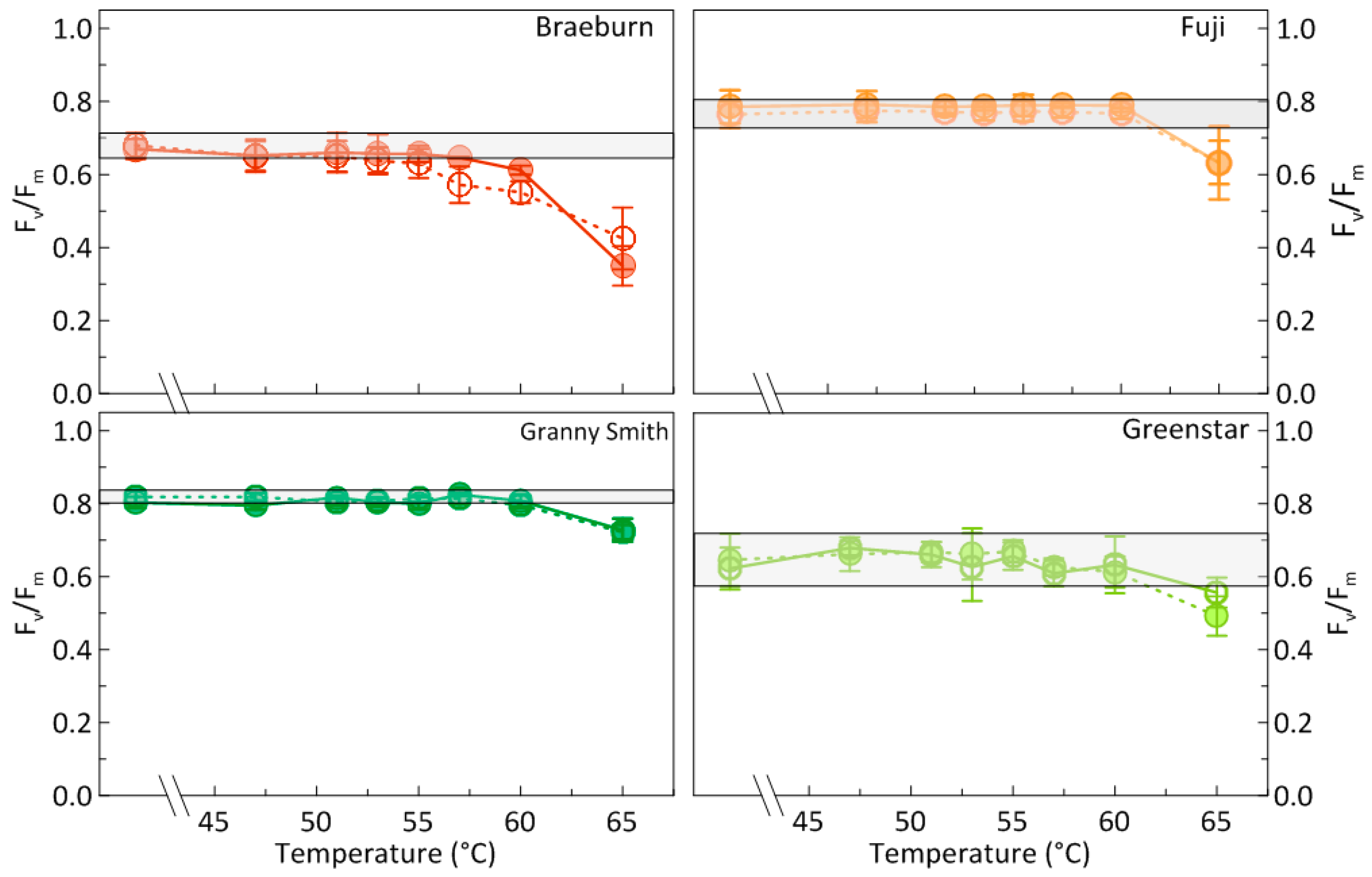
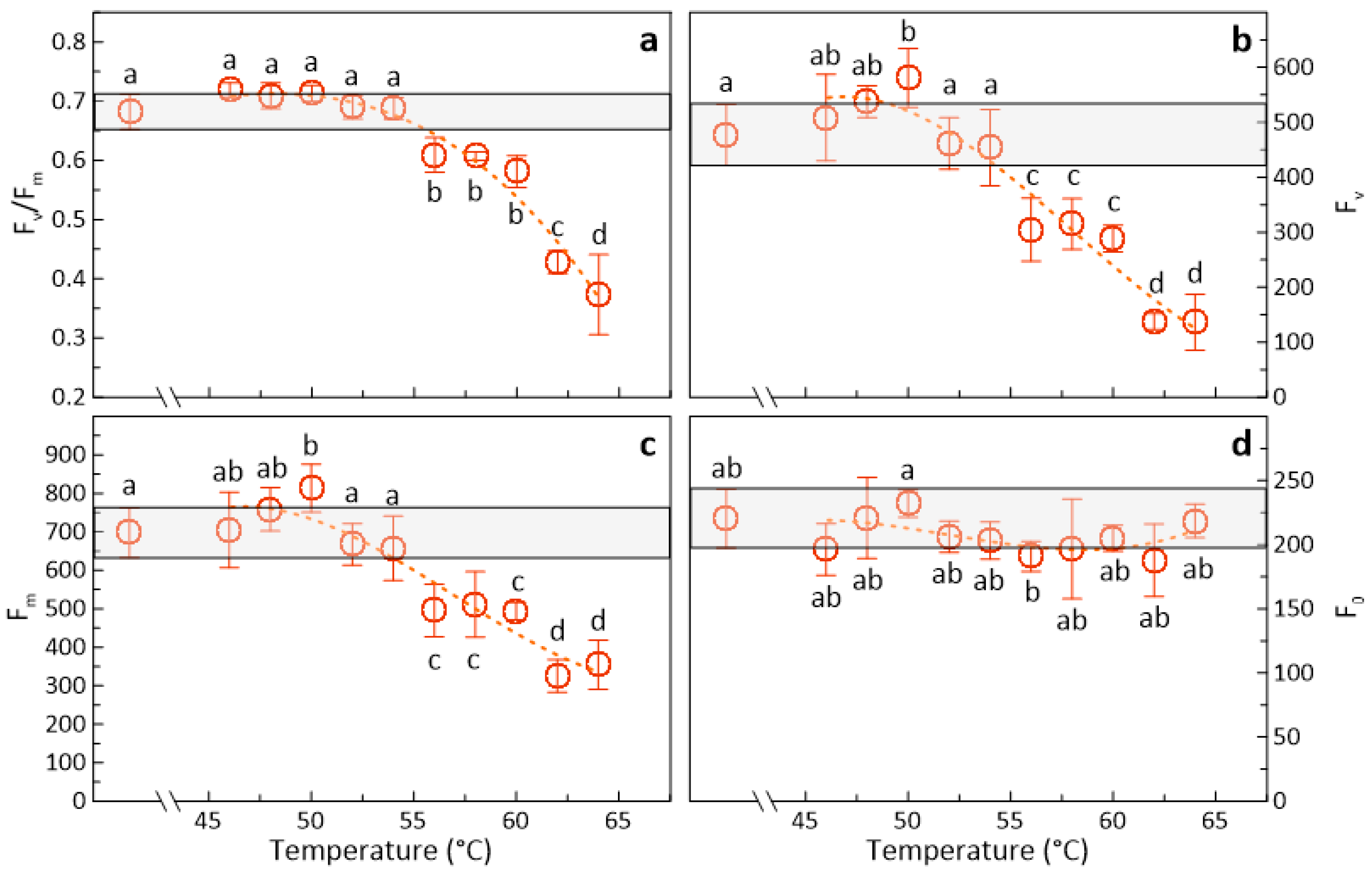
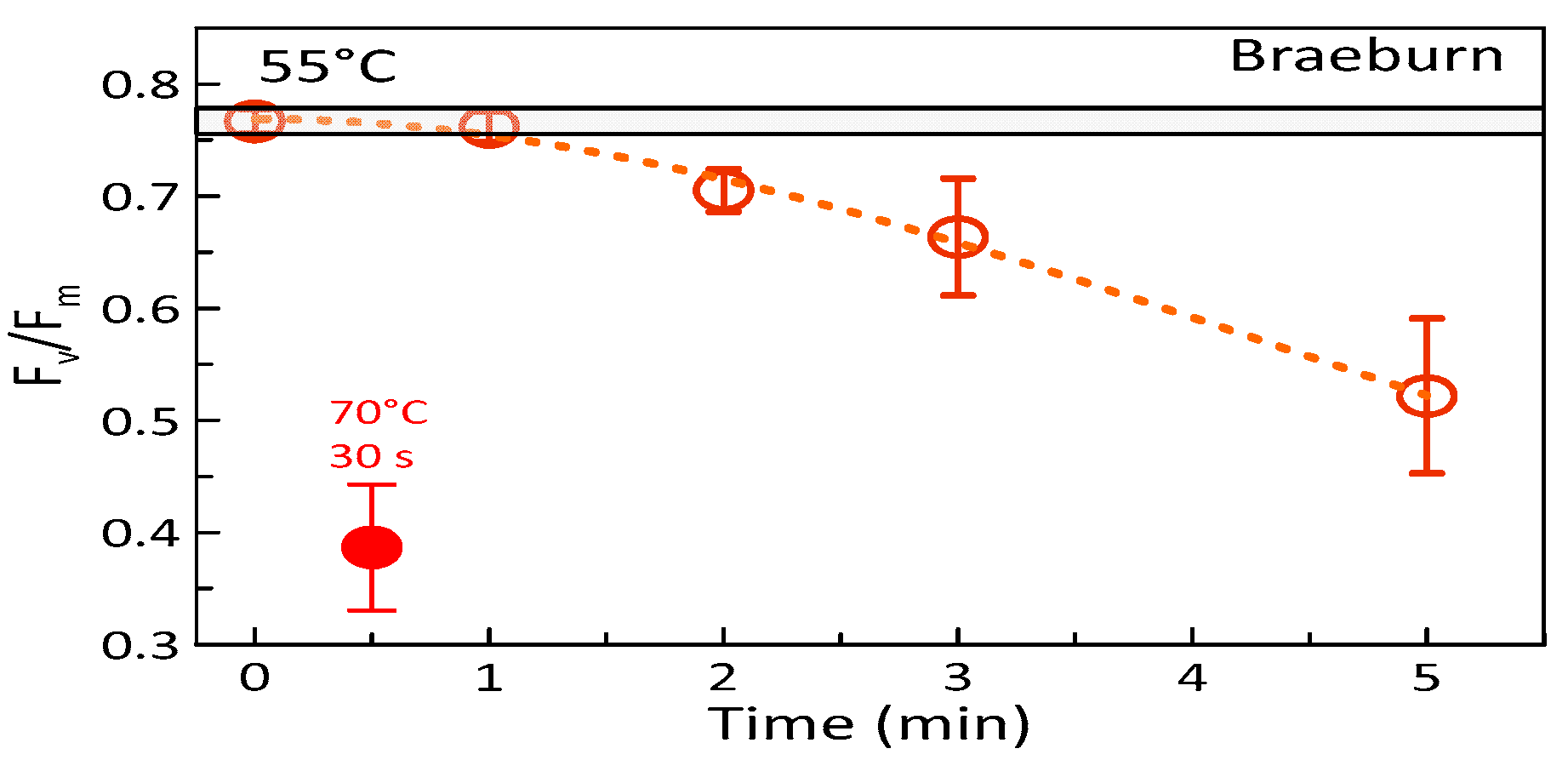
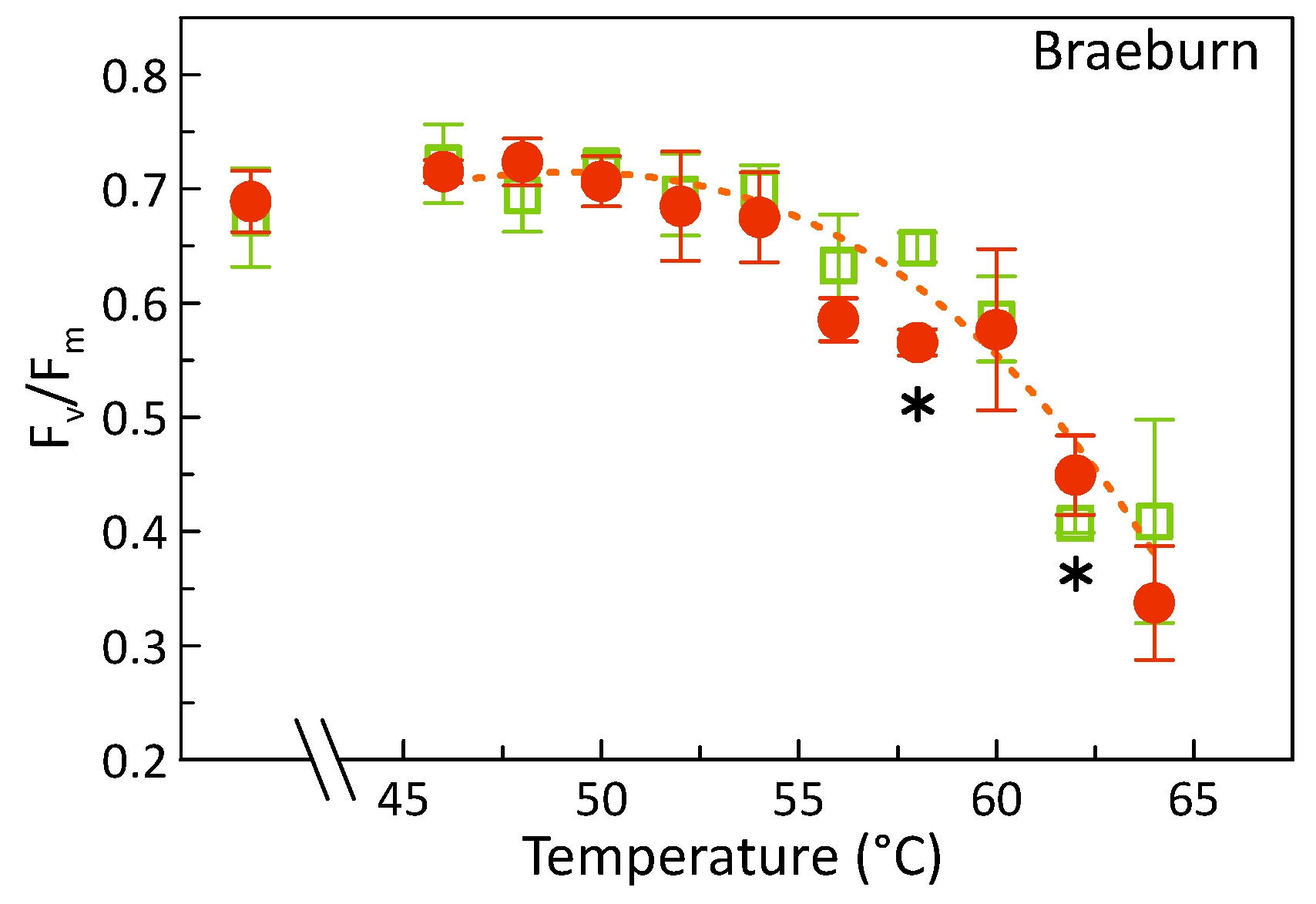
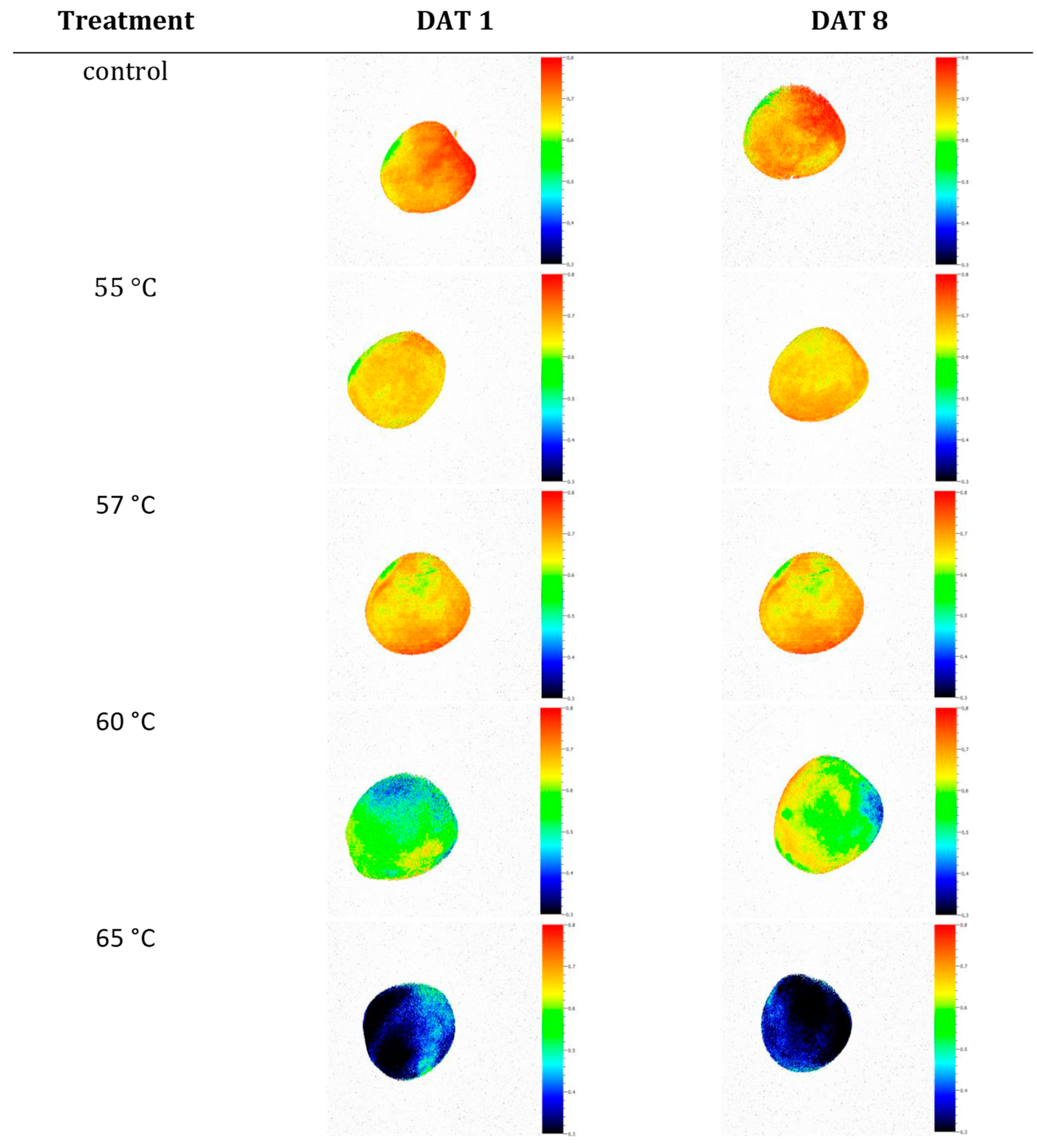
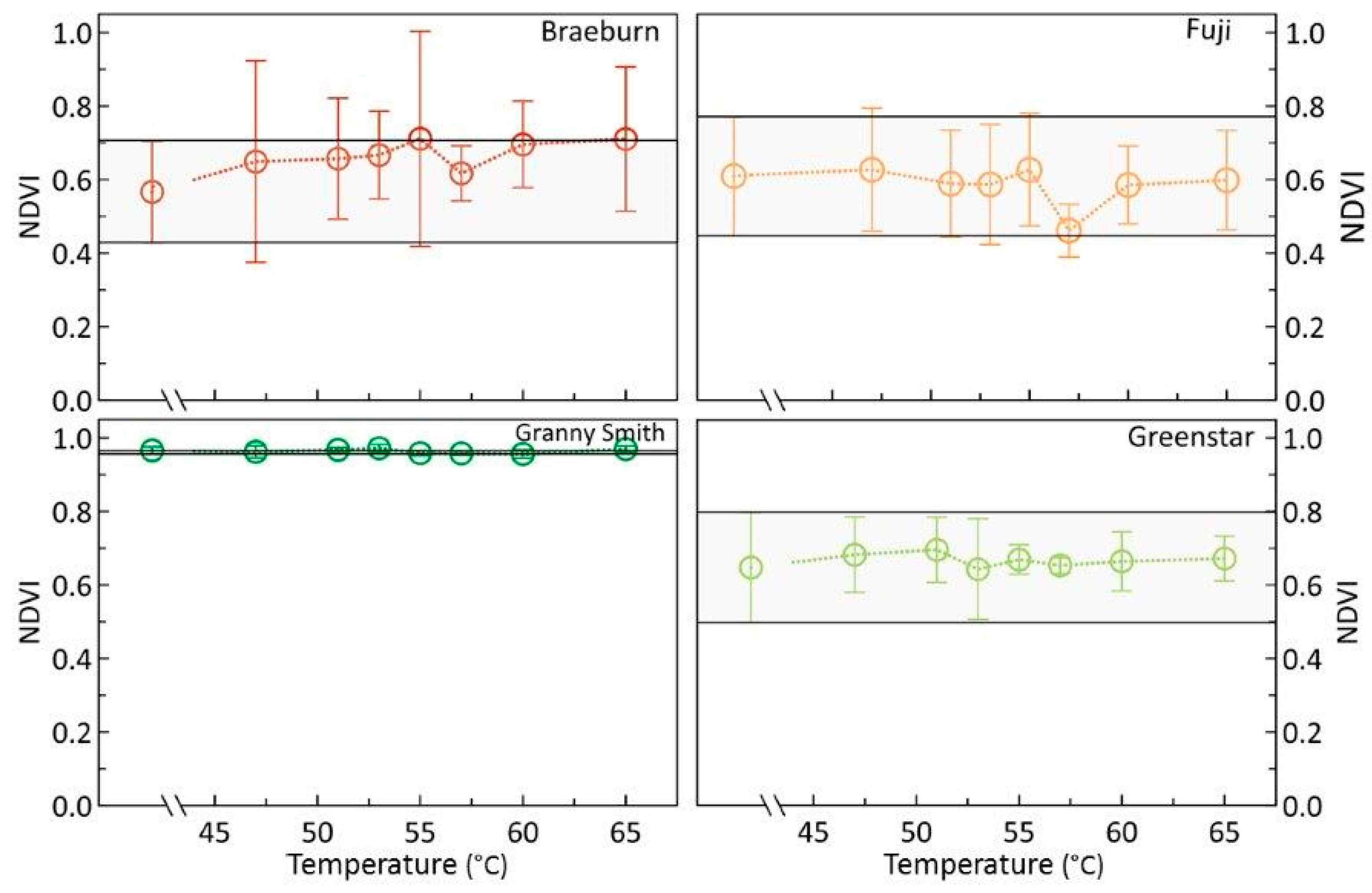
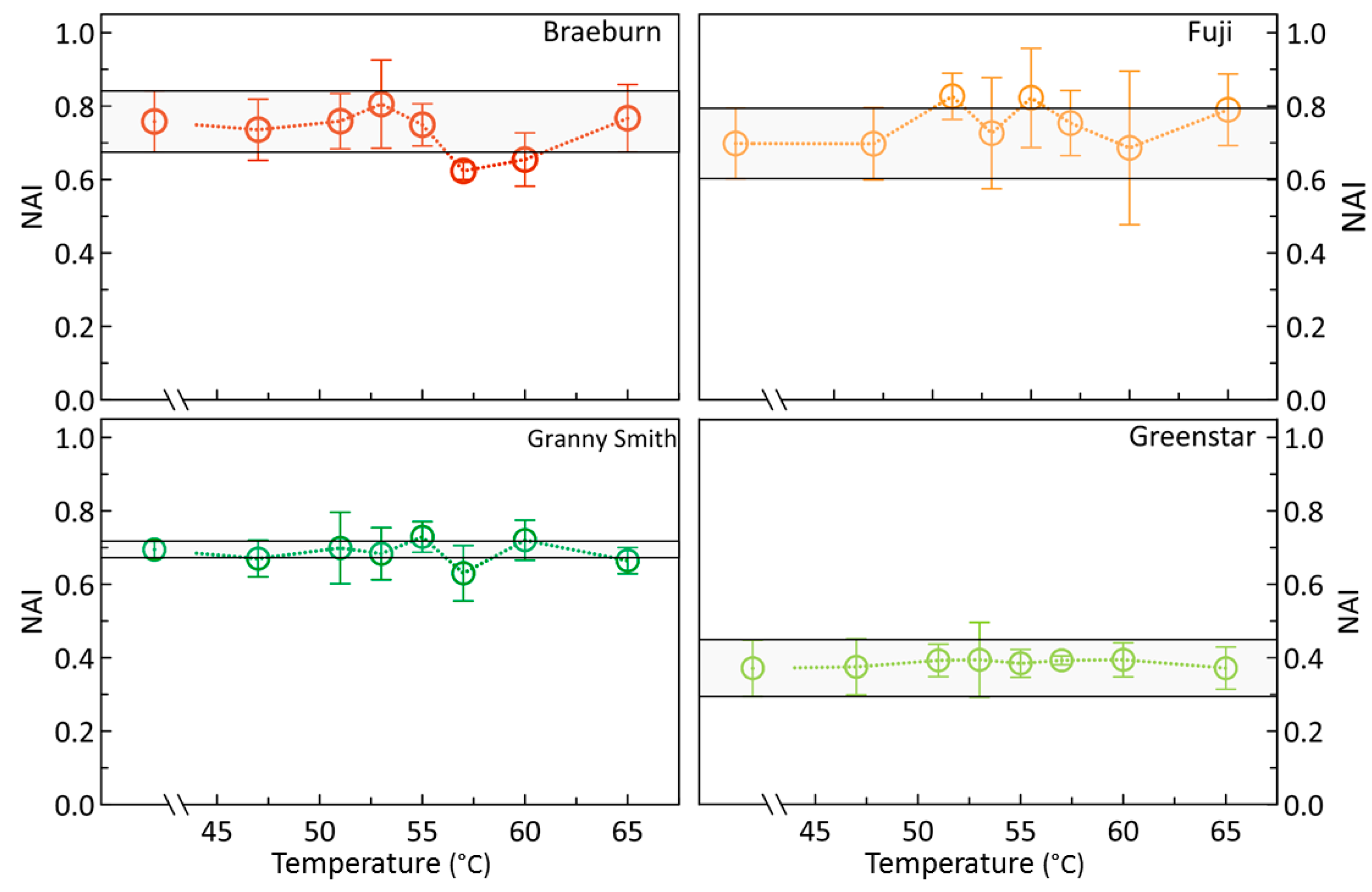
3.1. Effects of Heat Treatments on Photosynthetic Activity of Apples
3.2. Effects of Heat Treatments on Relevant Apple Pigments
3.3. Effects of Heat Treatments on Relevant Quality Parameters of Apples
4. Discussion
4.1. Temperature Effects on Fruit Quality
4.2. Are sHWT Effects Cultivar-Specific?
4.3. Temperature Effects on Fruit Photosynthesis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rux, G.; Caleb, O.; Fröhling, A.; Herppich, W.B.; Mahajan, P. Respiration and storage quality of fresh-cut apple slices immersed in sugar syrup and orange juice. Food Bioprocess Technol. 2017, 10, 2081–2091. [Google Scholar] [CrossRef]
- Trierweiler, B.; Schirmer, H.; Tauscher, B. Hot water treatment to control Gloeosporium disease on apples during long-term storage. J. Appl. Bot. 2003, 77, 156–159. [Google Scholar]
- Kabelitz, T.; Schmidt, B.; Herppich, W.B.; Hassenberg, K. Effects of hot water dipping on apple heat transfer and post-harvest fruit quality. LWT - Food Sci. Technol. 2019, 108, 416–420. [Google Scholar] [CrossRef]
- Spadoni, A.; Guidarelli, M.; Phillips, J.; Mari, M.; Wisniewski, M. Transcriptional profiling of apple fruit in response to heat treatment: involvement of a defense response during Penicillium expansum infection. Postharvest Biol. Technol. 2015, 101, 37–48. [Google Scholar] [CrossRef]
- Schirra, M.; D’hallewin, G.; Ben-Yehoshua, S.; Fallik, E. Host–pathogen interactions modulated by heat treatment. Postharvest Biol. Technol. 2000, 21, 71–85. [Google Scholar] [CrossRef]
- Auinger, A.; Trierweiler, B.; Lücke, F.K.; Tauscher, B. Influence of hot water treatment on different quality parameters of apples during storage. J. Appl. Bot. Food Qual. 2005, 79, 154–156. [Google Scholar]
- Li, L.; Li, X.; Wang, A.; Jiang, Y.; Ban, Z. Effect of heat treatment on physiochemical, colour, antioxidant and microstructural characteristics of apples during storage. Int. J. Food Sci. Technol. 2013, 48, 727–734. [Google Scholar] [CrossRef]
- Fallik, E.; Ilic, Z. Positive and negative effects of heat treatment on the incidence of physiological disorders in fresh produce. In Postharvest Physiological Disorders in Fruits and Vegetables; Tonetto de Freitas, S., Pareek, S., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 111–126. [Google Scholar] [CrossRef]
- Fallik, E. Prestorage hot water treatments (immersion, rinsing and brushing). Postharvest Biol. Technol. 2004, 32, 125–134. [Google Scholar] [CrossRef]
- Maxin, P.; Williams, M.; Weber, R.W. Control of fungal storage rots of apples by hot-water treatments: a Northern European perspective. Erwerbs-Obstbau 2014, 56, 25–34. [Google Scholar] [CrossRef]
- Kabelitz, T.; Hassenberg, K. Control of apple surface microflora for fresh-cut produce by post-harvest hot-water treatment. LWT - Food Sci. Technol. 2018, 98, 492–499. [Google Scholar] [CrossRef]
- Schlüter, O.; Foerster, J.; Geyer, M.; Knorr, D.; Herppich, W.B. Characterization of high hydrostatic pressure effects on fresh produce using chlorophyll fluorescence image analysis. Food Bioprocess Technol. 2009, 2, 291–299. [Google Scholar] [CrossRef]
- Baier, M.; Foerster, J.; Schnabel, U.; Knorr, D.; Herppich, W.B.; Schlüter, O. Direct “non-thermal plasma” treatment for the sanitation of fresh corn salad leaves. Evaluation of physical and physiological effects and antimicrobial efficacy. Postharvest Biol. Technol. 2013, 84, 81–87. [Google Scholar] [CrossRef]
- Herppich, W.B.; Flach, B.M.-T.; von Willert, D.J.; Herppich, M. Field investigations in Welwitschia mirabilis during a severe drought. II. Influence of leaf age, leaf temperature and irradiance on photosynthesis and photoinhibition. Flora 1997, 192, 165–174. [Google Scholar] [CrossRef]
- Matyssek, R.; Herppich, W.B. Experimentelle Pflanzenökologie: Chlorophyllfluoreszenzanalyse; Springer-Verlag GmbH: Berlin, Deutschland, Germany, 2018; pp. 1–55. [Google Scholar]
- Herppich, W.B.; Foerster, J.; Zeymer, J.; Geyer, M.; Schlüter, O. Chlorophyll fluorescence imaging for non-destructively monitoring of changes in fresh and fresh-cut produce. In Environmentally Friendly and Safe Technologies for Quality of Fruits and Vegetables; Nunes, C., Ed.; Universidade do Algave: Faro, Portugal, 2012; pp. 45–51. [Google Scholar]
- Kurenda, A.; Zdunek, A.; Schlüter, O.; Herppich, W.B. VIS/NIR spectroscopy, chlorophyll fluorescence, biospeckle and backscattering to evaluate changes in apples subjected to hydrostatic pressures. Postharvest Biol. Technol. 2014, 96, 88–98. [Google Scholar] [CrossRef]
- Hüve, K.; Bichele, I.; Rasulov, B.; Niinemets, U. When it is too hot for photosynthesis: heat-induced instability of photosynthesis in relation to respiratory burst, cell permeability changes and H2O2 formation. Plant, Cell Environ. 2011, 34, 113–126. [Google Scholar] [CrossRef]
- Kim, D.M.; Smith, N.L.; Lee, C.Y. Apple cultivar variations in response to heat treatment and minimal processing. J. Food Sci. 1993, 58, 1111–1114. [Google Scholar] [CrossRef]
- Shao, X.F.; Tu, K.; Zhao, Y.Z.; Chen, L.; Chen, Y.Y.; Wang, H. Effects of pre-storage heat treatment on fruit ripening and decay development in different apple cultivars. J. Hortic. Sci. Biotechnol. 2007, 82, 297–303. [Google Scholar] [CrossRef]
- Schloffer, K.; Linhard, D. Short time-high temperature hot water shower against Neofabraea rot. In Proceedings of the Ecofruit, 17th International Conference on Organic Fruit-Growing, Hohenheim, Germany, 15–17 February 2016; pp. 176–179. [Google Scholar]
- Hengari, S.; Theron, K.I.; Midgley, S.J.E.; Steyn, W.J. Differential dependence of apple (Malus domestica Borkh.) cultivars on the xanthophyll cycle for photoprotection. South Africa. J. Plant Soil 2016, 33, 69–76. [Google Scholar] [CrossRef]
- Gitelson, A.; Merzlyak, M.N. Quantitative estimation of chlorophyll-a using reflectance spectra: Experiments with autumn chestnut and maple leaves. J. Photochem. Photobiol. B: Biol. 1994, 22, 247–252. [Google Scholar] [CrossRef]
- Zude-Sasse, M.; Truppel, I.; Herold, B. An approach to non-destructive apple fruit chlorophyll determination. Postharvest Biol. Technol. 2002, 25, 123–133. [Google Scholar] [CrossRef]
- Merzlyak, M.N.; Solovchenko, A.E.; Gitelson, A.A. Reflectance spectral features and non-destructive estimation of chlorophyll, carotenoid and anthocyanin content in apple fruit. Postharvest Biol. Technol. 2003, 27, 197–211. [Google Scholar] [CrossRef]
- Kuckenberg, J.; Tartachnyk, I.; Noga, G. Evaluation of fluorescence and remission techniques for monitoring changes in peel chlorophyll and internal fruit characteristics in sunlit and shaded sides of apple fruit during shelf-life. Postharvest Biol. Technol. 2008, 48, 231–241. [Google Scholar] [CrossRef]
- Landahl, S.; Herppich, W.B.; Herold, B.; Geyer, M.; De Baerdemaeker, J. A comprehensive evaluation of the interactions between produce elasticity and water status. Europ. J. Hortic. Sci. 2004, 69, 250–257. [Google Scholar]
- Smith, K.J.; Lay-Yee, M. Response of ‘Royal Gala’ apples to hot water treatment for insect control. Postharvest Biol. Technol. 2000, 19, 111–122. [Google Scholar] [CrossRef]
- Hemmaty, S.; Moallemi, N.; Naseri, L. Effect of UV-C radiation and hot water on the calcium content and postharvest quality of apples. Span. J. Agric. Res. 2007, 5, 559–568. [Google Scholar] [CrossRef]
- Tahir, I.I.; Johansson, E.; Olsson, M.E. Improvement of apple quality and storability by a combination of heat treatment and controlled atmosphere storage. HortScience 2009, 44, 1648–1654. [Google Scholar] [CrossRef] [Green Version]
- Rux, G.; Efe, E.; Ulrichs, C.; Huyskens-Keil, S.; Hassenberg, K.; Herppich, W.B. Effects of pre-processing hot-water treatment on quality and shelf life of fresh cut apple slice. Foods 2019, 8, 653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Birla, S.L.; Tang, J.; Hansen, J.D. Postharvest treatment to control codling moth in fresh apples using water assisted radio frequency heating. Postharvest Biol. Technol. 2006, 40, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.-S.; Li, P.; Cheng, L. Comparison of thermotolerance of sun-exposed peel and shaded peel of ‘Fuji’ apple. Environ. Exp. Bot. 2009, 66, 110–116. [Google Scholar] [CrossRef]
- Awad, M.A.; de Jager, A. Formation of flavonoids, especially anthocyanin and chlorogenic acid in ‘Jonagold’ apple skin: influences of growth regulators and fruit maturity. Sci. Hortic. 2002, 93, 257–266. [Google Scholar] [CrossRef]
- Delgado-Pelayo, R.; Gallardo-Guerrero, L.; Hornero-Méndez, D. Chlorophyll and carotenoid pigments in the peel and flesh of commercial apple fruit varieties. Food Res. Inter. 2014, 65, 272–281. [Google Scholar] [CrossRef] [Green Version]
- Fallik, E.; Tuvia-Alkalai, S.; Feng, X.; Lurie, S. Ripening characterisation and decay development of stored apples after a short pre-storage hot water rinsing and brushing. Innov. Food Sci. Emerg. Technol. 2001, 2, 127–132. [Google Scholar] [CrossRef]
- Yüzbaşıoğlu, E.; Dalyan, E.; Akpinar, I. Changes in photosynthetic pigments, anthocyanin content and antioxidant enzyme activities of maize (Zea mays L.) seedlings under high temperature stress conditions. Trakya Univ. J. Nat. Sci. 2017, 18, 97–104. [Google Scholar] [CrossRef]
- Fan, L.; Song, J.; Forney, C.F.; Jordan, M.A. Fruit maturity affects the response of apples to heat stress. Postharvest Biol. Technol. 2011, 62, 35–42. [Google Scholar] [CrossRef]
- Hengari, S.; Theron, K.I.; Midgley, S.J.E.; Steyn, W.J. Response of apple (Malus domestica Borkh.) fruit peel photosystems to heat stress coupled with moderate photosynthetic active radiation at different fruit developmental stages. Sci. Hortic. 2014, 178, 154–162. [Google Scholar] [CrossRef]
- Mpelasoka, B.S.; Behoudian, M.H.; Mills, T.M. Effects of deficit irrigation on fruit maturity and quality of ‘Braeburn’ apple. Sci. Hortic. 2001, 90, 279–290. [Google Scholar] [CrossRef]
- Garriz, P.I.; Colavita, G.M.; Alvarez, H.L.; Blackhall, V. Maturity of ‘Braeburn’ apple fruit as affected by harvest timing and season. Acta Hortic. 2012, 932, 245–250. [Google Scholar] [CrossRef]
- Hägele, F.; Baur, S.; Menegat, A.; Gerhards, R.; Carle, R.; Schweiggert, R.M. Chlorophyll fluorescence imaging for monitoring the effects of minimal processing and warm water treatments on physiological properties and quality attributes of fresh-cut salads. Food Bioprocess Technol. 2016, 9, 650–663. [Google Scholar] [CrossRef]
- Havaux, M. Stress tolerance of photosystem-II in vivo–antagonistic effects of water, heat, and photoinhibition stresses. Plant Physiol. 1992, 100, 424–432. [Google Scholar] [CrossRef] [Green Version]
- Pastenes, C.; Horton, P. Resistance of photosynthesis to high temperature in two bean varieties (Phaseolus vulgaris L.). Photosynth. Res. 1999, 62, 197–203. [Google Scholar] [CrossRef]
- Zhang, R.; Cruz, J.A.; Kramer, D.M.; Magallanes-Lundback, M.E.; Dellapenna, D.; Sharkey, T.D. Moderate heat stress reduces the pH component of the transthylakoid proton motive force in light-adapted, intact tobacco leaves. Plant Cell Environ. 2009, 32, 1538–1547. [Google Scholar] [CrossRef]
- Kouřil, R.; Laźar, D.; Ilík, P.; Skotnica, J.; Krchňák, P.; Nauš, J. High-temperature induced chlorophyll fluorescence rise in plants at 40–50 °C: experimental and theoretical approach. Photosynth. Res. 2004, 81, 49–66. [Google Scholar] [CrossRef]
- Ivanov, A.G.; Velitchkova, M.Y.; Allakhverdiev, S.I.; Huner, N.P.A. Heat stress-induced effects of photosystem I: an overview of structural and functional responses. Photosynth. Res. 2017, 133, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Krause, H.G.; Weiss, E. Chlorophyll fluorescence and photosynthesis: The basics. Annu. Rev. Plant Physiol. Plant Molec. Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Haltia, T.; Freire, E. Forces and factors that contribute to the structural stability of membrane proteins. Biochim. Biophys. Acta 1994, 1241, 295–322. [Google Scholar] [CrossRef]
- Maheswari, M.; Joshi, D.K.; Saha, R.; Nagarajan, S.; Gambhir, P.N. Transverse Relaxation Time of Leaf Water Protons and Membrane Injury in Wheat (Triticum aestivum L.) in Response to High Temperature. Ann. Bot. 1999, 84, 741–745. [Google Scholar] [CrossRef] [Green Version]
- Nauš, J.; Kuropatwa, R.; Klinkovskÿ, T.; Ilík, P.; Lattová, J.; Pavlová, Z. Heat injury of barley leaves detected by the chlorophyll fluorescence temperature curve. Biochim Biophys. Acta 1992, 1101, 359–362. [Google Scholar] [CrossRef]
- Bilger, H.-W.; Schreiber, U.; Lange, O.L. Determination of leaf heat resistance: comparative investigation of chlorophyll fluorescence changes and tissue necrosis methods. Oecologia 1984, 63, 256–262. [Google Scholar] [CrossRef]
- Wand, S.J.E.; van den Dool, K.; Smit, A.; Steyn, W.J. Heat injury thresholds in apples measured using chlorophyll fluorescence are influenced by orchard heat reduction technologies. Acta Hortic. 2008, 772, 273–278. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vitamin C (g m−3) | TSS (%) | Malic acid (kg m−3) | Strength (N) | Stiffness (kN m−1) | |
|---|---|---|---|---|---|
| ‘Braeburn’ control | 106.0 ± 21.6a | 11.70 ± 1.10a | 3.53 ± 0.80a | 67.9 ± 2.9a | 34.0 ± 3.6a |
| ‘Braeburn’ DAT1 | 91.9 ± 18.2ab | 11.70 ± 0.68a | 3.80 ± 0.91a | 55.5 ± 7.3b | 28.3 ± 3.0b |
| ‘Braeburn’ DAT8 | 83.6 ± 18.4b | 11.70 ± 0.67a | 3.78 ± 0.85a | 48.5 ± 6.5c | 25.4 ± 3.3c |
| ‘Fuji’ control | 76.2 ± 14.79a | 15.38 ± 1.27ab | 2.90 ± 0.28a | 80.6 ± 7.5a | 35.7 ± 0.3a |
| ‘Fuji’ DAT1 | 65.7 ± 11.6a | 14.27 ± 2.58a | 3.06 ± 0.48a | 79.2 ± 9.0a | 33.3 ± 2.4a |
| ‘Fuji’ DAT8 | 70.2 ± 11.9a | 15.72 ± 2.51b | 3.11 ± 0.56a | 80.9 ± 6.2a | 31.3 ± 4.0b |
| ‘Granny Smith’ control | 93.1 ± 5.4a | 11.22 ± 0.24ab | 9.70 ± 0.89a | 98.5 ± 6.6a | 39.9 ± 5.5a |
| ‘Granny Smith’ DAT1 | 90.9 ± 14.7a | 11.13 ±0.46a | 10.55 ± 1.52a | 96.6 ± 5.3a | 39.9 ± 3.9a |
| ‘Granny Smith’ DAT8 | 90.6 ± 14.7a | 11.62 ±0.47b | 10.08 ± 1.32a | 92.7 ± 5.3b | 38.8 ± 3.9a |
| ‘Greenstar’ control | 137.3 ± 10.8a | 11.27 ±0.92ab | 4.47 ± 0.99a | 92.0 ±.9.6a | 34.0 ± 5.7a |
| ‘Greenstar’ DAT1 | 144.3 ± 14.2a | 11.58 ±0.45b | 4.51 ± 0.93a | 94.0 ± 5.0a | 34.2 ± 3.1a |
| ‘Greenstar’ DAT8 | 135.1 ± 13.6a | 11.12 ±0.48a | 4.48 ± 0.83a | 90.9 ± 5.8a | 32.5 ± 2.9a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herppich, W.B.; Maggioni, M.; Huyskens-Keil, S.; Kabelitz, T.; Hassenberg, K. Optimization of Short-Term Hot-Water Treatment of Apples for Fruit Salad Production by Non-Invasive Chlorophyll-Fluorescence Imaging. Foods 2020, 9, 820. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9060820
Herppich WB, Maggioni M, Huyskens-Keil S, Kabelitz T, Hassenberg K. Optimization of Short-Term Hot-Water Treatment of Apples for Fruit Salad Production by Non-Invasive Chlorophyll-Fluorescence Imaging. Foods. 2020; 9(6):820. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9060820
Chicago/Turabian StyleHerppich, Werner B., Marco Maggioni, Susanne Huyskens-Keil, Tina Kabelitz, and Karin Hassenberg. 2020. "Optimization of Short-Term Hot-Water Treatment of Apples for Fruit Salad Production by Non-Invasive Chlorophyll-Fluorescence Imaging" Foods 9, no. 6: 820. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9060820